新城疫病毒在全球的變異研究
2012-12-10 08:33:12黎先偉譯
獸醫導刊 2012年5期
黎先偉譯
自從1926年發現新城疫病毒以來,已經鑒定出Ⅰ型病毒有9個基因型和Ⅱ型病毒有10個基因型,代表著一種形式多樣且不斷演變的病毒。從全球的家畜流行病中一種強毒的基因型的出現及觀察新城疫強弱毒的基因組序列年與年之間的變化暗示著新城疫病毒不同的基因型在全球不同的地理位置同時演變著。這巨大的基因組多樣性可能會使更多的各種各樣的鳥類易于受到新城疫病毒的感染以及以經常遷徙的野鳥為儲藏所。新城疫病毒基因組的多樣性增加了診斷失敗的可能性,結果導致不明感染。不斷地進行流行病學監測以及積極主動地鑒定流行株以確保免疫和PCR試劑能有效地鑒別世界范圍內新城疫病毒的流行株。一種新型基質聚合酶復合試驗能夠檢測目前全球大多數新城疫病毒的流行株。通過改進的融合蛋白技術目前已經用于鑒定美國和歐洲強毒力的鴿新城疫病毒流行株。在新分離的毒株中含有未知的基因序列,讓目前成熟的隨機引物測序的方法應用整合到診斷方法上來。此外,對于目前新城疫病毒的分型方法是不適當的。在本文中,我們回顧有關新城疫病毒進化的分子流行病學和目前診斷出現的問題。同時,解釋說明為什么需要在客觀的標準下建立一個新的分型方法。
一、新城疫(ND)
新城疫是由新城疫病毒感染引起的,可能能感染大多數種類的鳥類,對于易感的家禽具有高度的感染性和致死性。新城疫是最重要的家禽傳染病之一,因為該病廣泛存在與全世界,而且有可能造成巨大的經濟損失。該病至少發生于亞洲、非洲、美洲等六大洲,在許多國家也是地方性動物病。在美國,新城疫強毒株被歸類為特殊的病原體,這些毒株在家禽中并不普遍,有這些毒株引起的疾病通常被稱為外來的新城疫(END)(USDA,2006)。在世界各地,由新城疫強毒株暴發的新城疫一直威脅著養禽業。在2008年臨床確診暴發新城疫的國家有多米尼加共和國、伯利茲城、秘魯、芬蘭和日本。或許一些暴發新城疫卻沒有報道的國家遍及非洲。在亞洲新城疫強毒株也很普遍。
在雞新城疫的不同菌株中,臨床表現有明顯的差異。強毒株的感染有3種明確的臨床分型:(1)嗜內臟速發型新城疫,以急性致死性敗血性感染,病死雞有明顯的消化道出血性病變;(2)嗜神經速發型新城疫,呈急性致死性感染,以呼吸道和神經癥狀為特征,但通常很少發生內臟病變;(3)中發型新城疫,引起呼吸道和神經癥狀癥狀,但致死性較低。在一些家禽新城疫強毒株普遍存在的國家需要加強疾病監測和管制,以免對商業生產和家禽及其產品的國際貿易帶來嚴重影響。低毒力的新城疫病毒通常被認為是弱毒的,而且普遍存在于美國。另外,弱毒力的毒株被分型為緩發型新城疫,一般只引起輕度的呼吸道癥狀。弱毒株也可以被歸類為無癥狀腸型,主要引起腸道感染,無呼吸道癥狀。
二、新城疫病毒(NDV)
新城疫病毒作為禽副黏病毒I型是副粘病毒科禽腮腺炎病毒屬的成員。它是單股負鏈不分節段的有囊膜的RNA病毒。新城疫病毒基因組包括六個基因,分別編碼六個結構蛋白:NP蛋白、P蛋白、M蛋白、F蛋白、HN蛋白和L蛋白。在P基因的轉錄過程中會出現mRNA的編輯現象,因而可能會產生額外的蛋白質,如V蛋白及W蛋白。雖然新城疫病毒毒力是多基因相關的,但是F蛋白的裂解位點卻是毒力改變的主要因素。
三、新城疫病毒分類
不同基因型的禽I型副粘病毒在世界各地流行。盡管所有的新城疫病毒都屬于禽副粘病毒I型,但是也存在抗原和遺傳多樣性。目前對于新城疫病毒主要有兩種分類方法。一種分類方法是有Aldous組提出的將新城疫病毒分為6個譜系以及13個亞譜系,后來還增加了額外的三個亞譜。另一種分類方法是將新城疫病毒分成兩系即I系和II系,其中I系可以分為9個基因型,II系分為10個基因型。因為這兩種系統分類方法都是基于類似的基因組信息,所以他們之間的差異只是名義上的。1B圖中顯示的是II類病毒,顯示了兩個系統分類方法之間最顯著的差異。例如,基因III型,IV型,V型和VIII型組成了Aldous系統中的譜系3.兩種系統同時命名存在的問題在表1中說明,表1中已經列出不同基因型間的平均進化關系分歧。大部分基因型間的進化距離在0.041到0.181之間。但是在基因VI型中,VIc 和VIf之間的進化距離在0.061而基因III型和IV型之間的進化距離在0.041,說明基因VI型可能需要進一步分類。進化分析表明不同基因型之間的進化距離存在一個較大的變化。例如,在基因IX型中的進化距離是0.004而在基因III型中則是0.085。區分基因型在距離上的巨大差別以及其他標準的匱乏都表明現在需要一個科學會議去討論和建立一個客觀的標準以期得到廣泛的認可和一致的分類。
在本文中,我們將用第二種分類方法。禽副粘病毒I型基因組至少有三種長度,15186,15192和15198個核苷酸。I系病毒是來源于雞和形目鳥類的弱毒株。I系病毒的基因組最長為15198個核苷酸。這些病毒至少包括9個基因型。這些病毒可能分布在世界各地的野生鳥類,并且經常在活禽市場的樣本中分離到。
II系病毒研究的比較詳細,包括10個基因型。基因型包括早期的I, II, III, IV 和IX型,含有15186個核苷酸。后來出現的病毒V, VI,VII, VIII, and X型含有15192個核苷酸。除1998至2000導致澳大利亞新城疫暴發的病毒以外,其他II系病毒中的基因I型毒株都是弱毒株,其中很多經常被用來做弱毒疫苗。II類病毒中的基因II型病毒包括很多在世界范圍被用作疫苗候選株的弱毒株,如LaSota,B1和VG/GA株。它還包括一個強毒株TXGB,這個毒株在美國相當于一個標準強毒株,被用來評價新城疫疫苗的有效性。20世紀四五十年代,在美國上述提到的疫苗都可以很有效的抵抗強毒株如TX/GB株和堪薩斯州的曼哈頓株的攻擊,然而基因II型的強毒株在美國已不再流行。基因III型毒株大多是1960年前在日本分離的,但也有1969年和1985年在臺灣分離的以及1990年在津巴布韋分離的。基因IV型病毒是1970年前的歐洲主要的分離株。
基因V, VI, VII, 和VIII型病毒是全球范圍流行的主要的基因型,并且只有強毒株。基因V型病毒于1970年出現在南美和中美,導致了同年歐洲新城疫的暴發。這些病毒也造成了在北美佛羅里達州(1971年,1993年)和加利福尼亞州(1971年,2002年)新城疫的暴發,而且依然在墨西哥流行。基因VI型病毒出現在20世紀60年代,在亞洲直到1985年基因VII型病毒逐漸普及之前它都是主要的病毒基因型。基因VI型病毒可以進一步分為Via到VIg,其中VIb大部分分離自鴿。
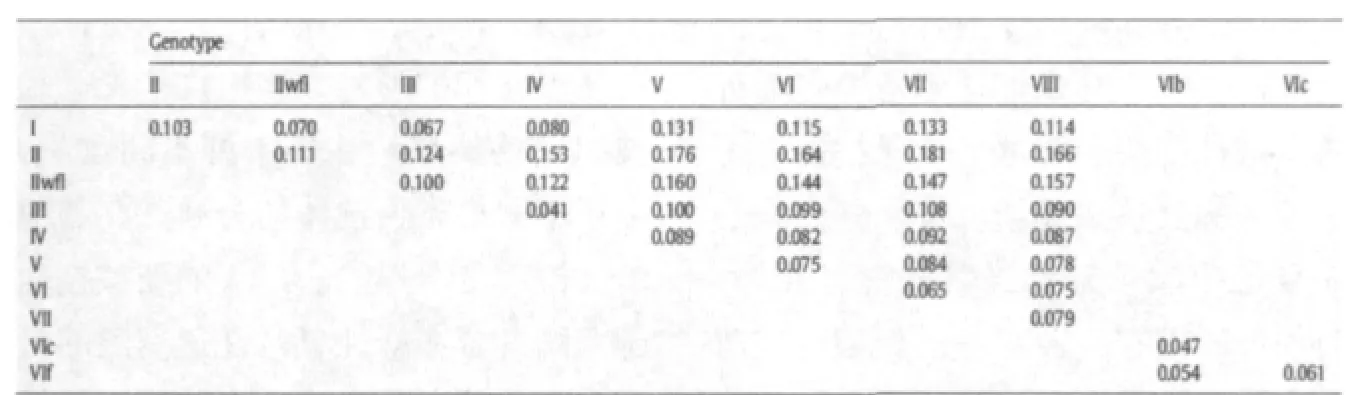
表1 評估不同基因型間的平均進化分歧
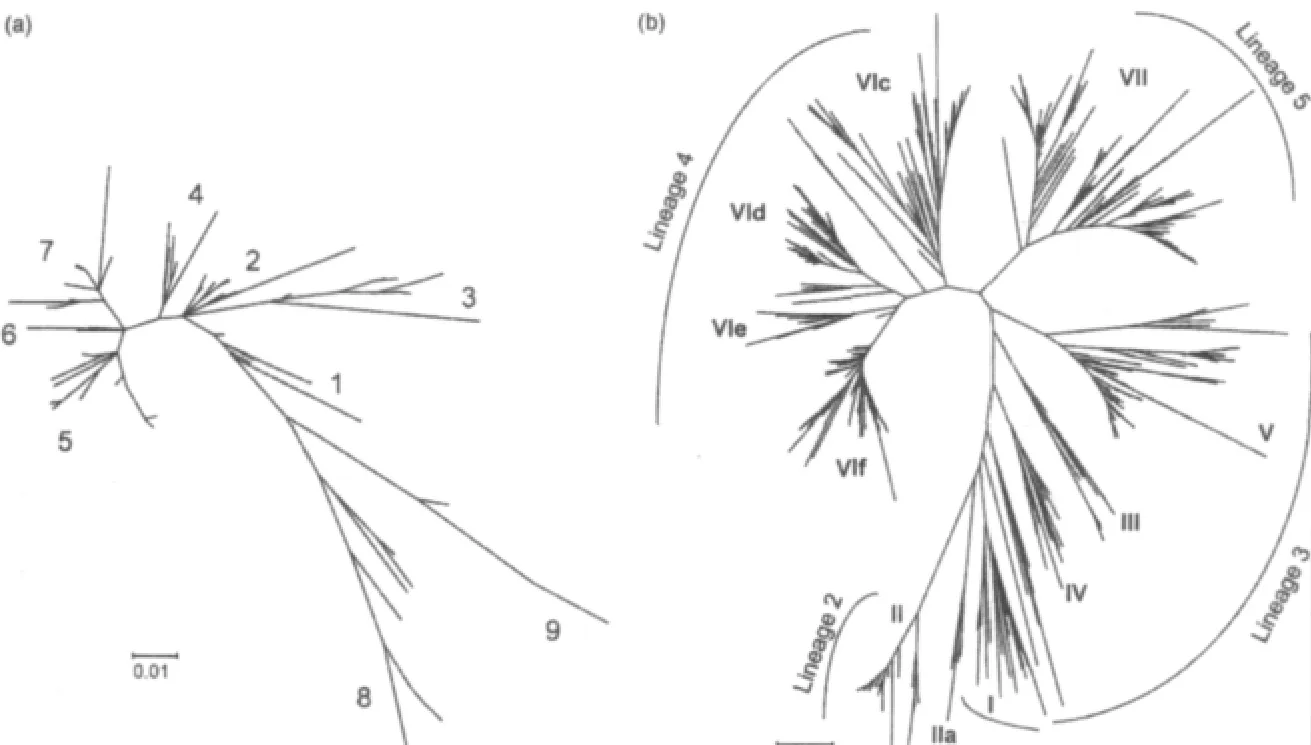
圖1 新城疫病毒系統進化樹 (a)為I類病毒(n=216);(b)為II類病毒(n=956)
基因VII型最初分為兩個亞基因型,VIIa和VIIb。VIIa的代表病毒主要在90年代出現在遠東,后來蔓延至歐洲和亞洲。VIIb的代表病毒出現在遠東并且蔓延至南非。這兩個基因亞型又可以進一步分為以中國,哈薩克斯坦,南非分離株為代表的VIIc,VIId,VIIe以及以非洲分離株為代表的VIIf,VIIg和VIIh。VIII型病毒在20世紀60年代流行于南非,并且在東南亞持續流行。基因IX型病毒是一個特殊的群體,它包括1948年從中國分離的第一個強毒株,而且在中國時有分離。基因X型病毒在1969年和1981年全部分離自臺灣。除了基因IV型以外(1989年以后再未報道過),大部分基因型現在仍在流行。
四、低致病性新城疫病毒的進化
關于低致病性NDV的進化鮮為人知,因為大部分的報道都是關于強毒株的研究。在全球范圍內來看,Ⅰ系的所有基因亞型和Ⅱ系的基因Ⅰ型和基因Ⅱ型的低致病性新城疫病毒通常是在家禽和野鳥體內分離得到。在美國,家鴨中的強毒株不是地方流行的,接種疫苗主要是為了保護由本土所流行的低致病性新城疫病毒引起的呼吸道疾病帶來的經濟損失。此外,假如禽群沒有一致接種活新城疫疫苗或者另一種疫苗干擾新城疫疫苗的作用,那么那些疫苗保護不佳的禽只幾天后就會發呼吸道疾病。疫苗的反作用被稱作“滾動效應”。病毒在禽只間傳播時,呼吸道疾病的傳播可能是接種疫苗失敗后的傳播比接種疫苗初更為嚴重。雖然在滾動效應中的相關病毒被假設是最初的疫苗,與這些毒株的基因特性無關,但是包含來自野鳥的低致病性新城疫病毒的可能性不能被忽視。
野禽感染的大量的不同型的無毒力病毒,這些病毒通常不會引起家禽的臨床癥狀。近來對分離自美國野禽和LBM的無毒力NDV進行系統發生分析發現大約70%的毒株屬于I系,30%的毒株屬于II系。幾乎一半的I系毒株來自馴服野鴨。在這個單一物種,代表了I系基因型的九分之六,這表明某些物種極易受到感染新城疫病毒,并可能成為儲存器。野禽是II系病毒的攜帶者但沒有致命的病毒存在。這些II系病毒分為基因I型和倆個新的亞型:IIa亞型和Ia亞型。Ia亞型病毒是與澳大利亞流行株相關毒株,包括來自澳大利亞1998年新城疫大暴發的強毒株。IIa亞型賭注是以前分離自阿根廷和西班牙的毒株。沒有野禽的病毒是通常用的疫苗株,B1和LaSota株。考慮到每年在美國和世界各地應用活病毒的家禽數量是令人驚訝的。如以前描述的通過比較單克隆抗體在不同的LaSota株之間的粘合模式來觀察II系病毒不同強毒株的抗體水平。例如,當一個小組的九種單克隆抗體對抗來自美國1986至1999年從野鴨分離出的58種病毒的測試,揭示七個不同的結合模式。考慮這項研究,只研究來自雁形目和!形目,新城疫病毒已經被紀錄的感染至少241種不同禽類,存在不同單克隆抗體結合模式在其他禽類似乎是合理的。廣大的新城疫病毒系統發育多樣性顯示在這有限的野生鳥類種群中,產生更多基因型的潛力沒有被發現。
一般而言,1系病毒屬于野禽病毒。在美國和亞洲發現的很多種類的新城疫病毒,在野生鳥類中相關病毒基因的存在表明,流行病存在這些種群之間。美國從1986到2005年收集的菌株發現 LBM 菌株和WS菌株之間有密切的親緣關系。
香港在 2003到2005年之間,從LBM家禽中分離出了Ⅱ類病毒。這表明這些病毒在LBM鳥類和野生鳥類世界中傳播。2003年分離自安大略12周齡的家養火雞的2系1a基因亞型病毒經鑒定與2001年分離自美國新澤西海鳥毒株的親緣關系最近,可能表明這些病毒種群在野鳥和家禽間傳播。
2系1a亞型和1系病毒已經在美國的LBM雞中分離到。分離自野禽的新城疫病毒被預言是低致病性病毒,在家禽中存在與低致病性到高致病性基因變異的可能。至少1系中的一株強毒株的病原學鑒定曾引發疫情大暴發。那些已在野禽和LBM被發現的相似亞型的強毒株已經復原。1990年愛爾蘭暴發的疫情是由1系強毒株引起,1998-2000年澳大利亞暴發的疫情是由2系Ia亞型強毒株引起。正是理論上的弱毒株在愛爾蘭水禽中的循環突變為強毒株并形成了引起1系強毒的準毒株。對來自澳大利亞疫情暴發時別的2系Ia亞型病毒進行基因組分析,也可提供弱毒株已經擁有隨著時間變為強毒株的潛力,也證明了是在天然病毒的融合蛋白的切割位點發生了改變導致毒力增強。
有些地方性弱毒病毒只要兩個位點發生變異就可形成強毒。除野外條件下,有些弱毒NDV在實驗條件下具有形成強毒株的能力。這些因素可能會增加疾病暴發的危險,對疾病的預測,需要研究進化對NDV的基因組所產生的影響。
(1)融合基因中極少部分的核酸發生改變就足以促使DNV由弱毒變為強度。
(2)在自然界中,家禽可能容易接觸到含有弱毒病毒的大型流動水庫。
五、新城疫病毒強毒株和新城疫疫苗在進化過程中的作用
雖然新城疫病毒強毒株最有可能的儲存庫在已接種過疫苗的家禽中,但有證據表明野鳥可能是中發型病毒的自然儲存庫。新城疫病毒強毒株屬于基因Ⅴ型從1975年到2008年已經在一種雙脊突的鸕鶿上分離到,而他們涉嫌與較早之前暴發的新城疫有關。1981年第一次從鴿子體內分離到鴿副粘病毒Ⅴ型強毒株在雞上臨床表現為嗜神經性,而且繼續在世界范圍內鴿屬動物內傳播。雖然在較早已有記錄,鴿子內分離到的是嗜內臟型的新城疫病毒強毒株,典型的鴿子分離株在引起嗜神經性基因Ⅵb型的位點發生變異,擁有特異的單克隆抗體結合模式,而且經常不能使雞紅細胞凝集。其他強毒株偶然會從進口的熱帶鳥類體內分離到,但在經確認在其他物種的尸體中也有病毒庫的存在。事實上,經研究表明一些野鳥并不是天然的新城疫強毒株,而是出口以前在檢驗站感染而來。
從鸕鶿和鴿子體內分離到新城疫強毒株被認為是中發型新城疫,因為它們在雞體內的ICPI變化范圍在0.7至1.5之間,而且在家禽內通常不會引起顯著的疾病。然而,在最近美國指出ICPI指數大于0.7或氨基酸序列與新城疫強毒株相一致就認為是強毒株,并把它們歸類為極好的因子,以遵循OIE和EU的標準。雖然鸕鶿和鴿子的病毒不斷進化和表現基因型也在不斷改變,但從野鳥獲得的樣品中新城疫病毒的毒力并沒有發生顯著的變化。
1系弱毒株和2系中發型新城疫以及鸕鶿和鴿子中的基因Ⅴ或Ⅵ所獨有的優勢與目前流行于接種過疫苗的家禽內嗜神經性新城疫病毒(Ⅱ類,基因Ⅴ-Ⅹ)相比,暗示著疫苗的免疫作用可能對多種形式的新城疫強毒株進行選擇。盡管加強疫苗的作用,家禽中新城疫強毒株不確定的存留在亞洲、非洲和美國中部的一些國家也會有周期復發的現象。目前家禽生產系統的結合特征可能包括:(1)宿主基因同質性,(2)高密度的飼養(增加了動物之間的接觸,有利于高致病性的毒株傳播并覆蓋弱毒株);(3)過強的免疫程序提供了病毒在免疫壓力下的選擇作用和在一些發展中國家試驗不合理的免疫程序,都有可能使強毒株不斷的進化。然而,良好的監測研究并沒有切實開展,用以證明免疫接種在新城疫病毒強毒株進化過程中所起到的作用。
系統進化樹和抗原的熒光分析在疫苗和目前流行的強毒株之間可能可以促進新城疫病毒強毒株的進化。許多研究已經證明目前使用的疫苗可以預防疾病,但并不能阻止病毒的脫落。另外有研究表明,使用基因重組疫苗可以顯著地減少病毒脫落的出現。
目前已經發現可以逃避免疫接種的變異毒株。在韓國不同地點和時間點分離到的變異新城疫病毒株的頻率從2002年的21%增加到2003年至2006年的85.7%。不幸的是,所分析的病毒株的總量較少(56個樣品),而使流行病學特性并不確定。這些作者進一步鑒定HN基因上一個變異表位,發展了以LaSota病毒株為骨架利用變異的HN基因病毒株的重組疫苗。使用含有相匹配的滅活的疫苗毒株并指出與商業疫苗在對抗致病率和死亡率方面有相同的保護作用。而且,這類重組疫苗在預防產蛋雞疾病方面有更佳的效果。可惜的是,在本次研究中并沒有對病毒的脫落進行評價。
基因變異也可能出現在墨西哥。新城疫病毒在墨西哥第一次被分離到的時間是在1946年。在最近一次報道的由于新城疫強毒株引起的大暴發是在2000年,當時屠殺了1.36×107只野鳥和對93個養殖場進行了隔離(OIE,2003)。這次新城疫大暴發以后,對受影響的區域加強免疫接種的程序。對墨西哥病毒的融合基因全基因編碼區域與已經公布的其他已知的Ⅱ類新城疫病毒基因型進行系統進化的比較,結果表明1998年到2006年從墨西哥分離到的病毒株歸類為Ⅱ類基因Ⅴ型病毒(Perozo et al.,2008)。然而,分析本次研究病毒分離的清晰地系統進化分為早期(1998-2001年)和近期(2004-2006年)兩組,遵循ICPI和MDT的結果。這次結果顯示1998年至2001年期間存在著一些明顯的選擇壓力導致病毒出現新的組別。1998-2001年的分離株與1996-2001年墨西哥報道的其他病毒具有高度的一致性,達到98%,但與最近的分離株只有93%~94%的一致性。可惜的是,沒有研究去描述兩組病毒抗原差異的特性。
有證據表明新城疫疫苗在一些特殊的條件下會沒有效果。另外,自2005年以來,已經發現在后備種雞持續的暴發新城疫強毒株感染,免疫接種良好,具有高水平新城疫抗體的禽類出現產蛋水平下降(Castilla and colleagues, 2009 ANECA abstract),這與韓國出現的產蛋問題非常相似(Cho et al., 2008b)。沒有臨床征兆,通過病毒的分離和組織病理學確證為新城疫強毒株。
六、結論
由于新城疫病毒具有高度的感染性以及臨床癥狀與高致病性禽流感相似,準確的監控和快速的診斷鳥類的感染對于任何控制和撲滅程序具有至關重要的作用。積極主動的檢測野鳥和家禽生產基地應該增加我們對NDV病毒的進化的理解。活禽交易市場有特別重要的意義,因為那里使多種不同品種的鳥類混合在一起,并提供了一個儲存物種的環境,例如,水禽跟鵪鶉飼養在附近。在檢測新城疫強毒株方面仍具有許多挑戰。快速的檢測和區分具有挑戰,因為新城疫病毒存在顯著的基因多樣性和經常不能區分疫苗和地方性毒株。另外,分析系統進化樹揭示新城疫病毒的弱毒株和強毒株仍在不斷進化。我們需要改良監測方法,增加對野鳥和國內家禽新城疫病毒的生態學的理解和特異的診斷方法來識別新城疫病毒所有不同的基因型。
