對(duì)嗜壓蛋白氨基酸壓力不對(duì)稱(chēng)指數(shù)的修訂
2012-12-27 06:55:26張光亞
張光亞
(華僑大學(xué) 化工學(xué)院,福建 廈門(mén) 361021)
對(duì)嗜壓蛋白氨基酸壓力不對(duì)稱(chēng)指數(shù)的修訂
張光亞
(華僑大學(xué) 化工學(xué)院,福建 廈門(mén) 361021)
選取嗜壓菌最多的4種希瓦氏菌屬(Shewanella)微生物共計(jì)473對(duì)同源蛋白,統(tǒng)計(jì)了21 662個(gè)氨基酸突變位點(diǎn),在此基礎(chǔ)上計(jì)算氨基酸壓力不對(duì)稱(chēng)指數(shù)(PAI),并將其同氨基酸70種理化性質(zhì)進(jìn)行相關(guān)性分析.結(jié)果表明:色氨酸和異亮氨酸的嗜壓性能最強(qiáng),而脯氨酸、甘氨酸和天冬酰胺的嗜壓性能最弱,這與之前的報(bào)道存在較大差異.此外,70種氨基酸理化性質(zhì)中有13種同PAI存在較強(qiáng)的相關(guān)性,其中僅有氨基酸的無(wú)規(guī)則傾向因子同PAI呈負(fù)相關(guān),表明高的PAI值不易形成卷曲.由于展示的PAI值更適合于處于低溫環(huán)境下的嗜壓微生物,因此對(duì)以前的PAI值進(jìn)行了修正.
嗜壓微生物;壓力不對(duì)稱(chēng)指數(shù);氨基酸;傾向因子;適壓機(jī)制
對(duì)多數(shù)海洋微生物尤其是深海微生物而言,在高壓下生存尤為重要,相比于其他極端微生物,人們對(duì)嗜壓微生物的關(guān)注較少[1].同溫度一樣,壓力也是嚴(yán)重影響生物體內(nèi)分子體系的熱動(dòng)力學(xué)參數(shù),但壓力對(duì)生物分子的影響一直沒(méi)有得到應(yīng)有的重視[2].近年來(lái)研究發(fā)現(xiàn),一些蛋白質(zhì)(或者酶)在高壓條件下表現(xiàn)出良好的特性,如在最適溫度下,生物分子的催化能力明顯上升,酶活力提高了約25%,或者其催化底物的特異性發(fā)生改變[3-4].基因組學(xué)和蛋白質(zhì)組學(xué)的迅猛發(fā)展,為了解嗜壓微生物內(nèi)生物分子體系提供了全新的視角.通過(guò)對(duì)一些已完成基因組測(cè)序的嗜壓微生物基因組或蛋白質(zhì)組的分析表明,它們?cè)谀さ鞍住⒑粑湣NA損傷修復(fù)、核糖體結(jié)構(gòu)及組裝等方面都存在明顯特點(diǎn)[5].然而,有關(guān)嗜壓蛋白質(zhì)中氨基酸使用的偏好與其理化性質(zhì)之間聯(lián)系的研究很少,但在嗜熱[6]、嗜冷[7]等極端蛋白質(zhì)中卻得到廣泛研究.Di Giulio首次利用蛋白質(zhì)組信息,系統(tǒng)比較了Pyrococcus嗜壓和非嗜壓微生物的141個(gè)直系同源序列(orthologs),結(jié)果發(fā)現(xiàn)側(cè)鏈較小精氨酸(Arg),甘氨酸(Gly),纈氨酸(Val)和天冬氨酸(Asp)在嗜壓蛋白質(zhì)中含量很高,而一些側(cè)鏈較大的氨基酸如酪氨酸(Tyr)和色氨酸(Trp)則含量很低.在此基礎(chǔ)上,他提出了氨基酸壓力不對(duì)稱(chēng)指數(shù)(pressure asymmetry index,PAI),即PAI與氨基酸分子量呈反比,而同其極性呈正比[8].然而,文獻(xiàn)[9-10]的研究中卻發(fā)現(xiàn),嗜壓蛋白中嗜壓氨基酸并不一定含量更高,相反非嗜壓氨基酸則明顯上升.本文從Shewanella兩對(duì)嗜壓和非嗜壓微生物中得到了473對(duì)同源蛋白,共統(tǒng)計(jì)了185 621個(gè)氨基酸,計(jì)算其氨基酸壓力不對(duì)稱(chēng)指數(shù),發(fā)現(xiàn)與Di Giulio的結(jié)果存在較大差異,并將其同氨基酸的70種理化性質(zhì)進(jìn)行了相關(guān)性分析,找出了與壓力顯著相關(guān)的氨基酸理化特性.
1 材料和方法
1.1 微生物的選取
目前發(fā)現(xiàn)的嗜壓微生物多為細(xì)菌,且同時(shí)也為嗜冷菌,Shewanella是目前分布于深海中最廣泛的嗜壓微生物[11].因此,共選取了Shewanella兩對(duì)微生物全蛋白質(zhì)組序列,它們分別為:S.benthica KT99和S.frigidimarina NCMIB400,以及S.violacea DSS12和S.sediminis HAW-EB3,前者均為嗜壓微生物,后者為均非嗜壓[12-13].
S.benthica KT99為專(zhuān)性嗜壓菌,分離自海平面下9 856m深處,其最適生長(zhǎng)溫度和壓力分別為10℃和50MPa,基因組G+C的摩爾分?jǐn)?shù)為46%;S.violacea DSS12分離自海平面下5 110m深處,最適生長(zhǎng)溫度和壓力分別為8℃和30MPa,基因組G+C的摩爾分?jǐn)?shù)為44.7%;S.frigidimarina NCMIB400為革蘭氏陰性菌,最適生長(zhǎng)溫度為10℃,基因組G+C的摩爾分?jǐn)?shù)為41.6%,S.sediminis HAW-EB3為兼性厭氧菌,最適生長(zhǎng)溫度為10℃,基因組G+C的摩爾分?jǐn)?shù)為46.1%.這4種微生物來(lái)源于同一個(gè)屬,親緣關(guān)系較近;其最適生長(zhǎng)溫度接近,基因組G+C的摩爾分?jǐn)?shù)也很接近,從而最大程度的降低了其他因素對(duì)同源蛋白氨基酸選擇的影響,使生長(zhǎng)壓力成為影響氨基酸使用偏好的最重要因素.
1.2 同源序列的獲取及序列比對(duì)
以上4種微生物全蛋白質(zhì)組序列來(lái)源于 UniProtKB(http://www.uniprot.org/)[14].為了減少信息冗余,剔除了所有長(zhǎng)度小于100個(gè)氨基酸的序列,因?yàn)樗鼈兇蠖酁槠瑪啵╢ragment)或者部分(partial).采用BlastP程序[15]選取直系同源序列(orthologs),其E 值為1.0×10-10,并找出90%長(zhǎng)度覆蓋范圍內(nèi),同一性(identity)大于30%的同源蛋白序列.樣本1包含222對(duì)同源序列,樣本2包含251對(duì),共計(jì)473對(duì),946條.該樣本量足夠用于探測(cè)嗜壓蛋白在氨基酸使用上的某種趨勢(shì).
獲得同源序列后,采用Clustal X軟件[16]將473對(duì)同源蛋白兩兩進(jìn)行比對(duì)[8],以統(tǒng)計(jì)同源蛋白在某些位點(diǎn)氨基酸發(fā)生突變的情況.
1.3 壓力不對(duì)稱(chēng)指數(shù)的計(jì)算
在正常情況下,兩種氨基酸發(fā)生互相突變的幾率應(yīng)該是50∶50的比例,即各占50%的幾率.若受到某種因素影響,就會(huì)偏離這種幾率,偏離程度越大,表明該因素的影響越大.壓力不對(duì)稱(chēng)指數(shù)(PAI)就是基于這種理念進(jìn)行計(jì)算的.文中的計(jì)算方法同文獻(xiàn)[8],均采用John H.McDonald等提出的方法[17-18].其計(jì)算式為
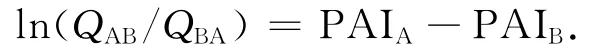
式中:A為非嗜壓蛋白中某位點(diǎn)的氨基酸;B為嗜壓蛋白質(zhì)對(duì)應(yīng)位點(diǎn)的氨基酸;QAB為非嗜壓蛋白中氨基酸A出現(xiàn)的位點(diǎn)在嗜壓蛋白相應(yīng)位點(diǎn)被B氨基酸取代的比例;QBA的含義則相反.
需要指出的是,最近,McDonald對(duì)算法進(jìn)行了修訂,將指數(shù)值的中點(diǎn)數(shù)從1改為0,他認(rèn)為這能更敏感的反應(yīng)數(shù)據(jù)的特性[6].因此,本文計(jì)算的PAI值不能直接和之前結(jié)果比數(shù)值大小,而主要是考察20種氨基酸PAI值的排序情況.
1.4 氨基酸理化性質(zhì)及相關(guān)性分析
文中使用了70種氨基酸的理化性質(zhì),如親疏水性、極性、分子量、等電點(diǎn)、分子表面可及性、分子體積、熱容、吉布斯自由能、形成二級(jí)結(jié)構(gòu)的傾向性、電荷分布、側(cè)鏈相互作用參數(shù)、平均柔性指數(shù)、長(zhǎng)距離相互作用等.這些數(shù)據(jù)來(lái)源于 AA Index數(shù)據(jù)庫(kù)(http://www.genome.jp/aaindex/)[19].
將計(jì)算所得的20種氨基酸的壓力不對(duì)稱(chēng)指數(shù)同其70種理化性質(zhì)分別進(jìn)行相關(guān)性分析,軟件為STATISTICA 9.0(30天完整功能試用版),選取相關(guān)系數(shù)較大,顯著性較高的理化性質(zhì)進(jìn)行分析[8].
2 結(jié)果和分析
2.1 壓力對(duì)蛋白質(zhì)氨基酸使用偏性的影響
比較了473對(duì)嗜壓和非嗜壓同源蛋白中20種氨基酸的替換情況,結(jié)果如表1所示.該樣本中共統(tǒng)計(jì)了185 621個(gè)氨基酸,其中有21 662個(gè)氨基酸位點(diǎn)發(fā)生了替換,樣本量足夠用于統(tǒng)計(jì)氨基酸使用上可能存在的某種趨勢(shì).
從表1可看出:在20種氨基酸中,有6種氨基酸沒(méi)有出現(xiàn)差異有統(tǒng)計(jì)學(xué)意義的氨基酸使用偏性,占總數(shù)量的30%,它們分別是C(半胱氨酸)、D(天冬氨酸)、H(組氨酸)、M(甲硫氨酸)、W(色氨酸)和Y(酪氨酸);而出現(xiàn)差異有統(tǒng)計(jì)學(xué)意義最多的氨基酸是K(賴(lài)氨酸),其次I(異亮氨酸)和N(天門(mén)冬酰胺).
經(jīng)統(tǒng)計(jì),在380個(gè)不同氨基酸對(duì)中,有12對(duì)存在差異有統(tǒng)計(jì)學(xué)意義的氨基酸使用偏性,結(jié)果如表2所示.表2中:Forward表示非嗜壓蛋白突變?yōu)槭葔旱鞍祝籖everse表示嗜壓蛋白突變?yōu)榉鞘葔旱鞍?從表2可知:差別最大是VI(纈氨酸-異亮氨酸),在嗜壓蛋白中有1 062個(gè)位點(diǎn)是Ile,而在非嗜壓蛋白對(duì)應(yīng)的位點(diǎn)卻為Val;同時(shí),嗜壓蛋白中有763個(gè)位點(diǎn)是Val,而對(duì)應(yīng)的位點(diǎn)在非嗜壓蛋白卻是Ile,即嗜壓蛋白有1 062個(gè)Val突變成Ile,僅有763個(gè)從Ile突變成Val,二者之間差異達(dá)極有統(tǒng)計(jì)學(xué)意義的水平.
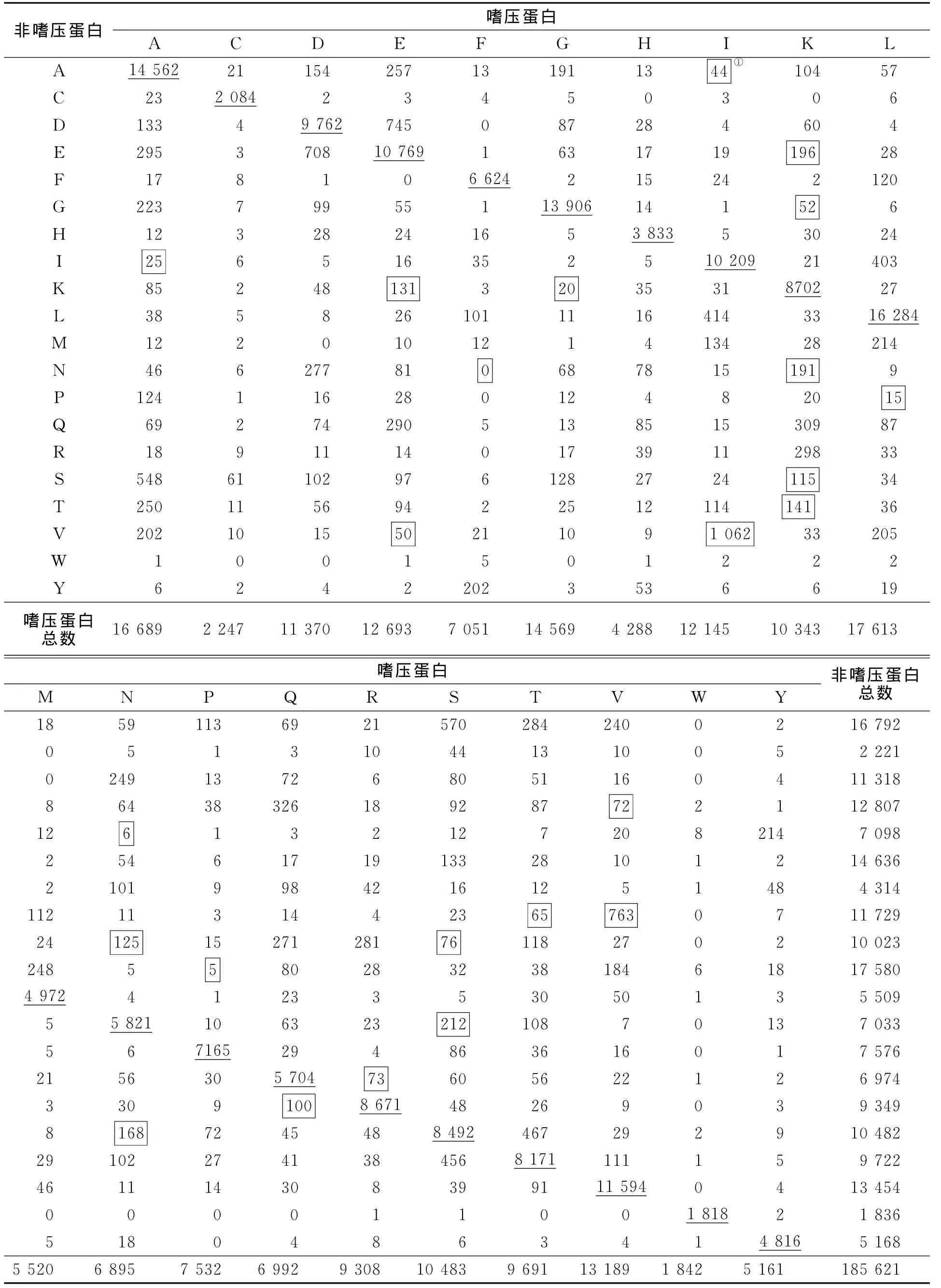
表1 嗜壓和非嗜壓菌473對(duì)同源蛋白中氨基酸替換的矩陣Tab.1 Amino acid substitution matrix in the 473orthologous proteins from piezophiles and non-piezphilies
然而,僅僅從表2中這些差異有統(tǒng)計(jì)學(xué)意義的氨基酸對(duì)仍難以得到一個(gè)完整的兩種類(lèi)型蛋白中氨基酸使用偏性的模式.因此,需要統(tǒng)計(jì)其他所有氨基酸向某種氨基酸突變的總趨勢(shì).例如:非嗜壓蛋白中除W(色氨酸)以外的其他19種氨基酸向嗜壓蛋白中W突變的總數(shù)量,以及嗜壓蛋白中除W(色氨酸)以外的其他19種氨基酸向非嗜壓蛋白中W突變的總數(shù)量.通過(guò)這兩組數(shù)據(jù)的比較,找出其突變總趨勢(shì),并在此基礎(chǔ)上計(jì)算W的PAI值.
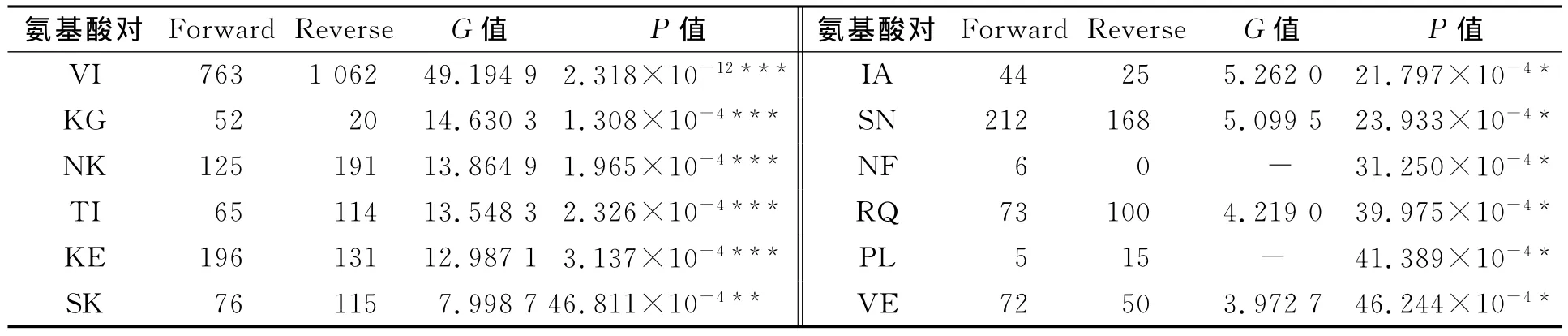
表2 473對(duì)同源蛋白中替換差異有統(tǒng)計(jì)學(xué)意義的氨基酸對(duì)Tab.2 Most statistical bias amino acid pairs in 473homologous proteins
經(jīng)統(tǒng)計(jì)及計(jì)算后,20種氨基酸替換總體數(shù)據(jù)及其PAI值如表3所示.PAI值越大,表明該氨基酸更容易出現(xiàn)在嗜壓蛋白中,其嗜壓性能越強(qiáng).由表3可知:Trp(色氨酸,W)和Ile(異亮氨酸,I)的嗜壓性能最強(qiáng),其次分別為L(zhǎng)ys(賴(lài)氨酸,K)、Cys(半胱氨酸,C),Met(甲硫氨酸,M)和 Leu(亮氨酸,L);而Pro(脯氨酸,P)、Gly(甘氨酸,G)和Asn(天冬酰胺)則為最不嗜壓氨基酸.這與之前報(bào)道差異明顯.

表3 20種氨基酸的PAI值Tab.3 PAI values of the 20amino acids
例如:文中Trp為嗜壓性能最強(qiáng)的氨基酸,而在之前研究中它確是屬于最不嗜壓的氨基酸之一;而文中最不嗜壓的Gly在之前研究中卻是位于第2嗜壓的氨基酸.Trp側(cè)鏈具有芳香結(jié)構(gòu),其分子中的π電子能夠與帶正電的離子如鈉離子和鉀離子相互作用,被稱(chēng)為“陽(yáng)離子-p”相互作用,這對(duì)維系蛋白分子結(jié)構(gòu)穩(wěn)定性非常重要[20].Ile的側(cè)鏈為分支的異丙基,能更有效提高蛋白分子表面的柔性,增加其穩(wěn)定性[21],故其嗜壓性能也較強(qiáng).Pro的剛性較強(qiáng),過(guò)多的Pro容易導(dǎo)致蛋白分子剛性太強(qiáng),不利于其穩(wěn)定性[22].Gly雖側(cè)鏈很小,易占據(jù)蛋白分子內(nèi)外空間,方便其折疊,但它卻是非極性氨基酸,而極性氨基酸之間及其與水分子間較容易形成極性相互作用,這對(duì)維持蛋白質(zhì)在高壓條件下的穩(wěn)定性非常重要[5].Chou等[23]的研究表明,Asn,Gly和Pro很容易出現(xiàn)在轉(zhuǎn)角和無(wú)規(guī)則轉(zhuǎn)曲中,是強(qiáng)烈的轉(zhuǎn)角和轉(zhuǎn)曲形成子;過(guò)多的轉(zhuǎn)角或無(wú)規(guī)則轉(zhuǎn)曲會(huì)導(dǎo)致蛋白質(zhì)結(jié)構(gòu)不緊湊,從而降低其穩(wěn)定性[24].因此,這3種氨基酸的嗜壓性能最低.
2.2 氨基酸壓力不對(duì)稱(chēng)指數(shù)與其理化性質(zhì)相關(guān)性分析
將計(jì)算所得PAI值與氨基酸的70種理化性質(zhì)分別進(jìn)行相關(guān)性分析,得到了相關(guān)系數(shù)較大(|r|>0.49,p<0.05)的13個(gè)理化性質(zhì),如表4所示.表4中的13個(gè)理化性質(zhì),包括氨基酸固有的理化性質(zhì)參數(shù)(分子體積、折射率等)、熱力學(xué)參數(shù)(熱容、水解自由能)、結(jié)構(gòu)參數(shù)(螺旋作用面積、螺旋-無(wú)規(guī)則卷曲平衡常數(shù)、無(wú)規(guī)則轉(zhuǎn)曲形成趨勢(shì))、親疏水性參數(shù)(疏水性轉(zhuǎn)移指數(shù)、疏水性環(huán)繞指數(shù))和可及表面積(accessible surface area,ASA)等.在這13個(gè)理化性質(zhì)中,只有氨基酸的無(wú)規(guī)則卷曲傾向性(coil tenden-cies,CT)與PAI值呈負(fù)相關(guān),其他的均為正相關(guān).其中,氨基酸分子的大小與其PAI值呈正相關(guān)(r=0.531,p=0.013).這說(shuō)明氨基酸分子體積越大,其嗜壓特性越強(qiáng),這與之前的報(bào)道相反[8].

表4 與PAI相關(guān)系數(shù)最大的13個(gè)理化性質(zhì)Tab.4 Significant physicochemical properties correlated with PAI
雖然體積小的氨基酸殘基更容易占據(jù)蛋白分子內(nèi)外空間,有助于蛋白質(zhì)折疊[8],但分子體積更大的氨基酸更容易讓蛋白分子中的空穴的數(shù)量和體積減少,而這對(duì)蛋白質(zhì)的穩(wěn)定性具有積極作用[25].氨基酸水合自由能是指其在溶解狀態(tài)下與其周?chē)粚铀肿影l(fā)生相互作用的強(qiáng)弱,它與氨基酸自身的疏水性及與水分子的接觸面積正相關(guān)[26].研究表明:高壓條件下蛋白質(zhì)結(jié)構(gòu)的變化主要是由于其水合作用的變化所引起的,而可及表面積較大的氨基酸(尤其是位于分子表面)更容易和水分子接觸,增加這種水合作用,從而保證其在高壓條件下的穩(wěn)定性[27].這同時(shí)也解釋了氨基酸分子可及性與PAI之間這種明顯的正相關(guān)性(表4和圖1).

圖1 PAI與氨基酸可及表面積的相關(guān)性Fig.1 Correlation of amino acids accessible surface area with PAI
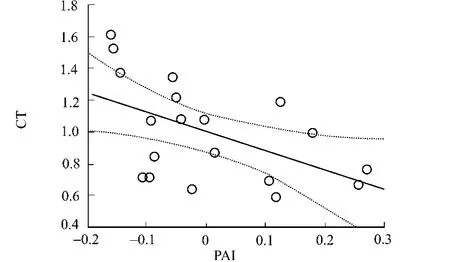
圖2 PAI與氨基酸無(wú)規(guī)則卷曲傾向的相關(guān)性 Fig.2 Correlation of amino acids coil tendencies with PAI
此外,在高壓條件蛋白變性過(guò)程中,當(dāng)水分子進(jìn)入到疏水內(nèi)核區(qū)后會(huì)加劇蛋白質(zhì)變性的過(guò)程.因此,在其分子中需要增加疏水性氨基酸的數(shù)目,以抵制水分子的進(jìn)入,同時(shí)也減少分子中空穴的數(shù)量和體積,避免水分子快速進(jìn)入[28].這說(shuō)明了PAI值和氨基酸疏水性及氨基酸體積大小有較強(qiáng)的正相關(guān)性.
在其他一些極端蛋白(如嗜熱和嗜鹽)中,二級(jí)結(jié)構(gòu)的組成含量及不同二級(jí)結(jié)構(gòu)中氨基酸組成存在明顯差異[29].文獻(xiàn)[9]的研究也表明:嗜壓和非嗜壓蛋白在不同二級(jí)結(jié)構(gòu)中氨基酸組成差異明顯,這種現(xiàn)象可通過(guò)PAI值與氨基酸不同二級(jí)結(jié)構(gòu)傾向因子解釋.經(jīng)檢查,PAI值和α-螺旋傾向因子的相關(guān)系數(shù)為0.34,和β-折疊傾向因子的相關(guān)系數(shù)為0.44,和轉(zhuǎn)角傾向因子的相關(guān)系數(shù)為-0.43,而和無(wú)規(guī)則卷曲傾向因子的相關(guān)系數(shù)為-0.53,4種相關(guān)性均達(dá)有統(tǒng)計(jì)學(xué)意義的水平(p<0.05).可見(jiàn),PAI值與α-螺旋傾向因子相關(guān)性最小,與無(wú)規(guī)則卷曲傾向因子相關(guān)性最強(qiáng)(表4和圖2).PAI值與β-折疊傾向因子呈較明顯的正相關(guān),即表明越容易形成β-折疊的氨基酸其嗜壓性能越強(qiáng),如Trp和Ile是強(qiáng)烈的β-折疊形成子,其嗜壓性能最強(qiáng),因此,在嗜壓蛋白的β-折疊出現(xiàn)的頻率也最高[9].另一方面,PAI值和轉(zhuǎn)角及無(wú)規(guī)則卷曲傾向性呈負(fù)相關(guān),尤其是和無(wú)規(guī)則卷曲.眾所周知,Pro和Gly是強(qiáng)烈的轉(zhuǎn)角及無(wú)規(guī)則卷曲形成子,其非嗜壓性能最強(qiáng),因此,在非嗜壓蛋白的轉(zhuǎn)角或無(wú)規(guī)則卷曲中含量也最高[9],而Ile是強(qiáng)烈的轉(zhuǎn)角及無(wú)規(guī)則卷曲破壞子,故其嗜壓性能很強(qiáng),在嗜壓蛋白的轉(zhuǎn)角或無(wú)規(guī)則卷曲中含量也最高[9].
3 結(jié)論
由于高壓對(duì)生物分子(尤其是蛋白質(zhì)或酶)的獨(dú)特影響使其在不同領(lǐng)域受到了共同的關(guān)注[2].但在真正廣泛應(yīng)用之前,有一個(gè)基本的問(wèn)題擺在面前,那就是高壓條件下蛋白質(zhì)結(jié)構(gòu)與其功能有何關(guān)系,是否能在認(rèn)知其構(gòu)效關(guān)系的基礎(chǔ)上設(shè)計(jì)出耐高壓的酶.由此也可見(jiàn),了解蛋白質(zhì)在高壓條件下穩(wěn)定性的機(jī)理在理論和應(yīng)用上均具有重要價(jià)值.
通過(guò)嚴(yán)格選取目前發(fā)現(xiàn)嗜壓菌最多的Shewanella的2對(duì)微生物,獲取其全蛋白質(zhì)組序列,并從中選取473對(duì)直系同源序列,盡最大可能地減少了其他因素對(duì)最終結(jié)果的影響,計(jì)算出氨基酸壓力不對(duì)稱(chēng)指數(shù).該結(jié)果與之前報(bào)道存在較大差異.究其主要原因,可能是由于之前研究選取的是Pyrococcus屬的2個(gè)微生物,這兩種微生物屬古菌,同時(shí)也是生長(zhǎng)在高溫條件下的是嗜熱菌.本實(shí)驗(yàn)選取的是細(xì)菌,且生長(zhǎng)在低溫條件下,也許是其中的嗜壓蛋白在不同溫度及不同系統(tǒng)發(fā)育階段會(huì)有不同適應(yīng)壓力的機(jī)制,已有研究者注意到這種現(xiàn)象[30].但無(wú)論如何,研究結(jié)果至少可作為對(duì)之前所得氨基酸壓力不對(duì)稱(chēng)指數(shù)的補(bǔ)充及修正,這對(duì)了解占嗜壓菌大多數(shù)的低溫菌中嗜壓蛋白穩(wěn)定性機(jī)制應(yīng)該更具有參考價(jià)值,相關(guān)研究仍需進(jìn)一步深入.
[1]EI-HAJJ Z W,ALLCOCK D,TRYFONA T,et al.Insights into piezophily from genetic studies on the deep-sea bacterium,Photobacterium profundum SS9[J].Ann NY Acad Sci,2010,1189:143-148.
[2]ABRAM A,F(xiàn)ILIP M,MARC E G H,et al.Biotechnology under high pressure:Applications and implications[J].Trends Biotechnol,2009,27(7):434-441.
[3]BUCKOW R,WEISS U,HEINZ V,et al.Stability and catalytic activity ofα-amylase from barley malt at different pressure-temperature conditions[J].Biotechnol Bioeng,2007,97(1):1-11.
[4]MOMBELLI E,SHEHI E,F(xiàn)USI P,et al.Exploring hyperthermophilic proteins under pressure:Theoretical aspects and experimental findings[J].Biochim Biophys Acta,2002,1595(1/2):392-396.
[5]FRANCESCA S,STEFANO C,F(xiàn)EDERICO M L,et al.Piezophilic adaptation:A genomic point of view[J].J Biotechnol,2006,126(1):11-25.
[6]MCDONALD J H.Temperature adaptation at homologous sites in proteins from nine thermophile-mesophile species pairs[J].Genome Biol Evol,2010,2:267-276.
[7]GOODARZI H,TORABI N,NAJAFABADI H S,et al.Amino acid and codon usage profiles:Adaptive changes in the frequency of amino acids and codons[J].Gene,2008,407:30-41.
[8]DI GIULIO M.A comparison of proteins fromPyrococcus furiosus and Pyrococcus abyssi:Barophily in the physicochemical properties of amino acids and in the genetic code[J].Gene,2005,346:1-6.
[9]張光亞,李紅春,高嘉強(qiáng),等.氨基酸和二肽組成對(duì)嗜壓微生物蛋白質(zhì)穩(wěn)定性的影響[J].微生物學(xué)報(bào),2009,49(2):198-203.
[10]張光亞,李紅春,高嘉強(qiáng),等.嗜壓蛋白氨基酸殘基溶劑可及性對(duì)其穩(wěn)定性的影響[J].微生物學(xué)報(bào),2010,50(5):621-627.
[11]FANG J,ZHANG L,BAZYLINSKI D A.Deep-sea piezosphere and piezophiles:Geomicrobiology and biogeochemistry[J].Trends Microbiol,2010,18(9):413-422.
[12]DELONG E F,F(xiàn)RANKS D G,YAYANOS A A.Evolutionary relationships of cultivated psychrophilic and barophilic deep-sea bacteria[J].Appl Enviorn Microbiol,1997,63(5):2105-2108.
[13]AONO E,BABA T,ARA T,et al.Complete genome sequence and comparative analysis of Shewanella violacea,a psychrophilic and piezophilic bacterium from deep sea floor sediments[J].Mol Biosyst,2010,6(7):1216-1226.
[14]JAIN E,BAIROCH A,DUVAUD S,et al.Infrastructure for the life sciences:Design and implementation of the UniProt website[J].BMC Bioinformatics,2009,10:136.
[15]ALTSCHUL S F,MADDEN T L,SCHAFFER A A,et al.Gapped BLAST and PSI-BLAST:A new generation of protein database search programs[J].Nucleic Acids Res,1997,25(17):3389-3402.
[16]LARKIN M A,BLACKSHIELDS G,BROWN N P,et al.Clustal W and Clustal X version 2.0[J].Bioinformatics,2007,23(21):2947-2948.
[17]MCDONALD J H,GRASSO A M,REJTO L K.Patterns of temperature adaptation in proteins from Methanococcus and Bacillus[J].Mol Biol Evol,1999,16(12):1785-1790.
[18]MCDONALD J H.Patterns of temperature adaptation in proteins from the bacteria Deinococcus radiodurans and Thermus thermophilus[J].Mol Biol Evol,2001,18(5):741-749.
[19]KAWASHIMA S,POKAROWSKI P,POKAROWSKA M,et al.AAindex:Amino acid index database,progress report 2008[J].Nucleic Acids Res,2008,36(S1):202-205.
[20]MA J C,DOUGHERTY D A.The cation-πinteraction[J].Chemical Reviews,1997,97(5):1303-1324.
[21]PACK S P,YOO Y J.Protein thermo-stability:Structure-based difference of amino acid between thermophilic and mesophilic proteins[J].J Biotechnol,2004,111(3):269-277.
[22]ARNóRSDóTTIR J,KRISTJáNSSON M M,F(xiàn)ICNER R.Crystal structure of a subtilisin-like serine proteinase from a psychrotrophic Vibrio species reveals structural aspects of cold adaptation[J].FEBS J.2005,272(3):832-845.
[23]CHOU P Y,F(xiàn)ASMAN G D.Empirical predictions of protein conformation[J].Ann Rev Biochem,1978,47:251-276.
[24]SONAN G K,RECEVEUR-BRECHOT V,DUEZET C,et al.The linker region plays a key role in the adaptation to cold of the cellulase from an Antarctic bacterium[J].Biochem J,2007,407(2):293-302.
[25]ANDRáS S,PéTER Z.Structural differences between mesophilic,moderately thermophilic and extremely thermophilic protein subunits:Results of a comprehensive survey[J].Structure,2000,8(5):493-504.
[26]OOI T,OOBATAKE M,NMMETHY G,et al.Accessible surface areas as a measure of the thermodynamic parameters of hydration of peptides[J].Proc Natl Acad Sci USA,1987,84(10):3086-3090.
[27]SILVA J L,F(xiàn)OGUEL D.Hydration,cavities and volume in protein folding,aggregation and amyloid assembly[J].Phys Biol,2009,6(1):15002.
[28]NOLWENNIG R,JEAN R,GéRARD D.Development of high hydrostatic pressure in biosciences:Pressure effect on biological structures and potential applications in biotechnologies[J].Biotechnol Adv,2010,28(6):659-672.
[29]SANDIP P,SUMIT K B,SABYASACHI D,et al.Molecular signature of hypersaline adaptation:Insights from genome and proteome composition of halophilic prokaryotes[J].Genome Biol,2008,9R(4):70.
[30]RIE S,CHIAKI K,AKIHIKO N.Amino acid substitutions in malate dehydrogenases of piezophilic bacteria isolated from intestinal contents of deep-sea fishes retrieved from the abyssal zone[J].J Gen Appl Microbiol,2006,52(1):9-19.
Revision of the Amino Acid Pressure Asymmetry Index of Piezophilic Proteins
ZHANG Guang-ya
(College of Chemical Engineering,Huaqiao University,Xiamen 361021,China)
A comparison was made between 473orthologous proteins fromShewanella.The pattern of asymmetries in the 21 662amino acid mutation sites reveals some common trends.The pressure asymmetry index(PAI)identifies the amino acids tryptophan(Trp)and isoleucine(Ile)as those having the most piezophilic behavior,and proline(Pro),glycine(Gly)and asparagines(Asn)as the least piezophilic,which is totally different from the former result.The PAI makes it possible to visualize the amino acid properties that best explain piezophily.Among the 70properties of amino acids,there are 13significantly correlated with PAI value.Only the coil tendencies are negatively correlated with PAI value,that is to say,on average,higher PAI values are associated with fewer tendencies to form coil.The PAI we established here may be more suitable to the piezophiles living in the cold environment,and a revision of the previous PAI values was made.
piezophilic microbes;pressure asymmetry index;amino acid;coil tendencies;appropriate pressure mechanism
黃曉楠 英文審校:劉源崗)
Q 936
A
1000-5013(2012)01-0044-07
2011-06-11
張光亞(1975-),男,副教授,主要從事酶與生物信息學(xué)的研究.E-mail:zhgyghh@hqu.edu.cn.
福建省自然科學(xué)基金資助項(xiàng)目(2007J0360);華僑大學(xué)高層次人才科研啟動(dòng)項(xiàng)目(10BS220)
