
不同H5N1亞型高致病性禽流感病毒誘導(dǎo)IFN-α/β和MxA mRNA表達(dá)*
2012-11-06 06:13:40張永標(biāo)溫景蕓王艷紅焦培榮張桂紅辛朝安張扣興
中國病理生理雜志 2012年4期
關(guān)鍵詞:檢測
冼 盈, 朱 翔, 張永標(biāo), 溫景蕓, 王艷紅,焦培榮, 張桂紅, 廖 明, 辛朝安, 張扣興△
(中山大學(xué)附屬第三醫(yī)院 1感染科ICU,2急診科,3腫瘤科,廣東 廣州 510630;4華南農(nóng)業(yè)大學(xué)禽病教研室,廣東 廣州 510642)
1000-4718(2012)04-0708-06
2011-08-16
2012-01-09
廣東省自然科學(xué)基金資助項(xiàng)目(No.06034508);廣東省科技計(jì)劃資助項(xiàng)目(No.2008B030301302)
△通訊作者 Tel:020-85179179;E-mail:kxz6210@tom.com
不同H5N1亞型高致病性禽流感病毒誘導(dǎo)IFN-α/β和MxA mRNA表達(dá)*
冼 盈1, 朱 翔1, 張永標(biāo)2, 溫景蕓3, 王艷紅1,焦培榮4, 張桂紅4, 廖 明4, 辛朝安4, 張扣興1△
(中山大學(xué)附屬第三醫(yī)院1感染科ICU,2急診科,3腫瘤科,廣東 廣州 510630;4華南農(nóng)業(yè)大學(xué)禽病教研室,廣東 廣州 510642)
目的用兩株鵝源H5N1禽流感病毒(AIV)接種A549細(xì)胞,比較兩毒株在細(xì)胞中的增殖情況,以及病毒誘導(dǎo)細(xì)胞干擾素(INF)-α/β和黏病毒抗性蛋白A(MxA) mRNA表達(dá)的情況。方法將兩組病毒懸液接種A549細(xì)胞,按Reed-Muench法計(jì)算兩毒株的組織培養(yǎng)半數(shù)感染量(TCID50),PCR擴(kuò)增檢測細(xì)胞中病毒的HA和M1片段。Real-time PCR方法檢測細(xì)胞中IFN-α/β和MxA mRNA表達(dá)。結(jié)果兩株病毒均能感染A549細(xì)胞, AIV128誘導(dǎo)細(xì)胞的IFN-α/β和MxA mRNA表達(dá)均較AIV75高。結(jié)論H5N1亞型AIV75和AIV128毒株能在人肺泡癌上皮細(xì)胞A549中復(fù)制,細(xì)胞IFN-α/β和MxA mRNA的表達(dá)誘導(dǎo)依賴于病毒的復(fù)制。
禽流感病毒; A549細(xì)胞; 干擾素; 黏病毒抗性蛋白類
禽流感病毒(avian influenza virus, AIV)是禽類中分離出來的A型流行性感冒病毒各種亞型的總稱。目前,在世界各地分離的16個(gè)H亞型和9個(gè)N亞型禽流感病毒中,高致病性禽流感均由高致病性禽流感病毒(highly pathogenic avian influenza virus,HPAIV)H5或H7亞型病毒引起。1997年香港H5N1禽流感傳人事件的發(fā)生,改變了人們傳統(tǒng)上認(rèn)為禽流感不能直接傳人的概念。截止2010年8月31日,全世界已先后有505例人感染H5N1,導(dǎo)致300人死亡。AIV的致病性取決于病毒與宿主之間的相互作用,Ⅰ型干擾素(interferon,IFN)在宿主抗病毒感染中發(fā)揮著極其重要的作用,它先于機(jī)體的免疫應(yīng)答反應(yīng),是機(jī)體第一道病毒防御體系,也是影響流感病毒感染細(xì)胞的關(guān)鍵因素[1]。黏病毒抗性蛋白A(myxovirus resistance protein A,MxA)基因?qū)儆诟蓴_素刺激基因(interferon-stimulated genes,ISGs),在防御流感病毒機(jī)制中起到重要作用[2]。同時(shí),流感病毒的非結(jié)構(gòu)蛋白1(non-structural protein 1,NS1)能拮抗干擾素的產(chǎn)生,逃避干擾素的抗病毒作用[3]。目前,AIV致病機(jī)制還不清楚,可能是多基因共同作用的結(jié)果,研究機(jī)體對病毒的免疫反應(yīng)及AIV的NS1蛋白,有望進(jìn)一步揭示NS1蛋白的功能及其調(diào)節(jié)AIV致病性的機(jī)制。因此,我們設(shè)計(jì)本課題旨在建立人呼吸道上皮細(xì)胞模型,更好地觀察禽流感病毒H5N1不同毒株對細(xì)胞的致病性,并在細(xì)胞水平研究對禽流感病毒誘導(dǎo)產(chǎn)生的機(jī)體固有免疫反應(yīng)情況,從NS1基因方面尋找影響禽流感病毒致病力差異的原因,從而為研究NS1蛋白參與AIV致病的機(jī)制奠定基礎(chǔ)。
材 料 和 方 法
1材料
1.1病毒 A/Duck/Guangdong/75/2004(H5N1)(記為AIV75), A/Duck/Guangdong/128/2004(H5N1)(記為AIV128),由華南農(nóng)業(yè)大學(xué)禽病研究室分離、鑒定和保存。
1.2細(xì)胞 DF-1細(xì)胞(雞胚成纖維細(xì)胞)由華南農(nóng)業(yè)大學(xué)禽病研究室惠贈(zèng)、保存。人肺泡癌上皮A549細(xì)胞(CCL185,ATCC)為肺腺癌患者肺組織分離建立的、有肺泡上皮特征的細(xì)胞,為中山大學(xué)北校區(qū)實(shí)驗(yàn)動(dòng)物中心細(xì)胞庫保存。
1.3Real-time PCR試劑盒及引物設(shè)計(jì) SYBR Premix Ex Taq為寶生物工程(大連)有限公司產(chǎn)品,根據(jù)GenBank中的已知人β-actin、IFN-α/β和MxA全基因序列,及針對已知的AIV75、128兩毒株血凝素(hemagglutinin,HA)和M1序列,用Primer Primer 5.0軟件設(shè)計(jì)引物,由上海英駿生物技術(shù)有限公司合成,見表1。
2方法
2.1AIV75和AIV128接種 A549細(xì)胞 將2組病毒懸液用細(xì)胞維持液稀釋,用感染復(fù)數(shù)(multiplicity of infection,MOI)=2的感染劑量,接種于A549細(xì)胞已長成單層的6孔板中,每個(gè)病毒每個(gè)時(shí)點(diǎn)接種3個(gè)復(fù)孔,37℃、5%CO2培養(yǎng)箱中孵育2 h,棄病毒液,用PBS洗板2次,最后1次控干加入1 mL維持液,同時(shí)設(shè)正常細(xì)胞對照組,倒置顯微鏡下每8 h觀察細(xì)胞病變效應(yīng)(cytopathic effect,CPE)。分別在感染接種后8 h、16 h、24 h和48 h收集細(xì)胞及細(xì)胞上清液。

表1 實(shí)時(shí)定量PCR引物設(shè)計(jì)
2.2DF-1細(xì)胞內(nèi)檢測A549細(xì)胞中AIV的復(fù)制滴度 觀察實(shí)驗(yàn)結(jié)果后,將上述各孔A549細(xì)胞反復(fù)凍融3次,使細(xì)胞裂解,病毒釋放,5 000 r/min離心15 min,收集上清液.將收集到的上清液用細(xì)胞維持液做10倍稀釋為10-1~10-11,共10個(gè)稀釋度,加入DF-1細(xì)胞已長成單層的96孔板中,每個(gè)稀釋度接種4孔,每孔100 μL,37 ℃、5%CO2培養(yǎng)箱中吸附1 h,再添加維持液100 μL,置37 ℃、5%CO2培養(yǎng)箱中培養(yǎng),逐天觀察,記錄CPE情況,72 h取上清液測定病毒血凝效價(jià),并按Reed-Muench法[4]計(jì)算TCID50。RT-PCR 方法檢測A549細(xì)胞中病毒片段。
2.3熒光定量PCR檢測感染AIV后A549細(xì)胞中IFN-α/β和MxA mRNA的表達(dá) 分別收集AIV75、128感染后0 h、8 h、16 h、24 h和48 h的A549細(xì)胞,及收集不同濃度IFN-α2a作用后的A549細(xì)胞,提取細(xì)胞總RNA, 逆轉(zhuǎn)錄合成cDNA,用熒光定量PCR擴(kuò)增IFN-α、IFN-β和MxA片段,計(jì)算表達(dá)量。RT-PCR反應(yīng)組分,在7500 Fast Real-time PCR儀(ABI)中執(zhí)行以下程序:預(yù)變性95 ℃ 10 s,95 ℃ 5 s, 60 ℃15 s, 延伸72 ℃ 34 s, 共40個(gè)循環(huán),最后72 ℃ 34 s。擴(kuò)增完后,進(jìn)行熔解曲線分析,獲得熔解曲線。所用樣品檢測均設(shè)置陰性對照(無模板)以排除假陽性。每個(gè)樣本做3個(gè)復(fù)孔,獨(dú)立重復(fù)實(shí)驗(yàn)3次,同時(shí)擴(kuò)增人β-actin作為內(nèi)參照。反應(yīng)結(jié)束后,讀取各產(chǎn)物Ct值,并在溶解曲線上得到的特異性峰來判斷PCR擴(kuò)增的單一性。PCR產(chǎn)物取2 μL在1.0%的瓊脂糖凝膠(含0.5 mg/L EB)進(jìn)行電泳檢測以判斷擴(kuò)增產(chǎn)物是否為目的片段。采用相對定量法分別分析各基因相對相對mRNA表達(dá)水平,其計(jì)算公式為:相對mRNA表達(dá)量=2-ΔΔCt×100%,其中ΔCt=Ct(IFN-α/IFN-β/MxA)-Ct(β-actin),ΔΔCt=ΔCt-ΔCt(對照),其中病毒感染0 h和無IFN-α2a作用的A549細(xì)胞為對照組。
3統(tǒng)計(jì)學(xué)處理

結(jié) 果
1感染A549細(xì)胞后不同時(shí)點(diǎn)AIV75和AIV128的復(fù)制情況
正常A549細(xì)胞對照組細(xì)胞培養(yǎng)液清亮,細(xì)胞梭形,排列整齊致密,呈上皮形,胞漿清楚,見圖1A。A549細(xì)胞在接種兩株病毒72 h后均未出現(xiàn)CPE,見圖1B、C。加大病毒的感染劑量,仍無CPE。以上結(jié)果提示, AIV75、128兩毒株在A549細(xì)胞中不引起細(xì)胞病變。
AIV128感染A549細(xì)胞各時(shí)點(diǎn)的病毒上清液均引起DF-1細(xì)胞發(fā)生CPE,見圖2B;48 h時(shí)TCID50最高,與血凝效價(jià)基本一致,見表2。而 AIV75感染A549細(xì)胞各時(shí)點(diǎn)的病毒上清液均未能引起DF-1細(xì)胞發(fā)生CPE。
病毒RT-PCR檢測,在AIV75、128感染的A549細(xì)胞中,各時(shí)點(diǎn)均能檢出病毒HA和M1片段。AIV128株各時(shí)點(diǎn)PCR擴(kuò)增的病毒HA和M1片段,電泳條帶亮度均較AIV75強(qiáng),見圖3、4。
表2AIV128在A549細(xì)胞中不同時(shí)點(diǎn)的增殖結(jié)果
Table 2. The replication of AIV128 in A549 cells at different time points

Time(h)Virustiter(TCID50/mL)Hemagglutinationefficiency0008102.831∶1616103.771∶6424104.001∶6448104.671∶64

Figure 1. The growth of A549 cells infected by different AIV(×100).A:normal cultured A549 cells (adherent growth);B:A549 cells infected by AIV75;C:A549 cells infected by AIV128.
圖1不同禽流感病毒感染后A549細(xì)胞的生長情況

Figure 2. The growth of DF-1 cells infected by different AIV(×100).A:normal DF-1 cells (adherent growth);B: the supernatant with AIV128 was inoculated with DF-1 cells (cells floating in the culture medium, swollen and round).
圖2不同禽流感病毒感染后DF-1細(xì)胞的生長情況

Figure 3. The detection of HA and M1 gene fragments in A549 cells by RT-PCR after infected by AIV128 at diffe-rent time points.M: 2 000 bp ladder (molecular weight marker); 1: HA gene expression of mock-infected control cells; 2~5: HA gene of AIV128 which infected A549 cells 8 h,16 h,24 h and 48 h after infection; 6: M1 gene expression of mock-infected control cells; 5~10: M1 gene of AIV128 which infected A549 cells 8 h,16 h,24 h and 48 h after infection.
圖3RT-PCR檢測A549細(xì)胞中不同時(shí)點(diǎn)AIV128的HA和M1基因片段

Figure 4. The detection of HA and M1 gene fragments in A549 cells by RT-PCR after infected by AIV75 at different time points.M: 2 000 bp ladder (molecular weight marker); 1: HA gene expression of mock-infected control cells; 2~5:HA gene of AIV75 which infected A549 cells 8 h,16 h,24 h and 48 h after infection; 6: M1 gene expression of mock-infected control cells; 5~10: M1 gene of AIV75 which infected A549 cells 8 h,16 h,24 h and 48 h after infection.
圖4RT-PCR檢測A549細(xì)胞中不同時(shí)點(diǎn)AIV75的HA和M1基因片段
2病毒感染A549細(xì)胞后IFN-α和IFN-βmRNA的表達(dá)
在AIV75和AIV128感染的A549細(xì)胞,感染后8 h能檢測到IFN-α/β mRNA,表達(dá)量隨感染時(shí)間延長而增加,在24 h達(dá)到高峰。在每個(gè)檢測時(shí)點(diǎn),AIV128誘導(dǎo)細(xì)胞產(chǎn)生的IFN-β mRNA表達(dá)量均較AIV75高,見表3。
表3兩毒株各時(shí)點(diǎn)誘導(dǎo)A549細(xì)胞IFN-α、IFN-β和MxAmRNA的表達(dá)量
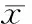

GroupIFN-βIFN-αMxAAIV75-8h2.67±0.350.37±0.0738.30±1.67AIV75-16h4.51±0.580.45±0.0339.75±1.85AIV75-24h31.12±2.354.50±0.2298.24±6.44AIV75-48h9.59±0.512.58±0.17114.07±6.67AIV128-8h5.97±0.31**0.56±0.05*103.65±12.94*AIV128-16h23.49±2.76☆☆2.24±0.14☆☆135.01±6.70☆☆A(yù)IV128-24h32.46±0.79##2.60±0.11##216.23±10.13##AIV128-48h29.79±0.84△△1.47±0.43△△401.36±42.91△△
*P<0.05,**P<0.01vsAIV75-8 h group;☆☆P<0.01vsAIV75-16 h group;##P<0.01vsAIV75-24 h group;△△P<0.01vsAIV75-48 h group.
3病毒感染A549細(xì)胞后MxAmRNA的表達(dá)
在AIV75和AIV128感染A549細(xì)胞后8 h能檢測到MxA mRNA表達(dá),表達(dá)量隨感染時(shí)間延長而增加,每個(gè)時(shí)點(diǎn)的MxA mRNA表達(dá)量比IFN-α/β mRNA表達(dá)量高。在感染后24 h IFN-α/β mRNA表達(dá)達(dá)到高峰時(shí),MxA mRNA表達(dá)量仍有增加,而在無病毒感染的細(xì)胞中沒有檢測到MxA mRNA表達(dá)。在每個(gè)檢測時(shí)點(diǎn),AIV128誘導(dǎo)細(xì)胞產(chǎn)生的MxA mRNA表達(dá)量均較AIV75高,見表3。
4不同濃度IFN-α2a誘導(dǎo)下A549細(xì)胞MxAmRNA的相對表達(dá)量
在沒有IFN-α2a處理的情況下,細(xì)胞不表達(dá)MxA mRNA, MxA mRNA表達(dá)水平隨著IFN-α2a作用濃度的增加而增加,見表4。
表4IFN-α2a對A549細(xì)胞MxAmRNA表達(dá)的誘導(dǎo)作用

IFN-α2a(103U/L)MxAmRNAexpression00108.5334±0.3800**1009.7884±0.7900**50023.0929±0.7200**
**P<0.01vsIFN-α2a 0 U/L.
討 論
A549細(xì)胞有著Ⅱ型肺泡上皮細(xì)胞的特征性形態(tài),有研究證實(shí)A549細(xì)胞可以作為研究Ⅱ型肺泡上皮細(xì)胞的標(biāo)準(zhǔn)細(xì)胞模型[5]。A549細(xì)胞源于人類肺泡上皮細(xì)胞,相對于其它傳代細(xì)胞,其分子結(jié)構(gòu)以及感染后細(xì)胞因子的分泌更接近活體感染試驗(yàn),因而檢測A549細(xì)胞感染H5N1亞型禽流感病毒,對于禽流感病毒跨種屬傳播的機(jī)制,以及感染人體后免疫系統(tǒng)的變化都將有很大的意義。本研究選取兩株H5N1亞型的禽流感病毒接種于A549細(xì)胞,運(yùn)用一系列病毒血清學(xué)試驗(yàn)方法及分子生物學(xué)方法,檢測病毒在細(xì)胞中的增殖能力。實(shí)驗(yàn)結(jié)果顯示, AIV128能感染A549細(xì)胞,隨著感染時(shí)間的增加,病毒復(fù)制增加。AIV75能感染A549細(xì)胞,但復(fù)制效率低下,未能引起DF-1細(xì)胞病變。說明AIV128在A549細(xì)胞中的增殖能力要強(qiáng)于 AIV75。
病毒的致病性很大程度上就是取決于宿主與病毒之間的相互作用。固有免疫反應(yīng)是病毒進(jìn)入宿主細(xì)胞遇到的第一道防線,被病毒感染的細(xì)胞可產(chǎn)生Ⅰ型干擾素,Ⅰ型干擾素能誘導(dǎo)細(xì)胞進(jìn)入一種抗病毒狀態(tài)。它與細(xì)胞表面的IFN-α/β受體結(jié)合后可激活JAK-STAT信號(hào)轉(zhuǎn)導(dǎo)通路,激活一系列抗病毒蛋白基因的轉(zhuǎn)錄從而抑制病毒的復(fù)制和擴(kuò)散[1, 6]。IFN通過信號(hào)通路誘導(dǎo)許多抗病毒蛋白的表達(dá)從而發(fā)揮其抗病毒作用,目前研究最多的主要有3個(gè)抗病毒通道:PKR(雙鏈RNA依賴蛋白激酶)、2’,5’-OAS(2’,5’-寡腺苷酸合成酶)、Mx蛋白通路。雖然PKR以及 2’,5’-OAS需要雙鏈RNA (double-strand RNA,dsRNA)才能發(fā)揮功能,但是它們處于干擾素介導(dǎo)的抗病毒反應(yīng)的下游,并不參與干擾素本身基因表達(dá)的調(diào)控過程。研究發(fā)現(xiàn):Toll樣受體(Toll-like receptor,TLR) 和 RIG-I/MDA5 細(xì)胞信號(hào)轉(zhuǎn)導(dǎo)通路在識(shí)別 dsRNA、誘導(dǎo)Ⅰ型干擾素大量表達(dá)的過程中發(fā)揮了舉足輕重的作用。TLR 是定位于細(xì)胞內(nèi)涵體膜上的模式識(shí)別受體(pattern recognition receptor,PRR),表達(dá)于免疫細(xì)胞,可識(shí)別進(jìn)入細(xì)胞囊泡的病毒dsRNA,誘導(dǎo)Ⅰ型 IFN的大量表達(dá),RIG-Ⅰ/MDA5是定位于細(xì)胞質(zhì)內(nèi)的 PRR,可識(shí)別細(xì)胞質(zhì)內(nèi)病毒復(fù)制產(chǎn)生的dsRNA,通過不依賴TLR 的方式誘導(dǎo)IFN合成。通常來說,疾病的嚴(yán)重程度和細(xì)胞因子水平強(qiáng)相關(guān)。1997年香港H5N1暴發(fā)的病人中發(fā)現(xiàn)高水平的血清IL-6、TNF-α、IFN-γ和sIL-2R[7]。在體外實(shí)驗(yàn)中,也表明了與H3N2和H1N1病毒比較,H5N1感染能大量地誘導(dǎo)前炎癥細(xì)胞因子,特別是TNF-α和IFN-β[8]。TLR 信號(hào)通路和 RIG-Ⅰ信號(hào)通路的過度激活會(huì)導(dǎo)致Ⅰ型 IFN 的過量表達(dá),大量Ⅰ型 IFN 對機(jī)體將造成嚴(yán)重的免疫損傷,因此,宿主細(xì)胞必然具有對 TLR 和 RIG-Ⅰ信號(hào)通路進(jìn)行下調(diào)的分子系統(tǒng),并且許多病毒都編碼特異的蛋白質(zhì)分子,抑制 TLR 和 RIG-Ⅰ介導(dǎo)的 IFN 的表達(dá),逃避宿主免疫系統(tǒng)的清除。病毒蛋白調(diào)控也是在多個(gè)水平進(jìn)行的。研究發(fā)現(xiàn),A型流感病毒編碼的非結(jié)構(gòu)蛋白 NS1 可與細(xì)胞內(nèi)的 PRR 競爭結(jié)合病毒產(chǎn)生的dsRNA,從而抑制人干擾素調(diào)節(jié)因子3(IRF3) 的激活,缺失 NS1 表達(dá)的A型流感病毒可大量誘導(dǎo) IFN 產(chǎn)生[9]。AIV75、 AIV128均能誘導(dǎo)A549細(xì)胞產(chǎn)生IFN-α/β mRNA,而正常對照組無IFN-α/β mRNA表達(dá),提示誘導(dǎo)細(xì)胞IFN-α/β mRNA表達(dá)依賴于病毒復(fù)制。AIV128誘導(dǎo)IFN-α/β mRNA表達(dá)量較AIV75高,提示不同毒株誘導(dǎo)IFN的作用是不一樣的,具有病毒株特異性。在一定感染劑量下,AIV128毒株隨著感染細(xì)胞時(shí)間延長,病毒復(fù)制增加,在細(xì)胞中復(fù)制即能誘導(dǎo)IFN產(chǎn)生,又能逃脫IFN的抗病毒作用,在細(xì)胞中有效增殖。不同流感病毒誘導(dǎo)和抑制Ⅰ型干擾素的能力是不同的[10]。與AIV75比較,AIV128在A549細(xì)胞中的復(fù)制水平高且誘導(dǎo)產(chǎn)生的IFN mRNA水平也增高,可能原因是:(1) 不同毒株對IFN的敏感性不同。AIV128在誘導(dǎo)細(xì)胞產(chǎn)生較高水平Ⅰ型干擾素時(shí),仍能在細(xì)胞內(nèi)有效增殖,與AIV75比較,干擾素對AIV128的抗病毒作用較弱。(2) 病毒NS1蛋白有表達(dá)缺陷,不能有效拮抗IFN的產(chǎn)生。(3) 除了NS1蛋白以外,病毒的其它基因也可能影響IFN的產(chǎn)生水平。有研究表明,含有A/Sydney/5/97HA和NA基因的流感重組病毒能在上皮細(xì)胞誘導(dǎo)TNF-α的生成[11], 因此,這些病毒基因片段很可能通過同一方式影響IFN的產(chǎn)生。
在小鼠實(shí)驗(yàn)中,已經(jīng)用基因重組的方法認(rèn)識(shí)了Mx蛋白能保護(hù)病毒感染。實(shí)驗(yàn)證明,MxA蛋白與RNP中的NP緊密結(jié)合后,MxA的構(gòu)型發(fā)生改變并被活化,活化的MxA發(fā)揮對病毒核衣殼的水解作用以阻止病毒對細(xì)胞的吸附與穿入,從而阻止病毒基因組在核內(nèi)的復(fù)制。與上述兩種抗病毒蛋白不同,MxA蛋白在沒有病毒感染的情況下是沒有持續(xù)、低水平表達(dá)的,低濃度的Ⅰ型干擾素即能誘導(dǎo)MxA基因轉(zhuǎn)錄,產(chǎn)生較高濃度的MxA蛋白。MxA蛋白在體外又具有廣譜抗病毒效應(yīng),特別對RNA病毒作用顯著,而且比IFN抗病毒作用更為直接。但是關(guān)于Mx蛋白在其它動(dòng)物和人體中作用的研究很少。H5N1禽流感病毒能夠誘導(dǎo)細(xì)胞產(chǎn)生大量IFN,并且逃脫IFN的抗病毒作用[12],盡管病毒阻斷了多個(gè)IFN誘導(dǎo)通路,但是并不完全,仍有IFN的分泌。I型或III型IFN信號(hào)通路能誘導(dǎo)MxA蛋白表達(dá)。人MxA基因的調(diào)節(jié)是受Ⅰ型干擾素嚴(yán)格控制的。我們實(shí)驗(yàn)結(jié)果顯示:感染后8 h能檢測到MxA mRNA表達(dá),感染后48 h仍有持續(xù)升高,表達(dá)量隨感染時(shí)間延長而增加。MxA mRNA與IFN-α/β-mRNA表達(dá)水平相平行,而表達(dá)量更高,表達(dá)半衰期更長,預(yù)先用IFN-α2a處理,可劑量依賴性地誘導(dǎo)MxA mRNA表達(dá),在缺乏IFN-α2a刺激的情況下,細(xì)胞不表達(dá)MxA mRNA(正常對照),與文獻(xiàn)報(bào)道的實(shí)驗(yàn)結(jié)果一致[13],提示MxA mRNA表達(dá)是受干擾素調(diào)控的,有較長的半衰期,少量的IFN-α/β即可刺激產(chǎn)生大量的MxA mRNA表達(dá),是評(píng)價(jià)Ⅰ型干擾素活性的可靠標(biāo)志,因此MxA抗病毒蛋白具有開發(fā)利用的潛能。
H5N1亞型AIV75和AIV128毒株能在人肺泡癌上皮細(xì)胞A549中復(fù)制,不同的毒株隨著感染時(shí)間的延長,病毒復(fù)制效率有所不同。兩毒株誘導(dǎo)細(xì)胞IFN-α/β和MxA mRNA表達(dá)依賴于病毒的復(fù)制。
[1] Samuel CE. Antiviral actions of interferons[J].Clin Microbiol Rev,2001,14(4):778-809.
[2] Haller O,Kochs G.Interferon-induced Mx proteins:dynamin-like GTPases with antiviral activity[J].Traffic,2002,3(10):710-714.
[3] Krug RM,Yuan W,Noah DL,et al. Intracellular warfare between human influenza viruses and human cells:the roles of the viral NS1 protein[J]. Virology,2003, 309(2):181-189.
[4] 殷 震,劉景華.動(dòng)物病毒學(xué)[M].第2版.北京:科學(xué)出版社,1997:329-330.
[5] Foster KA, Oster CG, Mayer MM. Characterization of the A549 cell line as a type II Pulmonary epithelial cell model for drug metabolism[J]. Exp Cell Res,1998,243(2):359-366.
[6] 陳敏慧, 徐 軍. H5N1禽流感病毒HA蛋白通過JAK3介導(dǎo)急性肺損傷的機(jī)制研究[J]. 中國病理生理雜志, 2010, 26(10): 2073.
[7] de Jong MD,Simmons CP, Thanh TT, et al. Fatal outcome of human influenza A (H5N1) is associated with high viral load and hypercytokinemia[J]. Nat Med, 2006, 12(10):1203-1207.
[8] Chan MCW,Cheung CY,Chui WH, et al. Proinflammatory cytokine responses induced by influenza A (H5N1) viruses in primary human alveolar and bronchial epithelial cells[J]. Respir Res,2005, 6:135.
[9] Talon J, Horvath CM, Polley R, et al. Activation of interferon regulatory factor 3 is inhibited by the influenza A virus NS1 protein[J].J Virol, 2000, 74 (17): 7989-7996.
[10]Hayman A,Comely S,Lackenby A, et al. Variation in the ability of human influenza A viruses to induce and inhibit the IFN-β pathway[J].Virology, 2006,347(1):52-64.
[11]Seo SH, Webby R,Webster RG. No apoptotic deaths and different levels of inductions of inflammatory cytokines in alveolar macrophages infected with influenza viruses[J].Virology, 2004,329(2):270-279.
[12] Seo SH, Hoffmann E, Webster RG. Lethal H5N1 influenza viruses escape host anti-viral cytokine responses[J]. Nat Med, 2002,8(9):950-954.
[13]Holzinger D,Jorns C,Stertz S,et al. Induction of MxA gene expression by influenza A virus requires type I or type III interferon signaling[J]. J Virol, 2007,81(14):7776-7785.
ThemRNAexpressionofIFN-α/βandMxAinducedbydifferentstrainsofH5N1highlypathogenicavianinfluenzaviruses
XIAN Ying1, ZHU Xiang1, ZHANG Yong-biao2, WEN Jing-yun3, WANG Yan-hong1, JIAO Pei-rong4, ZHANG Gui-hong4, LIAO Ming4, XIN Chao-an4, ZHANG Kou-xing1
(1IntensiveCareUnit,DepartmentofInfectiousDisease,2DepartmentofEmergency,3DepartmentofOncology,TheThirdAffiliatedHospital,SunYat-senUniversity,Guangzhou510630,China;4DepartmentofPoultryDiseases,SouthChinaAgriculturalUniversity,Guangzhou510642,China.E-mail:kxz6210@tom.com)
AIM: To compare the proliferative efficiency of two strains of H5N1 avian influenza viruses(AIV), AIV75 and AIV128, in A549 cells and the mRNA expression of interferon(IFN)-α/β and myxovirus resistance protein A (MxA) induced by the two strains of H5N1 AIV.METHODSA549 cells were infected with appropriate dilution of virus stock. The titers of avian influenza viruses were expressed as 50% tissue culture infectious dose (TCID50) by Reed-Muench method. Viral HA and M1 cDNA fragments were amplified by PCR. Real-time PCR were performed to detect the mRNA expression of IFN-α/β and MxA.RESULTSBoth AIV75 and AIV128 were able to infect A549 cells. The mRNA expression levels of IFN-α/β and MxA induced by AIV128 were dramatically higher than those induced by AIV75.CONCLUSIONA suitable epithelial cell line A549 can support the growth of AIV. The mRNA expression of IFN-α/β and MxA depends on the replication of the viruses.
Avian influenza virus; A549 cells; Interferon; Myxovirus resistance proteins
R363
A
10.3969/j.issn.1000-4718.2012.04.022
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
