植物數量性狀“主基因+多基因”混合遺傳模型及其在煙草上的應用
2013-03-21 01:29:50張興偉王志德劉艷華任民楊洋
中國煙草學報 2013年3期
張興偉,王志德,劉艷華,任民,楊洋
1 農業部煙草生物學與加工重點實驗室,中國農業科學院煙草研究所,青島 266101;
2 丹東市煙草專賣局(公司)寬甸分公司,丹東 118222
煙草育種所注重的若干性狀,一般都是數量性狀(quantitative character)。數量性狀由多個基因控制,每個基因控制性狀表現的一部分,難以根據表型的差異檢測出具體基因。經典數量遺傳學一般只能從一組基因的總體上得到有關遺傳效應及其相對重要的信息。而對植物遺傳改良來說,需要有關數量性狀遺傳體系以及其中具體基因的重要信息,以便通過育種手段進行遺傳操作。植物數量性狀“主基因+多基因”混合遺傳模型的出現,能有效地解決上述問題。“主基因+多基因”混合遺傳模型是植物數量性狀的通用模型,蓋鈞鎰等[1]在此基礎上發展了一套適合植物遺傳分析的分離分析方法。該方法適于育種工作者利用雜種分離世代的數據對育種性狀的遺傳組成做出判斷,制訂相應的育種策略,也可用以校驗QTL定位所揭示的性狀遺傳組成。
1 植物數量性狀“主基因+多基因”混合遺傳模型的主要環節
1.1 分離世代的理論分布
在二倍體核遺傳、不存在母體效應、主基因和多基因無互作和連鎖、配子和合子均無選擇的假定下,每一主基因型由于多基因和環境的修飾呈正態分布,整個分離世代的表型分布為多個主基因型正態成分分布的混合分布;不分離世代如P1、P2、F1等的表型分布為正態分布。
1.2 遺傳模型
目前,植物數量性狀“主基因+多基因”混合遺傳模型主要包括以下幾種模型:一對主基因(A)、兩對主基因(B)、多基因(C)、一對主基因+多基因(D)、兩對主基因+多基因(E)、三對主基因(F)、三對主基因+多基因(G)。
1.3 試驗群體類型
單世代分析,包括P1、P2、DH或RIL群體3個世代的聯合分析,P1、P2、F1、F24個世代的聯合分析,P1、P2、F1、F2、F2:35 個世代的聯合分析,P1、P2、F1、F2、B1、B26 個世代聯合分析以及 P1、P2、F1、F2:3、B1:2、B2:26個家系世代試驗聯合分析。
1.4 ECM算法及對數極大似然函數
按照試驗群體類型建立各種模型的混合分布函數,并建立各模型的對數極大似然函數。由各試驗群體及世代的觀察次數分布通過IECM(iterated expectation and conditional maximization)[2]算法對各模型的對數極大似然函數估計出相應的各世代、各成分分布的參數,以及相應的似然函數值。
1.5 篩選最優模型及估算遺傳參數
方法是由模型的似然函數值計算出AIC(Akaike’s information criterion)[3]值,AIC值最小的模型為最佳模型,模型間AIC值差異不大時要進行適合性測驗,這組測驗包括均勻性U12、U22及檢驗,nW2檢驗和Dn檢驗,同時選得各成分分布參數。采用最小二乘法由最優遺傳模型的各成分分布參數計算出主基因和多基因的相關遺傳參數。
2 植物數量性狀“主基因+多基因”混合遺傳模型在煙草中的應用(詳見表1)
2.1 在植物學性狀上的應用
王日新等[4]對株高研究結果表明,其遺傳符合2對加性-顯性-上位性主基因+加性-顯性多基因混合遺傳模型(E1),主基因遺傳率為71.60%-88.55%。張興偉等[5]對株高研究結果表明,其遺傳符合2對加性-顯性-上位性主基因+加行-顯性-上位性多基因混合遺傳模型(E0),主基因遺傳率為57.53%。張興偉等[6]對葉數、葉面積的研究結果表明,其遺傳也符合E0模型,主基因遺傳率分別為42.63%和30.32%。張興偉等[5]同時對其它幾個植物學性狀的研究表明,鮮葉重遺傳也符合E0模型,主基因遺傳率為44.26%;移栽至中心花開放天數遺傳符合E1模型,主基因遺傳率為64.79%;莖圍和比葉重均受1對完全顯性主基因+加性-顯性多基因(D3)控制,遺傳以多基因為主,其多基因加性效應和顯性效應大小相當,比葉重遺傳主基因、多基因的加性效應和顯性效應大致相當,主基因遺傳率分別為2.48%和38.71%。葉形指數受1對加性-顯性主基因+加性-顯性-上位性多基因(D0)控制,主基因加性效應與顯性效應基本相當,主基因遺傳率為49.64%。葉長、葉寬、節距和蒴果重受加性-顯性-上位性多基因(C0)控制,多基因遺傳率分別為60.75%、62.14%、75.08%和82.34%。
2.2 在抗病上的應用
蔡長春等[7]對黑脛病的研究結果表明,黑脛病發病率是由2對具有互補作用的主基因+多基因(E-1-7)控制,主基因遺傳率為54.14%。高加明等[8]對青枯病的研究結果表明,其遺傳符合E1模型,基因效應以上位性效應為主,主基因遺傳率為49.63%。牟建英等[9]對白粉病的研究結果表明,其遺傳符合E0模型,以顯性效應及上位性效應為主,主基因遺傳率為84.43%。
2.3 在化學成分上的應用
蔡長春等[10]對斬株前和晾制后白肋煙的煙堿含量研究結果表明,其遺傳受2對主基因控制,同時存在多基因的修飾作用,前者為累加作用(E-1-6),后者為互補作用(E-1-7),主基因遺傳率分別為54.88%和45.63%。尹天水等[11]對鉀含量的研究結果表明,其遺傳符合D0模型,主基因遺傳率為52.78%。
2.4 在生理性狀上的應用
張興偉等[12]對葉綠素含量研究結果表明,其遺傳符合1對加性-顯性主基因+加性-顯性-上位性多基因混合遺傳模型(D0),主基因遺傳率為48.94%。張興偉等[13]對光合性狀研究結果表明,光合速率和蒸騰速率的遺傳符合C0模型,多基因遺傳率分別為20.69%和13.56%,氣孔導度遺傳符合E0模型,以上位性效應為主,主基因遺傳率為40.59%。
2.5 在烘烤性狀上的應用
倪超等[14]、王英俊等[15]對易烤性研究結果表明,其遺傳符合2對加性-顯性-上位性主基因+加性-顯性多基因混合遺傳模型(E1),主基因遺傳率分別為64.09%、65.61%。郝賢偉等[16]對耐烤性研究結果表明,其遺傳符合E0模型,以加性效應為主,主基因遺傳率為56.81%-79.61%。
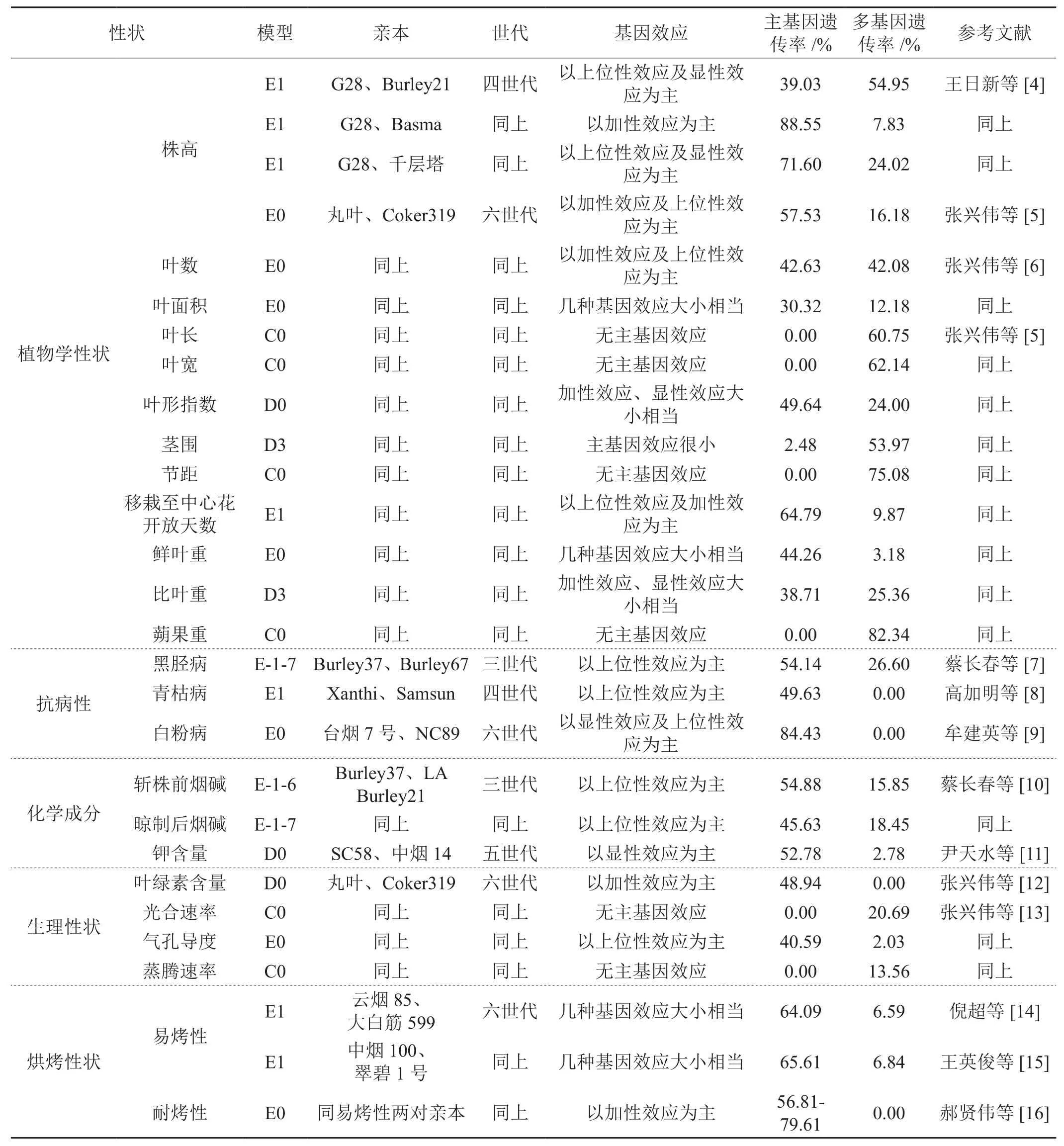
表1 近7年來對栽培煙草重要性狀的遺傳分析
植物數量性狀“主基因+多基因”混合遺傳模型在煙草上應用以來,在煙草植物學性狀、抗病性狀、化學成分、生理性狀及烘烤性狀等方面,對于揭示相關性狀的遺傳組成及遺傳效應發揮了積極的效用,該方法適于育種工作者利用雜種分離世代的數據對育種性狀的遺傳組成做出初步判斷,制訂相應的育種策略,也可用以校驗QTL定位所揭示的性狀遺傳組成。同時該模型也需要不斷拓展,以滿足復雜數量性狀分析的客觀要求。
3 關于“主基因+多基因”混合遺傳模型的幾點思考
“主基因+多基因”混合遺傳模型是植物數量性狀的通用模型,蓋鈞鎰等[1]在此基礎上發展了一套適合植物遺傳分析的1對主基因+多基因、2對主基因+多基因和3對主基因+多基因混合模型的分離分析方法。此方法自提出以來已廣泛應用于水稻[17]、棉花[17]、油菜[19]、小麥[20]、大豆[21]等多種糧食和經濟作物,而在煙草上的應用研究[4-16],這幾年也逐漸多了起來。在實際應用中,有些研究者拿來數據直接套用模型,存在不少誤區,下面是筆者對于“主基因+多基因”混合遺傳模型的幾點思考。
3.1 極端親本材料的選擇
親本一般要選擇關于某性狀的極端材料(如高抗和高感),這樣有助于消除中間材料的干擾和誤差,有利于研究該性狀真實的遺傳規律。
3.2 試驗設計
一般要進行正反交和隨機區組設計,三次重復。正反交結果可確定是細胞核效應還是質核互作效應;隨機區組設計重復之間和處理之間差異是否顯著,可說明2個親本某性狀在遺傳方面是否存在著真實穩定的差異,進而決定是否可以作為研究該性狀遺傳規律的試材進行分析。
3.3 數據分析
獲取數據后,首先要看不分離群體(P1、P2和F1)的數據是否符合正態分布,SAS可做正態分布分析,最簡單的方法畫個柱形圖看看;若不符合,剔除明顯變異的數據,再進行下一步分析;根據AIC準則選擇模型時,一般選擇3-5個作為備選模型,具體選擇多少,要視具體情況而定,若已選模型的AIC值與其它相比,差異較大,則少選幾個,反之,則多選幾個。
3.4 多年多點多時期數據的精準測定
對某一性狀最好進行多年多點多時期的精準測定,這樣可研究某一性狀數量基因的時空表達規律。
3.5 “主基因+多基因”混合遺傳模型基因對數的拓展
育種實踐和QTL定位的結果表明,很多數量性狀的遺傳受到4對甚至更多的主基因控制。章元明等[22]構建的遺傳模型最多只包含3對主基因,已不能滿足育種研究的需要,因此需要拓展遺傳模型,目前王金社等[23]已經把重組自交系群體初步拓展到4對主基因,但更多遺傳群體的基因對數仍需拓展。
3.6 “主基因+多基因”混合遺傳模型在煙草上的優先應用策略
近幾年,隨著煙草基因組計劃研究的深入,煙草諸多性狀的QTL定位成了研究熱點。筆者建議,在進行QTL分析前,可采用“主基因+多基因”混合遺傳模型對相應性狀進行優先分析。這樣既可以在早期對育種性狀的遺傳組成做出初步判斷,制訂相應的育種策略,也可用以校驗QTL定位所揭示的性狀遺傳組成。
[1] 蓋鈞鎰,章元明,王建康.植物數量性狀遺傳體系[M].北京:科學出版社,2003.
[2] 章元明,蓋鈞鎰. 數量性狀分離分析中分布參數估計的IECM算法[J]. 作物學報,2000,26(6):699-706.
[3] Akaike H. On entropy maximum principle [M]// Krishnaiah P R. Applications of Statistics. Amsterdam: NorthHolland Publishing Res, 1977:27-41.
[4] 王日新,任民,張興偉,等. 普通煙草栽培種內株高性狀主基因加多基因遺傳分析[J]. 中國煙草科學,2009,30(2):15-20.
[5] 張興偉,王志德,任民,等. 烤煙幾個植物學性狀的遺傳分析[J]. 中國煙草科學,2012,33(5):1-8.
[6] 張興偉,王志德,孫玉合,等. 烤煙葉數、葉面積的遺傳分析[J]. 植物遺傳資源學報,2012,13(3):467-472.
[7] 蔡長春,張俊杰,黃文昌,等. 利用DH群體分析白肋煙黑脛病抗性的遺傳規律[J]. 煙草科技,2009(1):54-59,63.
[8] 高加明,王志德,張興偉,等. 香料煙青枯病抗性基因的遺傳分析[J]. 中國煙草科學,2010,31(1):1-4.
[9] 牟建英,錢玉梅,張興偉,等. 煙草白粉病抗性基因的遺傳分析[J]. 植物遺傳資源學報,2013,14(4).
[10] 蔡長春,張俊杰,黃文昌,等. 利用DH群體分析白肋煙煙堿含量的遺傳規律[J]. 中國煙草學報,2009,15(4):55-60.
[11] 尹天水,王樹會,石磊. 烤煙煙葉鉀含量的遺傳分析[J].煙草科技,2005(5):34-38.
[12] 張興偉,王志德,牟建民,等. 烤煙葉綠素含量遺傳分析[J].中國煙草學報,2011,17(3):48-52.
[13] 張興偉. 煙草微核心種質構建及相關性狀數量遺傳分析[D]. 中國農業科學院研究生院博士學位論文,2013,10-11.
[14] 倪超,徐秀紅,張興偉,等. 烤煙品種易烤性相關性狀的主基因+多基因遺傳分析[J]. 中國煙草科學,2011,32(1):1-4, 11.
[15] 王英俊,倪超,翟明泉,等. 烤煙易烤性遺傳分析[J]. 作物研究,2011,25(1):42-46.
[16] 郝賢偉,徐秀紅,許家來,等. 烤煙品種耐烤性遺傳分析[J].中國農業科學,2012,45(23):4939-4946.
[17] 朱世楊,郭媛,洪德林.水稻種子抗老化遺傳分析[J].遺傳,2008,30(2):217-224.
[18] 袁有祿,張天真,郭旺珍,等. 棉花高品質纖維性狀的主基因與多基因遺傳分析[J]. 遺傳學報,2002,29(9):827-834.
[19] 張書芬,傅廷棟,朱家成,等. 甘藍型油菜芥酸含量的基因分析[J]. 中國農業科學,2008,41(10):3343-3349.
[20] 張立平,趙昌平,單福華,等. 小麥光溫敏雄性不育系BS210育性的主基因+多基因混合遺傳分析[J].作物學報,2007,33(9):1553-1557.
[21] 王春娥,蓋鈞鎰,傅三雄,等. 大豆豆腐和豆乳得率的遺傳分析與QTL定位[J]. 中國農業科學,2008,41(5):1274-1282.
[22] 章元明,蓋鈞鎰,王永軍. 利用P_1、P_2和DH或RIL群體聯合分離分析的拓展[J]. 遺傳,2001,23(5):467-470.
[23] 王金社,李海旺,趙團結,等. 重組自交家系群體4對主基因加多基因混合遺傳模型分離分析方法的建立[J].作物學報,2010,36(2):191-201.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
電子制作(2018年18期)2018-11-14 01:48:24
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
山東工業技術(2016年15期)2016-12-01 05:31:22
光學精密工程(2016年6期)2016-11-07 09:07:19
當代化工研究(2016年9期)2016-03-20 16:22:08
