單親滅活德氏乳桿菌和乳酸乳球菌原生質體融合條件優化
2012-10-28 07:06:02黃潔潔付瑞燕
食品科學 2012年5期
關鍵詞:融合
李 麗,房 杰,黃潔潔,付瑞燕*
(安徽農業大學茶與食品科技學院,安徽 合肥 230036)
單親滅活德氏乳桿菌和乳酸乳球菌原生質體融合條件優化
李 麗,房 杰,黃潔潔,付瑞燕*
(安徽農業大學茶與食品科技學院,安徽 合肥 230036)
利用單親滅活原生質體技術對德氏乳桿菌FQ菌株和乳酸乳球菌FL菌株的原生質體進行融合,考察原生質體制備、再生和融合條件的影響因素。結果表明:制備德氏乳桿菌FQ菌株原生質體最適條件為溫度37℃,在含有10μg/mL變溶菌素和1mg/mL溶菌酶溶液中超聲處理90min。在此條件下,原生質體再生率可達6.36%。乳酸乳球菌FL菌株添加1mg/mL甘氨酸處理后,用10mg/mL溶菌酶37℃恒溫酶解90min,原生質體形成率可達99.97%。65℃處理乳酸乳球菌FL菌株原生質體120min,原生質體滅活率可達96.89%。融合實驗結果表明,在PEG6000 400g/L(含0.02mol/L MgCl2和0.01mol/L CaCl2)、融合時間5min、融合溫度20℃、pH6.5的條件下促融,德氏乳桿菌FQ菌株和乳酸乳球菌FL菌株原生質體的融合率可達2.72×10-6。
單親滅活;德氏乳桿菌;乳酸乳球菌;原生質體融合
乳酸是世界上公認的三大有機酸之一,在食品、醫藥和生物材料等諸多領域具有良好的應用前景[1-2]。另外,乳酸經過聚合生成的直鏈或環狀聚乳酸,是一種無毒、無刺激性、強度高、可塑性好、生物相容性優良、具有良好生物可降解性的新型生物,已引起世界各國濃厚的興趣。因此,國內外對乳酸的生產研究非常重視[3]。
乳酸發酵的生產菌種主要為乳酸菌和根霉菌。與根霉菌發酵相比,乳酸菌發酵的產酸量高,并且對糖的利用率高,發酵不需要通氧氣,動力消耗小。但是,這種厭氧或微耗氧發酵的周期較長,造成生產效率偏低。究其原因,是因乳酸菌對氧的耐受能力不足,在接觸氧的過程中所產生的氧自由基對細胞活性造成損傷,從而降低菌株的最大生長速率和碳源利用程度。德氏乳桿菌是工業生產乳酸最常用的菌種[4-6],因此,選育耐氧型德氏乳桿菌具有重要的現實意義。與傳統誘變相比,原生質體融合具有集中雙親本優良性狀等優勢[7]。通過原生質體融合方法選育具有優良性狀融合菌株的方法已經成功應用于提高嗜酸乳桿菌耐酸及耐膽鹽能力[8]等諸多方面的研究。本實驗對德氏乳桿菌FQ菌株原生質體再生條件、乳酸乳球菌FL菌株原生質體溫度滅活條件及原生質體融合條件進行優化研究,對單親滅活原生質體技術融合選育耐氧乳酸菌進行探索,為耐氧型乳酸菌基因工程菌的構建提供基礎。
1 材料與方法
1.1 菌株與試劑
德氏乳桿菌FQ菌株和乳酸乳球菌FL菌株由安徽農業大學茶與食品科技學院微生物學實驗室保藏。
MRS、M17培養基和變溶菌素 美國Sigma公司;溶菌酶、聚乙二醇(PEG6000) 合肥博美生物科技有限責任公司;其他試劑均為國產分析純。
1.2 方法
1.2.1 生長培養基配方
德氏乳桿菌FQ菌株:MRS培養基,補充0.5mol/L蔗糖,簡稱SMRS;乳酸乳球菌FL菌株:M17培養基,補充10g/L葡萄糖,簡稱GM17。
1.2.2 再生培養基(RM)配方
再生固體培養基Ⅰ(RMⅠ):MRS培養基(不含Tween-80)組成成分,另外再補充0.5mol/L蔗糖、20mmol/L MgCl2、10mmol/L CaCl2、25g/L明膠、5g/L牛血清白蛋白(BSA)、15g/L瓊脂。再生半固體培養基瓊脂成分為6g/L。
再生固體培養基Ⅱ(RMⅡ):MRS培養基(含Tween-80)組成成分,另外再補充0.5mol/L蔗糖、20mmol/L MgCl2、10mmol/L CaCl2、25g/L明膠、15g/L瓊脂。
再生固體培養基Ⅲ(RMⅢ):MRS培養基(含Tween-80)組成成分,另外再補充0.5mol/L蔗糖、20mmol/L MgCl2、10mmol/L CaCl2、15g/L 瓊脂。
再生固體培養基Ⅳ(RMⅣ)[9]:MR S培養基(含Tween-80)組成成分,另外再補充0.5mol/L蔗糖、15g/L瓊脂。
再生固體培養基Ⅴ(RMⅤ)[10]:25mmol/L葡萄糖、5g/L胰蛋白胨、5g/L酵母提取物、2mmol/L L-半胱氨酸鹽酸鹽、18mmol/L磷酸二氫鉀、122mmol/L醋酸鈉、25mmol/L MgCl2、45mmol/L CaCl2、0.25mol/L 棉子糖、15g/L瓊脂。
再生固體培養基Ⅵ(RMⅥ)[11]:改良Chamers培養基組成成分,另外再補充0.7mol/L KCl、0.5mol/L蔗糖、0.75mol/L甘露醇、15g/L瓊脂。
1.2.3 原生質體穩定液(PB)的配制
Tris-HCl緩沖液[9,12]:10mmol/L Tris-HCl、0.5mol/L蔗糖和20mmol/L MgCl2,用超純水配制,用4mol/L HCl溶液調節pH值至6.5,115℃高壓滅菌20min,冷卻至室溫后4℃保存備用。
1.3 德氏乳桿菌FQ菌株原生質體的制備及再生
將變溶菌素和溶菌酶用滅菌原生質穩定液配制成100μg/mL和100mg/mL貯備液,濾膜過濾除菌,備用。取SMRS液體培養基培養至對數末期的菌液15mL,4℃、4000r/min離心5min,去上清液。用5mL PB反復洗滌兩次,去上清液。最后用1mL PB懸浮即得菌體懸浮液,最終濃度為1×109~4×109CFU/mL。懸浮液加入終質量濃度分別為1μg/mL變溶菌素、5μg/mL變溶菌素、10μg/mL變溶菌素、10μg/mL變溶菌素+1mg/mL溶菌酶、3mg/mL溶菌酶,37℃恒溫酶解,同時進行超聲處理120min。再利用最適酶質量濃度酶解不同時間(30、60、90、120mim),得到原生質體最大再生率。酶解后的菌液用PB洗滌兩次,用1mL PB懸浮得到原生質體液。將菌體懸浮液和原生質體液分別用0.85g/100mL生理鹽水、無菌水和PB梯度稀釋后涂布于MRS雙層培養基和6種雙層再生培養基,37℃恒溫培養5~7d,測定德氏乳桿菌FQ菌株原生質體的形成率和再生率。
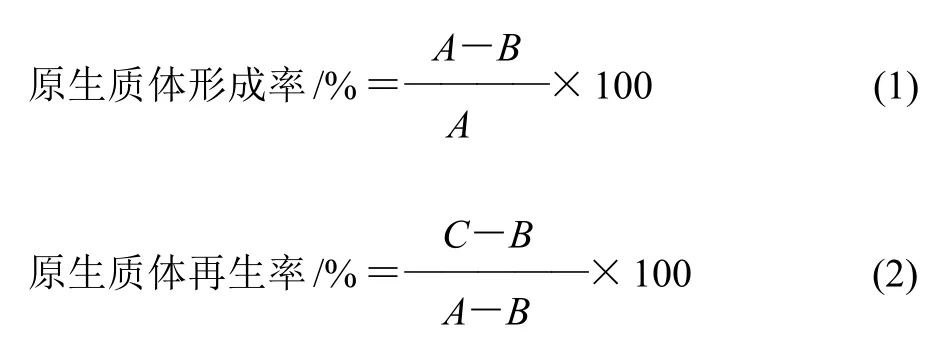
式中:A為酶解前的總菌落數(MRS培養基)/(CFU/mL);B為未形成原生質體菌落數(即未被酶裂解的剩余細胞數,MRS培養基)/(CFU/mL);C為酶解后的再生菌落數(RM培養基)/(CFU/mL)。
1.4 乳酸乳球菌FL菌株原生質體制備及形成
取GM17液體培養基培養至對數末期的菌液10mL,4℃、4000r/min離心5min,用PB反復洗滌兩次,用1mL PB懸浮得到菌體懸浮液,最終濃度為6×109~9×109CFU/mL。取1mL懸浮液加入終質量濃度分別為1、5、10mg/mL溶菌酶,37℃恒溫酶解30、60、90min。酶解后的菌液用PB洗滌兩次,用1mL PB懸浮得到原生質體液。取部分懸浮液和原生質體液,用0.85g/100mL生理鹽水和無菌水梯度稀釋后涂布在GM17單層培養基,30℃培養5d,測定乳酸乳球菌FL菌株原生質體形成率。
1.5 甘氨酸對乳酸乳球菌FL菌株原生質體形成率的影響
在GM17培養基中添加1、10、20、30mg/mL甘氨酸培養至對數期,以未添加甘氨酸為對照組。菌體懸浮液中加入終質量濃度為10mg/mL溶菌酶,37℃恒溫酶解90min,考察甘氨酸處理對乳酸乳球菌FL菌株細胞光密度(OD600nm)值、菌落數及原生質體形成率的影響。
1.6 乳酸乳球菌FL菌株原生質體溫度滅活條件的確定
將制備好的乳酸乳球菌FL菌株原生質體液等量分裝于編號分別為0、1、2、3、4、5的無菌離心管內,每管均設3個平行樣,分別在60、65、70℃的恒溫水浴鍋中熱滅活。同一溫度下的滅活時間間隔為30min。0編號管為對照管(未滅活管)。最后將各管原生質體液適當稀釋后涂布于單層再生培養基中,30℃培養5~7d后,計算原生質體滅活率。同時,用血球計數板觀察細胞是否破裂。
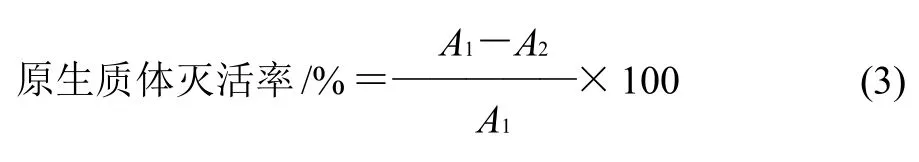
式中:A1為未被處理菌體長出的菌落數/(CFU/mL);A2為被處理菌體長出的菌落數/(CFU/mL)。
1.7 原生質體融合
親本菌株德氏乳桿菌FQ菌株在含有氯霉素再生培養基上不生長,另一親本乳酸乳球菌FL菌株在含有氯霉素再生培養基上可以生長,融合子如果能夠結合兩親本的特性,應該可以抗氯霉素,因此,在含有氯霉素的再生培養基上長出的菌落初步認為其是融合子。
分別取等量德氏乳桿菌FQ菌株原生質體液和滅活乳酸乳球菌FL菌株原生質體液混和(取樣稀釋涂布再生平板和MRS平板),離心,重懸于0.5mL PB中,加4.5mL 400g/L PEG6000(含 0.01mol/L CaCl2、0.02 mol/L MgCl2),混勻,于4、20、37℃條件下分別放置5、10、30min。最后用5mL PB洗滌兩次,適當稀釋后涂布于含氯霉素的最適雙層再生培養基,37℃培養7d后計數。以未經PEG6000處理的兩種原生質體再生菌株作為對照組。
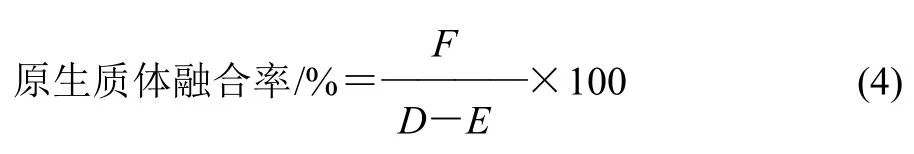
式中:F為融合子數/(CFU/mL);D為兩親本菌株在再生平板菌落數/(CFU/mL);E為兩親本菌株在MRS平板菌落數/(CFU/mL)。
1.8 融合子和兩親本菌株對氧耐受性的比較
將融合子、親本德氏乳桿菌FQ菌株和乳酸乳球菌FL菌株按體積分數5%接種量,融合子和親本乳酸乳球菌FL菌株接入含終質量濃度為10μg/mL氯霉素的MRS培養基,親本德氏乳桿菌FQ菌株接入不含氯霉素的MRS培養基,37℃條件下150r/min振蕩和靜置培養。在不含氯霉素的培養基中振蕩培養作為對照,定時測量其OD600nm值,比較它們對氧的耐受性。
2 結果與分析
2.1 德氏乳桿菌FQ菌株原生質體形成率和再生率
2.1.1 酶種類及其質量濃度對德氏乳桿菌FQ菌株原生質體形成率和再生率的影響
制備細菌原生質體主要采用溶菌酶,但乳桿菌細胞壁致密,對溶菌酶具有很強的抗性,所以在乳桿菌原生質體的制備過程中,采用質量濃度適合的溶菌酶和變溶菌素共同作用,以有效提高原生質體的形成率和再生率[13]。
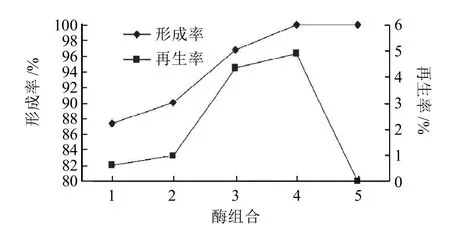
圖1 不同酶種類及其質量濃度對德氏乳桿菌FQ菌株原生質體形成率及再生率的影響Fig.1 Effects of types and concentration of enzymes on formation and regeneration rate of protoplast of Lactobacillus delbrueckii FQ
由圖1可知,采用不同質量濃度的溶菌酶和變溶菌素于37℃恒溫酶解及超聲處理菌液,其原生質體的形成率和再生率隨著變溶菌素質量濃度的增加,逐漸增大,變溶菌素和溶菌酶終質量濃度分別為10μg/mL和1mg/mL時,得到最大的形成率和再生率,分別為99.99%和4.93%。其原因可能是酶解過程中,酶質量濃度升高,增加了酶分子與細胞壁接觸的機會,使得細胞壁被水解的幾率增大,且原生質體具有較高活性。當溶菌酶終質量濃度為3mg/mL時,原生質體不能再生。可能是溶菌酶質量濃度過高,水解完細胞壁后,會再水解細胞膜上的部分蛋白成分從而導致細胞的死去,甚至破裂,致使原生質體不能再生。
2.1.2 酶解時間對德氏乳桿菌FQ菌株原生質體再生率的影響
酶解時間過短會導致細胞壁水解不充分,原生質體不能充分釋放,形成率較低;酶解時間過長又將導致原生質體皺縮,而且會損害細胞質膜導致再生率急劇下降[14],不利于原生質體的再生。本實驗用最適酶質量濃度為10μg/mL變溶菌素和1mg/mL溶菌酶,37℃恒溫水浴超聲處理3 0、6 0、9 0、1 2 0 m in,研究不同酶解時間對德氏乳桿菌FQ菌株原生質體再生率的影響。
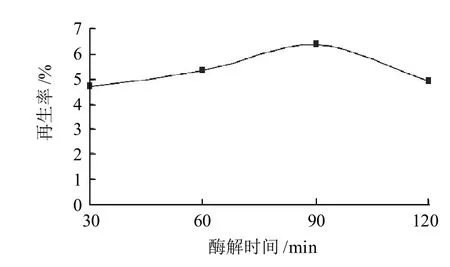
圖2 酶解時間對德氏乳桿菌FQ菌株原生質體再生率的影響Fig.2 Effect of enzymolysis time on protoplast regeneration rate of Lactobacillus delbrueckii FQ
如圖2所示,德氏乳桿菌FQ菌株用最適酶質量濃度于37℃恒溫水浴超聲處理,原生質體形成率為99.99%,沒有隨酶解時間的增加而增大。處理90min,達到最大再生率6.36%。究其原因,可能是適當酶質量濃度和酶解時間使酶分子與細胞壁充分接觸,細胞壁得到水解,原生質體能保持較高的活性,所以原生質體再生率較高。當酶解時間為120min時,可能是酶解時間太長導致部分原生質體皺縮,再生率降低。
2.2 再生培養基種類對德氏乳桿菌FQ菌株再生率的影響
融合子細胞壁合成、重建和融合子的再生,具體過程和一般原生質體再生一樣,酶解脫壁后的原生質體應該具有再生能力,這是原生質體融合育種的必要條件[7]。所以,德氏乳桿菌FQ菌株最適原生質體再生培養基,同樣適合于融合子的再生。本實驗在最適酶解條件下,研究6種再生培養基對德氏乳桿菌FQ菌株原生質體再生率的影響。
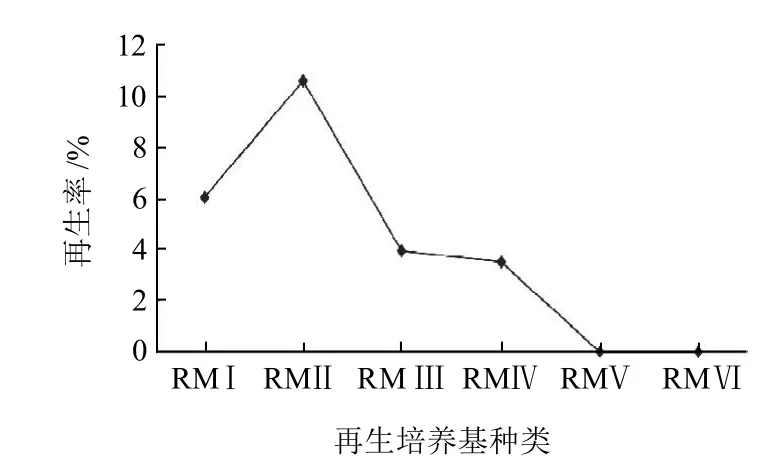
圖3 再生培養基對德氏乳桿菌FQ菌株原生質體再生率的影響Fig.3 Effect of regeneration medium on protoplast regeneration rate of Lactobacillus delbrueckii FQ
由圖3可知,再生培養基Ⅱ(RMⅡ)是德氏乳桿菌FQ菌株原生質體的最適再生培養基,再生率可達到10.59%,再生率大小為:RMⅡ>RMⅠ>RMⅢ>RMⅣ,而RMⅤ和RMⅥ上原生質體不能再生,其原因可能是其滲透壓達不到德氏乳桿菌FQ菌株原生質體再生的水平,導致原生質體無法再生。再生培養基中含有明膠、Ca2+和Mg2+有利于德氏乳桿菌FQ菌株原生質體再生。因此,選擇R MⅡ作為融合子的再生培養基。
2.3 酶質量濃度及酶解時間對乳酸乳球菌FL菌株原生質體形成率的影響
乳酸乳球菌FL菌株細胞壁因含大量肽聚糖而比較致密,而且可以在再生培養基上生長,為了得到純的融合子,要盡可能的使其酶解。
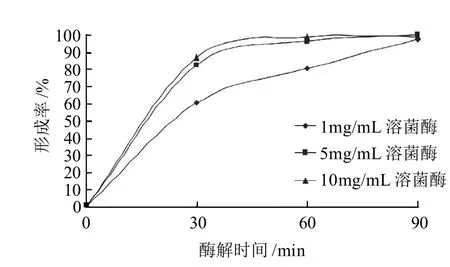
圖4 酶質量濃度及酶解時間對乳酸乳球菌FL菌株原生質體形成率的影響Fig.4 Effect of lysozyme concentration and enzymolysis time on protoplast formation rate of Lactococcus lactis FL
由圖4可知,乳酸乳球菌FL菌株原生質體的形成率隨著溶菌酶質量濃度的升高和酶解時間的增加逐漸升高。酶質量濃度升高,酶分子與細胞壁充分接觸,增大了細胞壁被水解的幾率;適當的酶解時間使細胞壁充分水解,從而得到較高的原生質體形成率。結果表明,用10mg/mL溶菌酶37℃恒溫處理90min,得到最大的原生質體形成率99.95%。
2.4 甘氨酸對乳酸乳球菌FL菌株原生質體形成率的影響
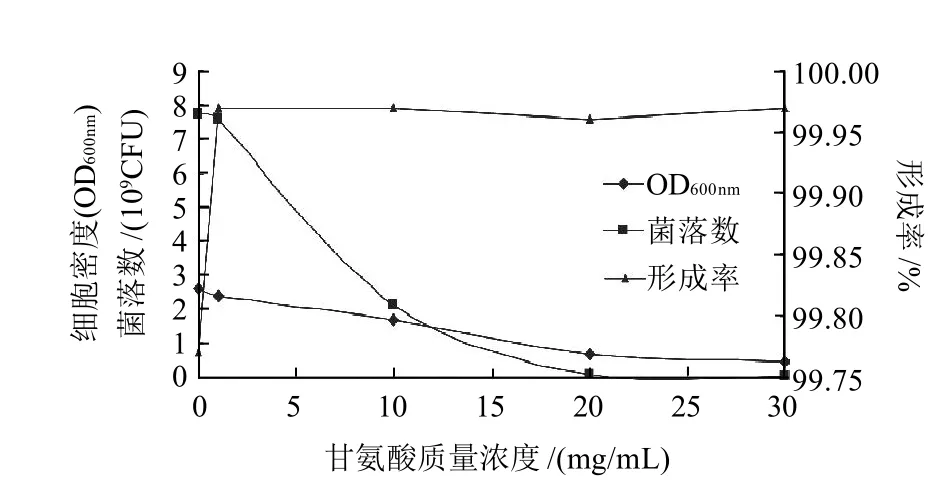
圖5 甘氨酸質量濃度對乳酸乳球菌FL菌株原生質體形成率的影響Fig.5 Effect of glycine concentration on protoplast formation rate of Lactococcus lactis FL
乳酸乳球菌FL菌株細胞壁相對于德氏乳桿菌FQ菌株比較致密,在其生長培養基中添加適當的化學物質可以影響細胞壁的生長,進而影響細胞壁的密度及厚度[15]。常用的細胞壁處理劑如甘氨酸,可以增加菌體對溶菌酶的敏感程度,提高原生質體的制備率。由圖5可知,與對照組相比,實驗組OD600nm及菌落數隨著甘氨酸質量濃度的增加逐漸減小,但原生質體的形成率都略有增加,加入過高質量濃度甘氨酸后,可能是滲透壓過高對細胞壁造成了過度破壞,致使細菌生長非常緩慢。綜合考慮三者,選擇在生長培養基中添加1mg/mL甘氨酸制備原生質體,原生質體的形成率可達到99.97%。
2.5 乳酸乳球菌FL菌株原生質體溫度滅活條件的確定微生物原生質體滅活是利用物理或化學方法使其某一小部位的生理結構受到損傷,造成原生質體喪失獨立的再生活性,但決不是被徹底殺死,原生質體的染色體仍然保持復制與重組能力,融合后原生質體的細胞核和線粒體等仍具有轉化和互補功能。目前認為,溫度滅活原生質體的作用主要在細胞質中,使核糖體或核糖體RNA受到損傷,結果使細胞內的功能蛋白、酶蛋白的合成受到影響或使其變性失活,產生致死作用[16]。在
60℃條件下,大多數細菌都即可死亡,溫度超過65℃,原生質體容易黏結成團,不利于原生質體的融合[17]。
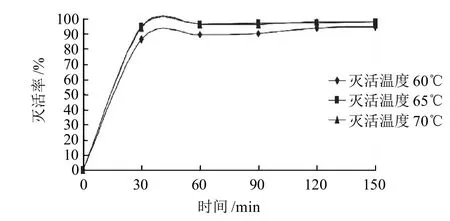
圖6 乳酸乳球菌FL菌株原生質體滅活溫度及時間對滅活率的影響Fig.6 Effect of inactivated time and temperature on inactivation rate of Lactococcus lactis FL protoplast
由圖6可知,在原生質體融合前,65℃對乳酸乳球菌FL菌株原生質體進行滅活120min,使其喪失再生的能力,同時用血球計數板觀察其沒有破裂,也沒有黏結成團。此滅活條件下,原生質體的滅活率最大可達到96.89%。被滅活的原生質體實際上起了遺傳物質的載體作用,與德氏乳桿菌FQ菌株融合后,融合菌株可能會具備雙親的優良性狀。
2.6 不同融合條件對原生質體融合率的影響
影響原生質體融合的因素有PEG6000處理時間、融合溫度和無機離子等。一般認為細菌原生質體融合處理時間為1~10min,融合溫度4、20℃比37℃好,同時融合過程中含有一定量的Ca2+和Mg2+會促進融合[18-19]。
由圖7可知,最適融合條件為:融合溫度為20℃,融合時間為5min,此條件下可得到最大的融合率2.72×10-6,相比文獻[20]單親滅活融合率2.74×10-7,融合率提高10倍左右。融合溫度為20℃時,細胞膜在較高溫度下流動性增加,PEG6000黏度下降,有利于原生質體的融合。隨著融合時間的增加,融合率逐漸降低,這可能是PEG6000處理時間過長,對原生質體有一定的毒性,從而導致其失活。
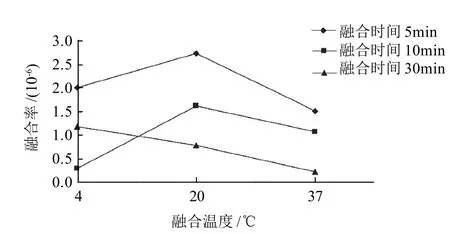
圖7 融合溫度和融合時間對原生質體融合率的影響Fig.7 Effects of temperature and time on protoplast fusion rate between Lactobacillus delbrueckii FQ and Lactococcus lactis FL
2.7 德氏乳桿菌FQ菌株、乳酸乳球菌FL菌株及融合子鏡檢結果

圖8 德氏乳桿菌FQ菌株(a)、乳酸乳球菌FL菌株(b)及融合子(c)的顯微鏡觀察圖(×400)Fig.8 Micrographs of Lactobacillus delbrueckii FQ (a), Lactococcus lactis FL (b) and their fusants (c) (×400)
用光學顯微鏡觀察親本菌株德氏乳桿菌FQ菌株、乳酸乳球菌FL菌株和融合子的細胞形態(圖8)可知,融合子的形態上發生了明顯的改變,變成短粗桿菌。
2.8 融合子和親本菌株對氧耐受性的比較
連續傳代5代后,篩選得到一株耐氧性融合子QL10。將融合子QL10和親本菌株在含終質量濃度為10μg/mL氯霉素的培養基中靜置和振蕩培養,在不含氯霉素的培養基振蕩培養作為對照,比較三者細胞密度的變化。由圖9可知,融合子QL10在3種培養方式下所得的細胞密度均高于親本德氏乳桿菌FQ菌株的細胞密度,且融合子能抗氯霉素,表明融合子的耐氧性能得到一定的改善。
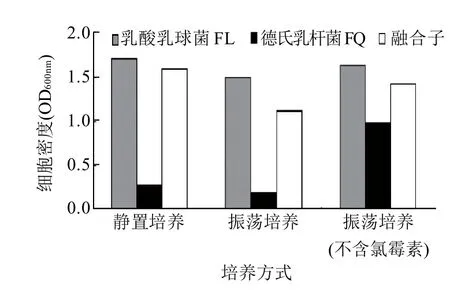
圖9 融合子和兩親本菌株細胞密度的比較Fig.9 Comparison of cell density among Lactobacillus delbrueckii FQ,Lactococcus lactis FL and their fusants under different incubation conditions
3 結 論
3.1 酶種類、酶質量濃度與德氏乳桿菌FQ菌株原生質體形成率及再生率密切相關。使用適當濃度的變溶菌素和溶菌酶進行酶解,可以促進酶解的進程,提高原生質體活性和再生率。制備德氏乳桿菌FQ菌株原生質體的最適酶解條件為:溫度37℃,在含有10μg/mL變溶菌素和1mg/mL溶菌酶的溶液中超聲處理90min,在此條件下原生質體形成率為99.99%,再生率為6.36%。德氏乳桿菌FQ菌株原生質體的最適再生培養基是RMⅡ。
3.2 乳酸乳球菌FL菌株生長培養基中添加1mg/mL甘氨酸,用終質量濃度為10mg/mL溶菌酶液37℃恒溫酶解處理90min,在此條件下原生質體形成率為99.97%。乳酸乳球菌FL菌株原生質體溫度滅活最適條件為65℃,滅活120min。
3.3 德氏乳桿菌FQ菌株和乳酸乳球菌FL菌株最適融合條件為:融合溫度20℃,融合時間5min,PEG6000 400g/L(含 0.02mol/L MgCl2和 0.01mol/L CaCl2),在此條件下融合率為2.72×10-6,該融合率是其他同類研究的10倍左右。
[1] GIVRY S, PREVOT V, DUCHIRON F. Lactic acid production from hemicellulosic hydrolyzate by cells of Lactobacillus bifermentans immobilized in Ca-alginate using response surface methodology[J]. World J Microbiol Biotechnol, 2008, 24(6): 745-752.
[2] WEE Y J, YUN J S, KIM D, et al. Batch and repeated batch production of L(+)-lactic acid by Enterococcus faecalis RKY1 using wood hydrolyzate and corn steep liquor[J]. J Ind Microbiol Biotechnol, 2006, 33(6):431-435.
[3] 楊登峰, 關妮, 潘麗霞, 等. 微生物發酵L-乳酸的研究進展[J]. 中國釀造, 2009(5): 1-3.
[4] CALABIA B P, TOKIWA Y. Production of D-lactic acid from sugarcane molasses, sugarcane juice and sugar beet juice by Lactobacillus delbrueckii[J]. Biotechnol Lett, 2007, 29(6): 1329-1332.
[5] WEE Y J, KIM J N, RYU H W. Biotechnological production of lactic acid and its recent applications[J]. Food Technol Biotechnol, 2006, 44(2): 163-172.
[6] VERINGA H A. Procedure for the preparation of D-lactic acid with Lactobacillus bulgaricus: US, 5322781[P]. 1994-06-21.
[7] 張蘭英, 劉娜, 孫立波, 等. 現代環境微生物技術[M]. 北京: 清華大學出版社, 2005: 45-69.
[8] 王玉華, 張桂榮, 劉景圣. 原生質體融合提高嗜酸乳桿菌耐酸及耐膽鹽能力[J]. 食品科學, 2006, 27(3): 96-99.
[9] SINGHVI M, JOSHI D, GAIKAIWARI S, et al. Protoplast formation and regeneration in Lactobacillus delbrueckii[J]. Indian J Microbiol,2010, 50(1): 97-100.
[10] 張莉滟, 陳林, 張德. 保加利亞乳桿菌原生質體的制備與回復研究[J]. 中國微生態學雜志, 2004, 16(2): 73-74.
[11] 曾獻春, 孟冬麗, 葛風偉. 乳酸菌原生質體融合篩選[J]. 中國釀造,2008(3): 26-28.
[12] 韓璞, 田洪濤, 苑社強. 羅伊氏乳桿菌原生質體的制備與再生條件的研究[J]. 中國食品學報, 2010, 10(1): 10-18.
[13] 王立梅, 齊斌. 基因組改組技術對L-乳酸產生菌耐熱性的影響[J]. 食品科學, 2008, 29(10): 395-398.
[14] 邱靜, 羅水忠, 姜紹通, 等. 高產L-乳酸米根霉的原生質體制備與再生條件研究[J]. 食品科學, 2011, 32(9): 174-178.
[15] 崔艷華, 張旭, 張蘭威. 德氏乳桿菌保加利亞亞種電轉化平臺的構建和優化[J]. 生物信息學, 2010, 8(3): 267-270.
[16] 陳志剛. 抗酸奶后酸化乳酸菌株的選育[D]. 烏魯木齊: 新疆農業大學, 2009.
[17] 裴曉林. 應用基因組改組技術選育L-乳酸高產菌株及其發酵工藝研究[D]. 長春: 吉林大學, 2007.
[18] 鄭重誼, 謝達平, 譚周進, 等. 影響微生物原生質體融合技術的因素[J]. 湖南農業科學, 2006(4): 35-38.
[19] 王登宇, 臧威, 孫劍秋, 等. 細菌原生質體融合育種技術及其應用進展[J]. 中國釀造, 2008(7): 1-6.
[20] 任柏林, 謝水波, 劉迎久, 等. 單親滅活檸檬酸桿菌與奇球菌原生質體融合[J]. 微生物學通報, 2010, 37(7): 975-980.
Optimization of Conditions for Protoplast Fusion between Single Inactivated Lactobacillus delbrueckii and Lactococcus lactis
LI Li,FANG Jie,HUANG Jie-jie,FU Rui-yan*
(College of Tea & Food Science and Technology, Anhui Agricultural University, Hefei 230036, China)
Single inactivated protoplast fusion technique was used for the fusion of Lactobacillus delbrueckii FQ with Lactococcus lactis FL. The protoplast formation, regeneration and fusion conditions were examined in this study. Results showed that the optimal enzymolysis conditions for L. delbrueckii FQ were ultrasound treatment for 90 min at 37 ℃ in the presence of 10 μg/mL mutanolysin and 1 mg/mL lysozyme. Under these conditions, the protoplast regeneration rate reached 6.36%. The protoplast formation rate of L. lactis FL was up to 99.97% after 90 min of treatment with 10 mg/mL lysozyme in the presence of 1 mg/mL glycine. L. lactis FL protoplasts showed an inactivated rate of 96.89% after 120 min of exposure to 65 ℃. The fusion rate between Lactobacillus delbrueckii FQ and Lactococcus lactis FL reached 2.72 × 10-6after 5 min of infusion at pH 6.5 in the presence of 400 g/L PEG 6000 (included 0.02 mol/L MgCl2 and 0.01 mol/L CaCl2).
single inactivation;Lactobacillus delbrueckii;Lactococcus lactis;protoplast fusion
Q813.2;Q939.117
A
1002-6630(2012)05-0193-06
2011-11-14
國家“973”計劃項目(2007CB707803);安徽農業大學穩定和引進人才科研資助項目
李麗(1987—),女,碩士研究生,主要從事乳品微生物生理方面的研究。E-mail:liliwxd312@163.com
付瑞燕(1976—),女,副教授,博士,主要從事食品微生物和生物技術研究。E-mail:fry@ahau.edu.cn
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
數學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
