蛋白激酶與植物滲透脅迫耐受研究進展
2012-12-02 04:50:24吳祖建
武夷科學 2012年0期
楊 靚,吳祖建
(福建農林大學福建省植物病毒學重點實驗室,福建福州350002)
Ferreira等在擬南芥中發現了P34cdc2蛋白激酶同系物之后(Ferreira et al.,1991),植物蛋白激酶的研究開始得到重視,雖然植物蛋白激酶的研究起步較晚,但是借助其他物種中蛋白激酶的研究平臺,其發展速度較快。目前的研究表明,蛋白激酶基因在植物中廣泛存在,據估計,蛋白激酶基因約占植物結構基因的1%-3%。植物中的蛋白激酶大都屬于絲/蘇氨酸類蛋白激酶,在植物體內是信號傳遞的重要載體(Bayer et al.,2012),它們通過催化功能蛋白的磷酸化,導致其構象發生改變,從而調節植物的生長、發育、自交不親和、雄性不育、抗逆和抗病等多種生理過程。
1 蛋白激酶的分類
對于激酶的分類,有不同的方法,其中一種以其底物蛋白被磷酸化的不同位點分為5類:(1)絲/蘇氨酸(Ser/Thr)蛋白激酶;(2)酪氨酸(Tyr)蛋白激酶;(3)組氨酸(His)蛋白激酶;(4)色氨酸(Try)蛋白激酶;(5)天冬氨酰基/谷氨酰基(Asp/Glu)蛋白激酶。目前植物中發現的蛋白激酶以前3類為主。而根據催化區氨基酸序列的相似性,又可將其分為五大組:AGC組、CaMK 組、CMGC 組、PTK 組和其它組,如下表(Bayer et al.,2012)。

表1 植物中蛋白激酶的分類Table 1 Classification of protein kinases in plant
2 蛋白激酶的結構
迄今為止發現的所有蛋白激酶,雖然其種類不同,作用方式和功能千差萬別,其作用底物也多種多樣,然而在它們的催化區域都具有相似的氨基酸序列。通過對蛋白激酶氨基酸序列的保守性分析發現,催化功能域由大約250-300個氨基酸殘基構成,包括12個亞區(Xu et al.,2011)。不同類型蛋白激酶的區別主要集中在Ⅵ、Ⅷ、Ⅸ亞域,例如:第Ⅵ區絲氨酸/蘇氨酸蛋白激酶的保守序列為:Asp-Leu-Lys-Pro-Glu-Asn,而酪氨酸蛋白激酶的保守序列則為:Asp-Leu-Arg-Ala-Asn或Asp-Leu-Ala-Ala-Arg-Asn。如果找到同一家族特有編碼基因的保守序列,就可以設計探針在cDNA或基因組文庫中篩選其他家族成員;也可以目標蛋白激酶編碼基因的保守序列,設計PCR引物,獲取其同源基因(鞏學千和陳受宜,1996)。
蛋白激酶對于底物蛋白中絲氨酸(Ser)、蘇氨酸(Thr)等氨基酸殘基的磷酸化是有選擇性的,往往只使特定部位的殘基磷酸化,這種選擇性取決于蛋白激酶可以識別的一個保守序列,蛋白激酶對靶位點的識別是由其附近的蛋白質一級結構所決定。遠距離的氨基酸及蛋白質的高級結構在此過程中的作用微乎其微,對于一種蛋白質,它所包含的磷酸化位點可能不止一個,它的作用可能在于不同位點發生磷酸化,使蛋白質產生不同的結構,而每種結構則對應于一種功能,從而使其功能更具多樣化。另外,有些蛋白質的磷酸化往往是多個激酶協同作用的結果。
3 植物滲透脅迫響應相關蛋白激酶
植物在生長發育過程中經常受到低溫、干旱等不良環境的脅迫。植物能夠快速的感知和主動防衛環境的變化,這一過程涉及到植物對脅迫信號的識別與傳導。大量的研究表明,蛋白激酶在植物信號識別與轉導過程中起著至關重要的作用(圖1)。與植物滲透脅迫信號傳遞關系最密切的主要有分裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK)、鈣依賴而鈣調素不依賴的蛋白激酶(calcium-dependent and calmodulin-independent protein kinase,CDPK)、受體蛋白激酶(receptor protein kinase,RPK)、核糖體蛋白激酶(ribosomal-protein kinase)、轉錄調控蛋白激酶(transcription-regulation protein kinase)。
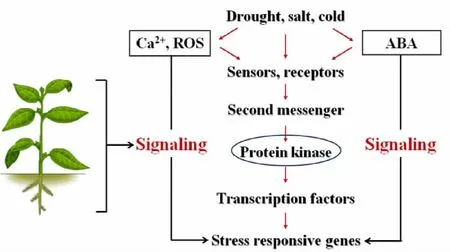
圖1 蛋白激酶參與的脅迫信號傳導通路Figure 1 Protein kinases as important key spot in stress signaling pathway
3.1 MAPK
3.1.1 植物MAPK級聯信號轉導系統 植物MAPK激酶級聯信號通路的研究在近幾年發展很快(Menke et al.,2004),現已在植物中發現了大量MAPK家族成員,而且在植物中識別出的MAPK數量比在動物和酵母中還多,不少編碼MAPK和一些編碼MAPKK與MAPKKK的基因也已從植物中分離到。MAPK是一類絲/氨酸蛋白激酶。廣泛存在于真核生物中。與一些其他的信號分子組成MAPK級聯途經。MAPK級聯途徑由三類激酶組成并通過依次磷酸化將逆境信號傳遞下去。即MAPKKK-MAPKK-MAPK,當MAPK被磷酸化后有3種可能的去向:(1)停留在細胞質中,激活一系列其他蛋白激酶;(2)在細胞質中使細胞骨架成分磷酸化;(3)進入細胞核,通過磷酸化轉錄因子,調控基因的表達。目前,在擬南芥中已經分離鑒定出幾條MAPK信號傳導通路(Cargnello and Roux,2011)。另外水稻等植物中MAPKKK、MAPKK或MAPK基因也已經被分離并驗證。這就為今后研究植物中的信號傳導機制打下了有力基礎。
3.1.2 植物MAPK的特點 植物MAPK與動物和酵母細胞中MAPK有相似的生化特點,多數是從胞質組分中分離到的,其分子量一般在44,000-48,000 Da之間,具有11個保守的蛋白激酶亞區。同動物和酵母一樣,MBP(脊髓堿性蛋白)也是植物MAPK的最適底物,這是MAPK的共有特征之一(Mizoguchi et al.,1997)。在其Ⅶ和Ⅷ亞區之間也存在一個催化區域。其中保守的TxY基序在此起關鍵作用。TxY基序在三維結構上定義“T環結構(T 1oop)”,T環是一個接近活化位點的表面環狀結構。它被認為是決定MAPK激酶活性的關鍵結構,在調節蛋白激酶活化狀態中起著非常重要的作用。與動物和酵母的不同之處在于:植物TxY中的X常為E或D,而動物和酵母中常為E、P或G,因此TDY基序為植物所特有。除了X氨基酸不同之外,不同植物MAPK在T環長度上也不同。TxY基序中x氨基酸的和T環氨基酸長度可以影響植物MAPK的底物特異性。并且T環長度在控制自主磷酸化上起關鍵性的作用。
3.1.3 植物MAPK與滲透脅迫 植物中已發現的多種MAPK除參與激素等的信號傳遞外,在脅迫信號傳遞尤其是滲透脅迫信號傳遞過程中也起著極其重要的作用。例如,擬南芥受到冷、水分脅迫的刺激會誘導出MAPK活性(B?gre et al.,1996),苜蓿葉片對干旱和冷處理的反應也涉及MAPK的激活(Jonak et al.,1996)。目前已從擬南芥中分離到兩個能被干旱、高鹽和低溫誘導的MAPK基因,一個是ATMPK3(編碼MAPK激酶),另一個是ATMEKK1(編碼MAPKKK激酶)。對它們的研究表明,MAPK級聯系統不僅在蛋白質水平上受到磷酸化和去磷酸化的調控,在轉錄水平也受到環境脅迫信號的誘導調控。在干旱、高鹽和低溫脅迫條件下,5 min內ATMPK3和ATMEKK1就被誘導表達,1 h內顯著增加,隨后表達持續增加,24 h后達到峰值(Mizoguchi et al.,1997)。ATMPK3基因啟動子克隆分析表明,TATA盒上游-10--6堿基處有一個與干旱、高鹽和低溫有關的DRE(dehydration responsive element)順式作用元件的核心序列(CCGAC)(Liu et al.,1998)。除了傳導滲透脅迫信號,一些MAPK也能感應滲透脅迫信號,正常情況下細胞壁和細胞質膜相接觸,質壁之間存在著相互作用。由于質壁的物理性質不同,細胞失水質膜收縮時質壁必然相互“撕扯”,這種“撕扯”實際是一種機械作用,而機械作用能被“機械感應器”識別。B?gre等證實這種“機械感應器”是一種MAPK,它在機械刺激后1 min就能被激活(B?gre et al.,1996)。
3.2 CDPK
3.2.1 植物中CDPK的功能與特點 Ca2+是植物中廣泛存在的第二信使,它在植物生長發育中都起到了重要作用。Ca2+所傳遞的信號可以被細胞內多種Ca2+感受器接受,并將之轉變成多種生物學反應(Saijo et al.,2002)。其中很重要的就是CDPK。CDPK是在豌豆中首先被報道的,并在大豆中首次得到了純化。
CDPK是一種在植物中廣泛存在的單鏈肽,它們在結構上具有明顯的特征,從N端到C端存在4個功能區:可變區、催化區、連接區和調控區。不同種類CDPK之間在可變區的差別較大,其他功能區則保守性較強。催化區由300多個氨基酸組成,具有典型的Ser/Thr蛋白激酶的催化保守序列。催化區的同源性較高,一般都在80%以上(Zhao et al.,2011)。連接區在各功能區中最為保守。是由20-30個氨基酸組成的,富含堿性氨基酸,它能以擬底物方式與催化區結合,在沒有Ca2+存在的情況下抑制催化區活性,故該區域又被稱為自抑制區(Hashimoto et al.,2012)。調控區即Ca2+結合區,為CDPK特有結構域,其保守性較差,其上有一段結構和功能類似于CaM的氨基酸序列,共有4個與Ca2+結合的EF手型結構,這是CDPK不依賴于CaM而對Ca2+高度親和的原因。
細胞內第二信使Ca2+的分布是嚴格區隔化的,正常情況下胞質中Ca2+濃度較低。當細胞受到刺激信號時,胞質中的Ca2+濃度發生短暫而復雜的變化,當Ca2+濃度達到一定濃度時便能和CDPK的調控區結合,解除連接區的自抑制作用使CDPK產生激酶活性,從而將信號級聯向下游傳遞。
3.2.2 CDPK與滲透脅迫 大量研究表明,很多植物中的CDPK與植物的脅迫反應有著密切關系。Sheen利用報告基因和效應基因共表達的方法,發現在玉米原生質體中CDPK1和CDPK1a能激活可被干旱和高鹽脅迫誘導的啟動子,去除CDPKl激酶區的突變體對脅迫和ABA刺激沒有反應(Sheen,1996)。此后一些學者從玉米、水稻、擬南芥等植物中發現了很多與滲透脅迫反應相關的 CDPK 基因(Urao et al.,1994;Saijo et al.,2000,2002;Sharma and Komatsu,2002)。其中,Saijo等報道來源于水稻的一種CDPK能被低溫、干旱和鹽脅迫激活,同時干旱和鹽脅迫能夠增強CDPK基因的表達,低溫則沒有這種效果,這說明在脅迫信號傳遞途徑中,干旱和鹽脅迫可能共用同一個類型的CDPK,而低溫脅迫信號傳遞用的是另一種類型(Saijo et al.,2000)。這些例子都說明,CDPK在滲透脅信號傳導過程中起著正調節因子的作用,能促進植物相關脅迫應答基因的表達。
3.3 其他激酶
3.3.1 CBK 人們首先是在蘋果中發現了CBK(Watillon et al.,1995),之后又在玉米、水稻、擬南芥中發現了多種 CBK(Wang et al.,2001;Zhang et al.,2002;Wang et al.,2004;Asano et al.,2011)。CBK的結構一般包括N端和C端可變區,中間還包括CaM結合區和催化區。CaM先與Ca2+結合再與CBK結合才能激活CBK的激酶活性,使信號向下傳遞。多種鈣調素結合蛋白激酶參與了植物對脅迫的反應(Yang and Poovaiah,2003;Giammaria et al.,2011)。另外,人們還在擬南芥、苜蓿和大豆中發現了鈣調素結合類受體胞質激酶(calmodulin-binding receptor-like cytoplasmic kinase,CRCK)(Yang et al.,2010),它的催化結構域類似于絲/蘇氨酸受體類激酶,不同的是它沒有跨膜和胞外結構域。CRCK與植物的抗脅迫反應有著密切聯系。
3.3.2 RPK 受體蛋白激酶位于細胞膜上,能夠感受外界刺激,并參與胞內信號轉導過程。它由胞外配體結合區(extracellular ligand-binding domain)、跨膜區(transmembrane domain)和胞質激酶區(cytosolic kinase domain)3個結構域組成。胞外配體結合區的功能是識別并結合配體感受外界信號,跨膜區則將被胞外配體識別的信號傳遞給胞質激酶區,而胞質激酶區能夠通過磷酸化作用將信號傳遞給下一級信號傳遞體。目前已在多種高等植物中分離出一系列受體蛋白激酶基因(Li and Chory,1997;Singla et al.,2008;Tanaka et al.,2012),并分析了它們的基因特征和表達特性。例如從擬南芥植株分離到一種類似受體的蛋白激酶基因RPK1,該基因編碼的氨基酸序列包含蛋白激酶中的11個保守亞區,屬絲氨酸/蘇氨酸蛋白激酶類型,具備胞外受體結構域、跨膜區和胞質激酶區等全部受體蛋白激酶的特征區。此外RPKl激酶的胞外區含有富亮氨酸重復序列(leucine-rich repeat sequence,LRR),該序列參與蛋白質-蛋白質間的相互作用,與感受發育和環境脅迫信號有關。表達特性分析表明,RPK1在植物的根、莖、葉和花中都能夠表達,干旱或高鹽(250 mmol/L NaCl)脅迫下,l h內RPK1的表達顯著增強,6 h后達到峰值,并至少在此后的18 h內保持穩定的高表達。Yang等從野生大豆中分離的受體類蛋白激酶GsAPK與ABA的信號通路相關,并且其激酶的酶活受ABA的特異誘導(Yang et al.,2012)。
3.3.3 轉錄調控蛋白激酶與核糖體蛋白激酶 Lee等(1999)在擬南芥的果實中分離出一個新的激酶類蛋白,其氨基酸序列與酵母的dbf 2基因產物不但有很高的同源性,而且通過酵母突變株的研究發現該基因能與酵母dbf2功能互補。故將之命名為At-DBF2,序列分析表明At-DBF2激酶屬于絲氨酸/蘇氨酸蛋白激酶類,包含有11個蛋白激酶的保守亞區。在酵母中,DBF2激酶是CCR4轉錄復合體的重要組分,它參與很多重要基因的轉錄,并且能影響許多轉錄因子的活性,所以推斷在擬南芥中它也是一種轉錄調控功能的蛋白激酶,位于信號傳遞的下游。高鹽、低溫及脫水均能很快誘導At-dbf2基因的表達,將該基因轉化酵母,At-dbf2超量表達的酵母DY株系抗高鹽、低溫及脫水能力均比對照有所提高。同時,分別將At-dbf2及其反義RNA序列轉入煙草BY2細胞系,使它們在煙草細胞中過量表達,經高鹽、干旱、低溫或高溫處理2周后發現,穩定表達At-dbf2基因的煙草細胞系抗脅迫能力明顯強于轉入反義RNA 的細胞系(Liu et al.,1997)。
Mizoguchi等(1995)在擬南芥中分離了2個基因ATPK6和ATPK19,這2個基因在序列上與動物的S6核糖體蛋白激酶有很高的同源性,在動物細胞中這類蛋白能夠通過磷酸化40S核糖體蛋白S6而激活蛋白質的合成。研究表明,ATPK6和ATPK19均能被干旱、低溫和高鹽誘導,由于S6激酶參與蛋白質的合成,所以ATPK6和ATPK19可能通過增加某些特定蛋白的合成使植物對外界脅迫作出反應。
4 問題與展望
同動物中蛋白激酶的研究相比,植物蛋白激酶的研究起步較晚,很多蛋白激酶還沒有被發現。大部分已克隆的蛋白激酶基因的功能也還沒有得到深入細致的研究,因而許多問題還不清楚,有些功能注釋只是參考動物中同源基因的功能注釋,然而,植物細胞中可能既存在類似于動物細胞的一些信號轉導因子及機制,同時又存在植物細胞特有的信號轉導因子及調節機制。參與環境脅迫信號的傳遞是蛋白激酶的重要功能之一。目前有關植物調控逆境脅迫和生長發育的研究重點逐漸從功能基因轉到蛋白激酶等調控基因的鑒定、功能分析及其調控機理等方面。但是植物的抗逆信號轉導途徑是一個相當復雜的過程,這一過程需要很多的受體蛋白、激酶、轉錄因子的協同作用,這就需要研究者們不僅僅克隆一些重要的蛋白激酶基因,更要從植物抗逆反應的整體全局來考慮,找到信號轉導途徑中每個關鍵的組分,為進一步闡明植物的生長發育和抗性機理奠定分子和生化基礎。
鞏學千,陳受宜.1996.蛋白激酶:一個飛速發展的領域.生物工程進展,16(1):11-15.
Asano T,Hakata M,Nakamura H,Aoki N,Komatsu S,Ichikawa H,Hirochika H,Ohsugi R.2011.Functional characterisation of OsCPK21,a calcium-dependent protein kinase that confers salt tolerance in rice.Plant Molecular Biology,75(1-2):179-191.
Bayer RG,Stael S,Rocha AG,Mair A,Vothknecht UC,Teige M.2012.Chloroplast-localized protein kinases:a step forward towards a complete inventory.Journal of Experimental Botany,63(4):1713-1723.
B?gre L,Ligterink W,Heberle-Bors E,Hirt H.1996.Mechanosensors in plants.Nature,383(6600):489-490.
Cargnello M,Roux PP.2011.Activation and function of the MAPKs and their substrates,the MAPK-activated protein kinases.Microbiology and Molecular Biology Reviews,75(1):50-83.
Ferreira PC,Hemerly AS,Villarroel R,Van MM,Inzé D.1991.The Arabidopsis functional homolog of the p34cdc2 protein kinase.Plant Cell,3(5):531-540.
Giammaria V,Grandellis C,Bachmann S,Gargantini PR,Feingold SE,Bryan G,Ulloa RM.2011.StCDPK2 expression and activity reveal a highly responsive potato calcium-dependent protein kinase involved in light signalling.Planta,233(3):593-609.
Hashimoto K,Eckert C,Anschütz U,Scholz M,Held K,Waadt R,Reyer A,Hippler M,Becker D,Kudla J.2012.Phosphorylation of calcineurin B-like(CBL)calcium sensor proteins by their CBL-interacting protein kinases(CIPKs)is required for full activity of CBL-CIPK complexes toward their target proteins.Journal of Biological Chemistry,287(11):7956-7968.
Jonak C,Kiegerl S,Ligterink W,Barker PJ,Huskisson NS,Hirt H.1996.Stress signaling in plants:a mitogen-activated protein kinase pathway is activated by cold and drought.Proceedings of the National Academy of Sciences of the United States of America,93(20):11274-11279.
Lee JH,Van Montagu M,Verbruggen N.1999.A highly conserved kinase is an essential component for stress tolerance in yeast and plant cells.Proceedings of the National Academy of Sciences of the United States of America,96(10):5873-5877.
Li J,Chory J.1997.A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction.Cell,90(5):929-938.
Liu HY,Toyn JH,Chiang YC,Draper MP,Johnston LH,Denis CL.1997.DBF2,a cell cycle-regulated protein kinase,is physically and functionally associated with the CCR4 transcriptional regulatory complex.EMBO Journal,16(17):5289-5298.
Liu Q,Kasuga M,Sakuma Y,Abe H,Miura S,Yamaguchi-Shinozaki K,Shinozaki K.1998.Two transcription factors,DREB1 and DREB2,with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought-and low-temperature-responsive gene expression,respectively,in Arabidopsis.Plant Cell,10(8):1391-1406.
Menke FL,van Pelt JA,Pieterse CM,Klessig DF.2004.Silencing of the mitogen-activated protein kinase MPK6 compromises disease resistance in Arabidopsis.Plant Cell,16(4):897-907.
Mizoguchi T,Hayashida N,Yamaguchi-Shinozaki K,Kamada H,Shinozaki K.1995.Two genes that encode ribosomalprotein S6 kinase homologs are induced by cold or salinity stress in Arabidopsis thaliana.FEBS Letters,358(2):199-204.
Mizoguchi T,Ichimura K,Shinozaki K.1997.Environmental stress response in plants:the role of mitogen-activated protein kinases.Trends in Biotechnology,15(1):15-19.
Saijo Y,Kinoshita N,Ishiyama K,Hata S,Kyozuka J,Hayakawa T,Nakamura T,Sanders D,Pelloux J,Brownlee C,Harper JF.2002.Calcium at the crossroads of signaling.Plant Cell,14Suppl:S401-S417.
Saijo Y,Hata S,Kyozuka J,Shimamoto K,Izui K.2000.Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants.Plant Journal,23(3):319-327.
Sharma A,Komatsu S.2002.Involvement of a Ca2+-dependent protein kinase component downstream to the gibberellin-binding phosphoprotein,RuBisCO activase,in rice.Biochemistry Biophysics and Reserach Communications,290(2):690-695.
Sheen J.1996.Ca2+-dependent protein kinases and stress signal transduction in plants.Science,274(5294):1900-1902.
Singla B,Khurana JP,Khurana P.2008.Characterization of three somatic embryogenesis receptor kinase genes from wheat,Triticum aestivum.Plant Cell Report,27(5):833-843.
Tanaka H,Osakabe Y,Katsura S,Mizuno S,Maruyama K,Kusakabe K,Mizoi J,Shinozaki K,Yamaguchi-Shinozaki K.2012.Abiotic stress-inducible receptor-like kinases negatively control ABA signaling in Arabidopsis.Plant Journal,70(4):599-613.
Urao T,Katagiri T,Mizoguchi T,Yamaguchi-Shinozaki K,Hayashida N,Shinozaki K.1994.Two genes that encode Ca2+-dependent protein kinases are induced by drought and high-salt stresses in Arabidopsis thaliana.Molecular and General Genetics,244(4):331-340.
Wang L,Liang S,Lu YT.2001.Characterization,physical location and expression of the genes encoding calcium/calmodulin-dependent protein kinases in maize(Zea mays L.).Planta,213(4):556-564.
Wang Y,Liang S,Xie QG,Lu YT.2004.Characterization of a calmodulin-regulated Ca2+-dependent-protein-kinase-related protein kinase,AtCRK1,from Arabidopsis.Biochemical Journal,383(Pt1):73-81.
Watillon B,Kettmann R,Boxus P,Burny A.1995.Structure of a calmodulin-binding protein kinase gene from apple.Plant Physiology,108(2):847-848.
Xu F,Meng T,Li P,Yu Y,Cui Y,Wang Y,Gong Q,Wang NN.2011.A soybean dual-specificity kinase,GmSARK,and its Arabidopsis homolog,AtSARK,regulate leaf senescence through synergistic actions of auxin and ethylene.Plant Physiology,157(4):2131-2153.
Yang T,Poovaiah BW.2003.Calcium/calmodulin-mediated signal network in plants.Trends in Plant Science,8(10):505-512.
Yang L,Ji W,Zhu YM,Gao P,Li Y,Cai H,Bai X,Guo DJ.2010.GsCBRLK,a calcium/calmodulin-binding receptorlike kinase,is a positive regulator of plant tolerance to salt and ABA stress.Journal of Experimental Botany,61(9):2519-2533.
Yang L,Ji W,Gao P,Li Y,Cai H,Bai X,Chen Q,Zhu YM.2012.GsAPK an ABA-activated and calcium-independent SnRK2-type kinase from G.soja mediates the regulation of plant tolerance to salinity and ABA stress.PLoS ONE,7(3):e33838.doi:10.1371/journal.pone.0033838.
Zhang L,Liu BF,Liang S,Jones RL,Lu YT.2002.Molecular and biochemical characterization of a calcium/calmodulin-binding protein kinase from rice.Biochemical Journal,368(Pt1):145-157.
Zhao R,Wang XF,Zhang DP.2011.CPK12:A Ca2+-dependent protein kinase balancer in abscisic acid signaling.Plant Signal And Behavior,6(11):1687-1690.
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
