中國恒河猴腸道菌群特征揭示
2013-09-13 11:47:06趙娜劉社蘭路紀琪何宏軒趙寶華
生物技術通報 2013年7期
趙娜 劉社蘭 路紀琪 何宏軒 趙寶華
人類和靈長類動物在形態、生理、基因和行為上的近似使得靈長類成為研究人類疾病的模型,其中恒河猴(Rhesus Macaques)是被研究得最清楚的舊世界猴,被廣泛應用于生命科學研究領域。在恒河猴養殖業,尤其是處于食性轉換期和特殊生理時期的恒河猴個體中,由于病毒和腸道病原體傳染造成的疾病嚴重威脅恒河猴的健康。傳統研究腸道菌群特點的方法主要是選擇性培養,但是由于腸道中大多數細菌不能用傳統研究方法進行培養、分離和鑒定[1,2],因此人們對恒河猴腸道及糞樣微生物的變化規律知之甚少。
DGGE技術可以快速檢測傳統條件不能培養的細菌,發現新的微生物,作為微生物分子生態學研究的重要手段,被廣泛地應用于微生物群落多樣性和動態分析[3]。16S rRNA基因DGGE圖譜條帶數目可以反映微生物菌群的多樣性,條帶染色后的亮度可以相應反映細菌在群體中的豐度。對PCR-DGGE分離得到的條帶進行克隆測序或雜交可以進一步準確鑒定細菌的種屬[4,5]。與普通PCR相比,實時熒光檢測技術(Real-time PCR)可以對整個PCR過程中產物的擴增過程加以監測,能夠直觀地反映PCR模板濃度與擴增產物含量之間的關系,具有高度準確性,目前,這項技術已經被廣泛用于動物腸道菌群檢測及環境微生態的檢測研究[6]。
McKenna等[7]運用焦磷酸測序技術研究了慢病毒感染及慢性結腸炎的恒河猴的腸道微生態特點,并簡單與健康恒河猴進行比較,但是迄今為止沒有系統關于不同年齡段健康恒河猴個體腸道主要菌群多樣性及其特點的報道。本研究采用PCR-DGGE和實時熒光定量PCR(Real-time quantitative PCR)技術研究24只處于6個不同年齡組、不同生理狀態的恒河猴個體糞樣中腸道菌群的變化規律,從定性和定量的角度加以揭示,以期發現由于食性改變及生理狀態改變造成的腸道疾病的根源。
1 材料與方法
1.1 糞樣的收集及處理
選擇飼養在河南省鄭州動物園的24只健康恒河猴,嬰幼齡(1月齡)、青年(1-4歲)、亞成年(4-5歲)、成年(5-10歲)、老年(10歲以上)、孕期(懷孕后期,孕齡15-18周)各4只,除了孕期組以外的五組中個體均雌雄各半。除嬰幼齡個體母乳喂養外,其他組別個體均以胡蘿卜、蘋果、馬鈴薯和玉米餅為飼料,每日投喂兩次。2011年2月,每個個體取3份樣品,每次取樣間隔1周。
所有72個糞便樣品在采樣時均收集在無菌塑料容器內,放置在干冰上,30min內運到實驗室。每份糞便樣品混合均勻后,分裝出3份至滅菌離心管中,200mg每份,然后凍存于-80℃直至分析。
1.2 方法
1.2.1 DNA提取 每個個體凍存的3份不同時間點收集的樣品各一份混合均勻,等量分成3份(200mg每份)。參照Yanfei等[8]方法將樣品加到提前加好300mg0.1mm玻璃珠(Sigma)的2mL破壁管中,加入1.4mL ASL緩沖液(QIAamp DNA Stool Mini Kit)之前,放于冰上。然后于FastPrep FP120震蕩儀上6.5速度震蕩45s,隨后于97℃水浴鍋中5min水浴。隨后參照QIAamp DNA Stool Mini Kit 說明書步驟進行。最后,用200μL TE緩沖液(10mmol/L Tris-HCl,1mmol/L EDTA,pH8.0)洗脫。取 1μL 于NanoDrop ND-1000測所提細菌基因組DNA濃度,其A260/A280在1.6-1.8之間,濃度范圍在50-70ng/μL之間。后將DNA樣品-80℃超低溫冰箱保存備用。
1.2.2 PCR-變性梯度凝膠電泳(PCR-DGGE)
1.2.2.1 PCR擴增 為避免非特異性PCR產物出現,16S rRNA V3可變區基因PCR擴增采用Muyzer等[3]提出的熱啟動touchdown方法。由通用引物(F357-GC clamp:5'-CGCCCGCCGCGCGCGGCGGGCGGGGCGG GGGCACGGGGGGCCTACGGAGGCAGCAG-3',R518:5'-ATTACCGCGGCTGCTGG-3')擴增。
25μL反應體系包括:1 U TaKaRa Taq熱啟動聚合酶,10×PCR buffer,上下游引物各10 pmol,20mmol/L dNTPs,20ng DNA模板。94℃預變性3min,94℃變性1min,65℃退火1min,72℃延伸 1min,20個循環,94℃變性1min,55℃退火1min,72℃延伸 1min,5個循環,72℃延伸 5min,4℃ 15min。為消除產物中的異二聚體,采用reconditioning PCR技術,以20ng的PCR產物為模板進行reconditioning PCR[9]。反應程序如下 :94℃預變性 3min,94℃變性20s,56退火30s,72℃延伸30s,5個循環;72℃延伸7min,4℃ 15min。1%瓊脂糖凝膠檢測擴增結果,UVI凝膠成像系統照相記錄,目的條帶200bp,條帶明亮單一,保存產物以進行DGGE。
1.2.2.2 DGGE 考慮同張DGGE指紋圖譜比較清晰,且樣品前期處理已經做到混合,每組選擇兩個樣品進行DGGE電泳,除孕期恒河猴,其余各組均是雌性個體和雄性個體各一。參照Muyzer的方法,對PCR產物進行DGGE電泳分離。電泳前,對PCR產物進行濃縮使所有樣品達到濃度一致。使用Bio-Rad 公司的D-code Universal Mutation Detection System進行DGGE電泳。配置37%-78%(100%變性梯度包含40%去離子甲酰胺和7mol/L尿素)變性范圍的8%聚丙烯酰胺變性膠,16S rRNA基因 V3 區PCR 產物 15μL+5μL 10×loading buffer 注射針上樣,恒溫60℃,69 V電壓1×TAE緩沖液中電泳16h。
電泳完畢后,使用現稀釋的0.1%SYBER Green I溶液避光染色2次,每次30min。染色結束后用去離子水沖洗膠2次,于UVI自動凝膠成像儀中拍照。對DGGE凝膠上感興趣的條帶,進行割膠回收。
1.2.3 PCR產物擴增及克隆測序
1.2.3.1 PCR擴增及產物純化 DGGE割膠條帶放入50μL RNA free H2O的離心管中,4℃浸泡過夜。取2μL浸泡液作為模板,無GC夾子引物F357和R518各10 pmol進行PCR擴增。程序:94℃預變性3min,94℃變性 20s,56℃退火 30s,72℃延伸 30s,25個循環;72℃延伸7min,4℃ 15min。PCR產物用1%瓊脂糖凝膠電泳拍照確定陽性條帶后,用天根公司DNA回收試劑盒回收。
1.2.3.2 克隆測序 回收產物連接Promega公司pGEM-T Easy Vector并轉化 Escherichia coli DH5α cells。為確保正確插入,陽性克隆子再度用pGEM-T 特異性引物 T7(5'-TAA TAC GAC TCA CTA TAG GG-3')和 Sp6(5'-ATT TAG GTG ACA CTA TAG-3')進行驗證。正確插入的克隆子重新重復之前DGGE的步驟進行DGGE電泳,確定電泳條帶的正確性。電泳位置與之前一致的條帶DNA送北京華大基因測序,每個條帶的平板測4個陽性克隆。
1.2.3.3 DNA序列進化樹構建 BLAST軟件分析序列去除載體序列并對序列加以整理對齊。將整理后的序列信息導入MOTHUR軟件,按照97%的相似性標準,將這些序列劃分分類操作單元(OTU)。將這些OTU序列與NCBI GenBank的16S rRNA基因序列進行比對,下載相似性較高的序列,BLAST處理后作為背景序列與測序所得序列一起導入MEGA5采用鄰接算法進行構建進化樹[10]。
1.2.4 DGGE分析 采用Quantity? One 1-D Analysis software對DGGE凝膠圖譜進行標化處理,根據圖譜中各條帶數目、遷移位置和強度進行膠的數字化處理,應用UPGMA算法進行泳道相似性聚類分析,將數據導入MATLAB 2008進行主成分分析(principal component analysis,PCoA)。主成分分析是將一組變量上分散的信息進行集中,主要對某幾個綜合性指標(主成分)進行探索性統計的分析方法。對DGGE圖譜進行PCoA分析可以彌補相似性聚類分析受背景及其他無關因素影響的缺點,可以得到樣品間是否存在差異,PCoA載荷量分析可以進而得到造成這種差異的原因[11,12]。對PCR-DGGE分離得到的條帶進行克隆測序或雜交可以進一步準確鑒定細菌的種屬。
1.2.5 腸道優勢菌屬實時熒光定量PCR檢測 采用美國 Bio-Rad 公司 DNA Engine Opticon? 2 apparatus實時熒光定量PCR儀對腸道菌群進行定量,菌屬特異性引物,見表 1。20μL 擴增體系由 10μL SYBR Premix Ex TaqTM(TaKaRa),300nmol/L引物以及 1μL 模板 DNA組成。每個反應都重復3次。熒光定量PCR反應條件為:95℃ 預變性5min,95℃變性30s,退火30s,72℃延伸40s,執行40個循環;每個循環結束后維持81℃ 10s,以避免引物二聚體、二級結構,收集熒光信號。最后72℃延伸5min。每組引物的退火溫度,見表1。每個熒光定量反應均以濃度101-108copies/μL的標準菌種16S rRNA基因片段的質粒DNA構建標準曲線[13]。
1.2.6 統計學分析 使用SPSS17.0對實時熒光PCR數據進行Fisher’s Exact Test 和one-way ANOVA進行統計學分析,每組數據分別以各自的“平均數±標準差”表示。樣本基因的拷貝數以每克濕重樣品中細菌數的對數值表示。
2 結果
2.1 腸道微生物群落組間和組內差異的DGGE圖譜
圖1-A是不同組別16S rRNA基因V3可變區PCR-DGGE圖譜,條帶反映了糞樣中的優勢菌群,其數量和位置的差異顯示了各個組別和各個體之間菌群的多樣性。對DGGE圖譜進行標準化分析,將DGGE指紋圖譜按遷移位置標號共有50條電泳條帶。DGGE圖譜經泳道相似性分析顯示屬于明顯的2組(圖1-A),其中嬰幼齡組和孕期組、成年組可歸為一組,其他3組歸為一組,帶譜間相似性均大于60%。嬰幼齡組和孕期組相似性最高,大于67%。

表1 實時熒光定量PCR引物
對DGGE圖譜進行數字化矩陣主成分分析,能夠更直觀地看到不同組別之間腸道菌群的差異。PCoA圖上兩點間距離越近表明主要菌群結構組成上的相似性越高,反之,則相似性越低。圖1-B顯示在主成分27.8%(X軸方向)和21.6%(Y軸方向)時,嬰幼齡組和孕期組有明顯的聚類。這些結果表明,處于哺乳期的嬰幼齡個體與懷孕期個體具有相似的腸道優勢菌群。
圖1-C主成分載荷值分析得到導致主成分分析聚類的主要3條條帶(圖1-A中的A、B、C3條條帶),對應DGGE圖譜進行割膠、克隆、測序鑒定,比對結果顯示它們分別是Ruminococcus bromii(98%,GenBank登錄號DQ882649.1),Bifidobacterium dentium(98%,GenBank登錄號HQ697642.1)和Lactobacillushelveticus(99%,GenBank登 錄 號 HQ61664-3.1),百分數顯示與GenBank比對后的序列相似性。
對DGGE圖譜中50個電泳條帶進行割膠,由于不能排除不同的堿基序列遷移到相同位置的可能,本研究共割膠62條,隨后進行克隆,測序后用MOTHUR軟件分析所有62個條帶屬于43個OTU。與GenBank序列進行比對后構建進化樹顯示所有62個條帶、43個OTUs屬于4個門:厚壁菌門(33/43 OTUs,47/62 DNA 序列),放線菌門(4/43 OTUs,8/62 DNA序列),擬桿菌門(5/43 OTUs,6/62 DNA序列)以及螺旋體門(1/43 OTUs,1/62 DNA序列),絕大多數條帶都屬于厚壁菌門(圖2)。
2.2 不同年齡組恒河猴腸道優勢菌種的定量結果
實時熒光定量結果(表2)顯示在所有恒河猴腸道優勢菌中,梭菌屬和雙歧桿菌最為豐富,將近109copies/g。腸球菌屬最少,不到104copies/g。統計結果顯示梭菌屬clusters ⅪⅤ,腸球菌屬和擬桿菌-普氏菌屬組間差異明顯(P<0.01)。另外梭菌屬clusters Ⅰ,clusters Ⅺ和乳酸桿菌也有較顯著差異(P<0.05)。
a:梭狀芽孢桿菌分組參考文獻[14];b:乳酸桿菌包括乳酸桿菌屬、片球菌屬、明串珠菌屬和魏斯氏菌屬[15];c:梭菌屬clustersⅠ、梭菌屬clusters Ⅺ、梭菌屬clusters ⅪⅤ和柔嫩梭菌屬均屬于梭狀芽孢桿菌
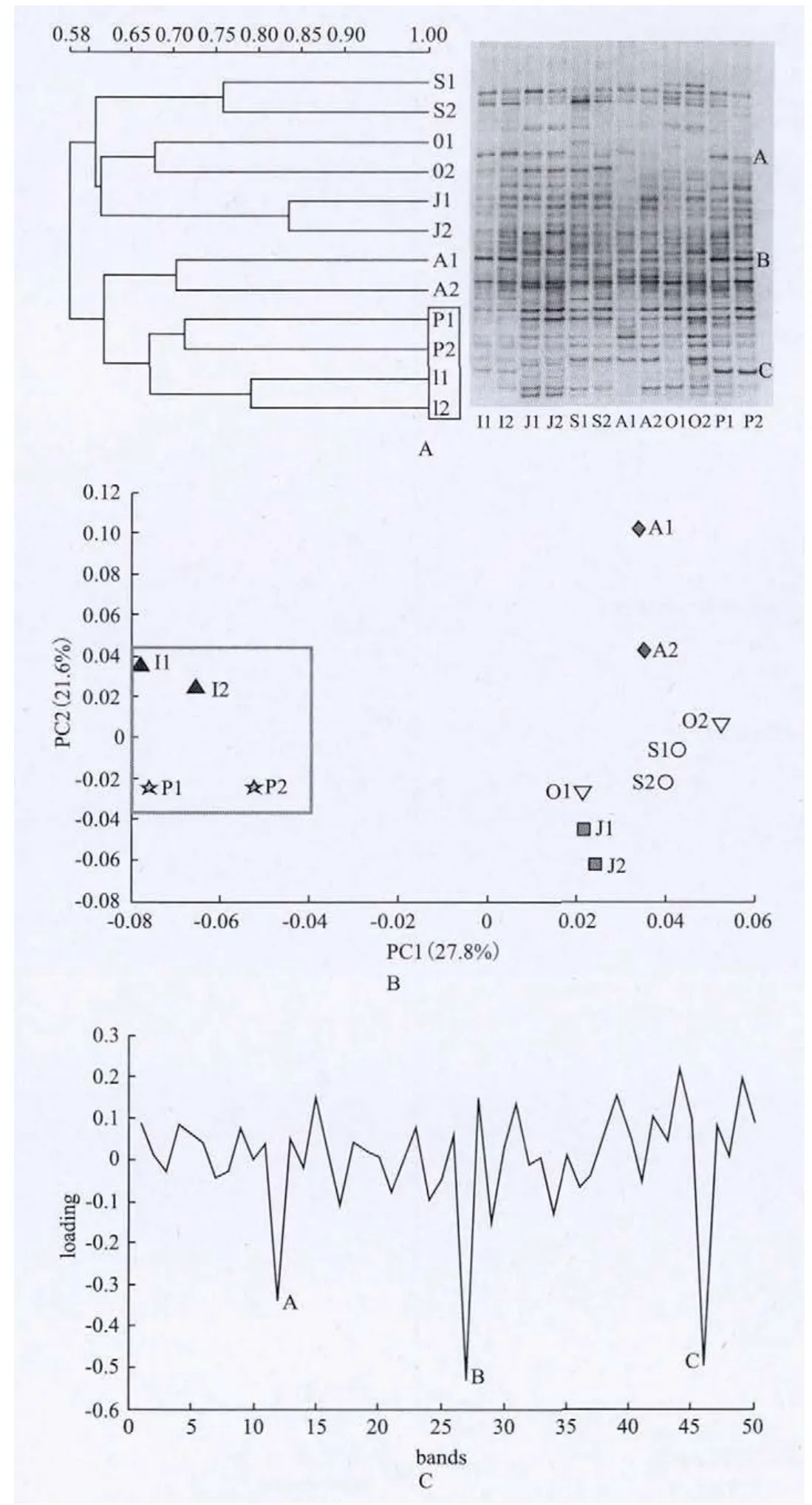
圖1 16S rRNA基因V3區DGGE指紋圖譜
組間統計分析顯示同種細菌在不同年齡恒河猴腸道中的定值存在差異。其中青年恒河猴腸道中擬桿菌-普氏菌屬基因拷貝數,與嬰幼齡恒河猴相比有明顯增多(P=0.049),梭菌屬clusters Ⅺ則減少明顯(P=0.021)。統計數據顯示老年組恒河猴擬桿菌-普氏菌屬細菌拷貝數,與成年組相比顯著增多(P=0.000)。而孕期組恒河猴腸桿菌拷貝數明顯高于同齡未懷孕亞成年和成年恒河猴(P=0.007和P=0.012);梭菌屬clusters Ⅺ拷貝數則明顯低于同齡未懷孕亞成年和成年恒河猴(P=0.017和P=0.023)。圖3顯示,作為顯示腸道健康水平標準的雙歧桿菌和腸桿菌比值(B/E值)與同齡亞成年和成年組恒河猴相比,在孕期恒河猴組有明顯的降低(P=0.002和 P=0.009)。
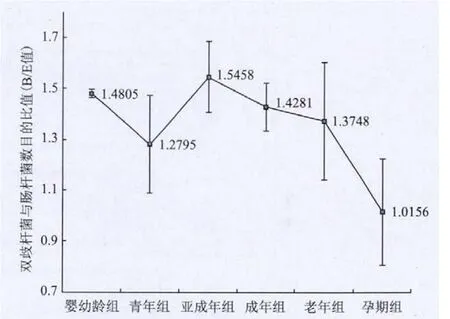
圖3 不同年齡組雙歧桿菌和腸桿菌比值(B/E值)的變化趨勢
另外,q-PCR結果(表2)顯示腸道梭狀芽孢桿菌數量占有絕對優勢,這與PCR-DGGE割膠條帶測序結果(厚壁菌門,33/43 OTUs,47/62 DNA序列)一致。
3 討論
PCR-DGGE方法最大優點是不依賴于培養過程,根據不同變性范圍DNA片段的溶解性而使之分離,因此,近年來分子微生態學研究采用PCR-DGGE技術用以區分各微生境中的優勢菌群,反映其菌群結構多樣性。實時熒光定量PCR不僅能夠監測整個PCR過程中產物的擴增情況,具有較高準確度,又因為該技術能夠在很低模板濃度下進行準確定量,因而被廣泛用于人類及其他動物腸道菌群方面的研究中。本研究首次采用這兩項技術研究不同年齡和不同生理狀態的中國恒河猴糞樣菌群的多樣性。研究結果表明,PCR-DGGE和實時熒光定量PCR技術在研究恒河猴腸道菌群多樣性上非常有效。
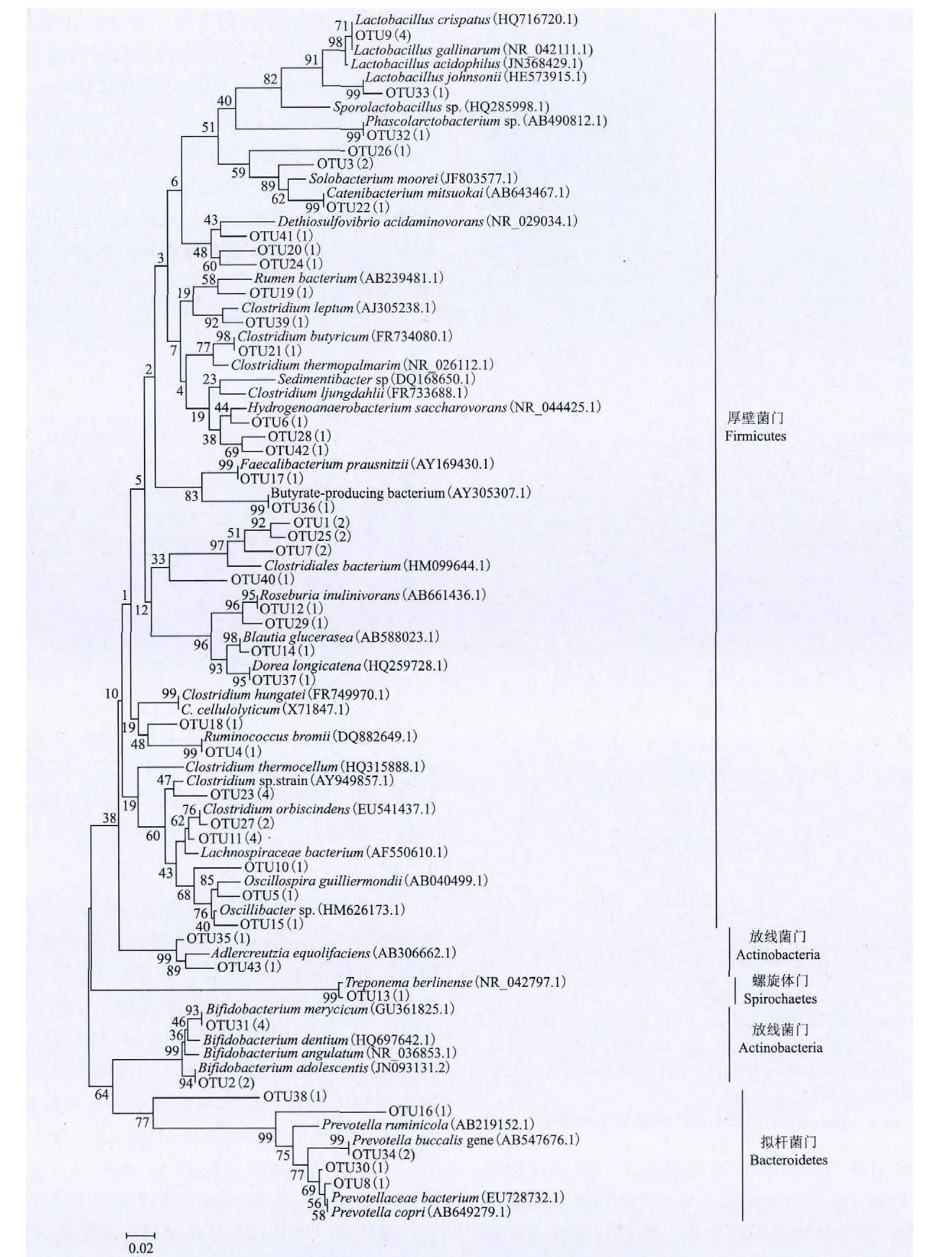
圖2 16S rRNA基因V3區進化樹構建
本研究采集24只不同年齡、不同生理狀況的恒河猴糞樣,分析其腸道菌群的變化。研究結果顯示,母乳喂養階段的嬰幼齡組恒河猴腸道DGGE圖譜與孕期組恒河猴相似度高于67%,主成分分析顯示二者出現明顯聚類。主成分分析載荷值顯示代謝相關功能菌Ruminococcus bromii,健康密切相關益生菌Bifidobacterium dentium和Lactobacillushelveticu為造成聚類的主要差異條帶。有報道稱嬰幼兒腸道菌群處于紊亂狀態[16,17],Jeremy 等[18]報道嬰兒體內腸道微生態處于紊亂的狀態可能跟日常飲食和身體狀況有關,但是機制尚不明確。而在孕期個體中,性激素可以通過膽汁分泌而分泌,接下來的細菌代謝導致苷配基和脫硫酸鹽的產生,這兩種成分會造成膽汁酸的重吸收,引發疾病[19-21]。還有很多關于健康女性在月經期和排卵期胃腸性疾病增多的報道,他們認為這些情況與雌激素和黃體酮水平的改變密切相關。關于胃腸型疾病的發生與性激素分泌的關系方面恰恰也有相反的報道,但是這些都清楚表明二者存在必然聯系,而我們現在無法確定這種聯系是不是導致嬰幼齡組和孕期組聚類的原因[22,23]。很多研究證明腸道菌群狀態與宿主的能量代謝有關,或許這也是導致聚類的原因。與之前的報道不同[24,25],在我們的研究中未發現腸道菌群與性別之間的關系。
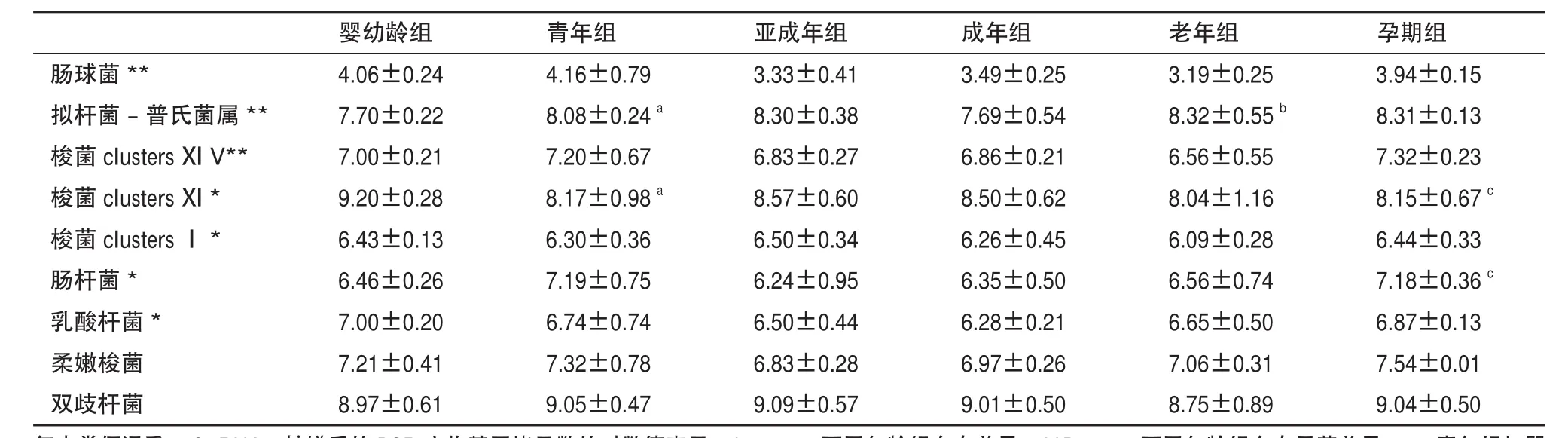
表2 不同年齡組中各個種屬細菌實時熒光定量q-PCR結果
Q-PCR結果顯示青年組與嬰幼齡組相比,擬桿菌和梭菌clusters Ⅺ差異顯著;老年組較之成年組,擬桿菌數量也顯著增多;與同齡亞成年和成年個體相比,孕期組梭菌clusters Ⅺ和腸桿菌數量有顯著差異,用以衡量腸道健康狀況的雙歧桿菌和腸桿菌比值(B/E值)[26]顯著低下。這些對于因斷乳等食性轉換、生理性免疫功能低下、生理性激素水平變化等造成的恒河猴腸道疾病具有一定的指示意義。
本研究中的母乳喂養期的嬰幼齡與孕期恒河猴腸道主要菌群特點相似尚屬首次發現,接下來我們會有針對性地展開研究,同時對患病個體樣品和健康個體進行比較,以期發現腸道菌群變化與疾病的潛在發生之間的規律。
4 結論
(1)以梭桿菌為主的厚壁菌門細菌是恒河猴腸道的主要優勢菌群,另外也有放線菌門、擬桿菌門和螺旋體門細菌的分布。
(2)恒河猴嬰幼齡組和孕期組出現明顯聚類,很可能與激素分泌密切相關。主成分載荷值顯示代謝相關功能菌R.bromii,健康密切相關益生菌Bif.dentium和Lac.helveticus為造成聚類的主要差異條帶。
(3)處于食性轉換期的青年組、免疫力下降的老年組以及體內激素水平發生變化的孕期組恒河猴腸道主要菌群水平有不同程度的改變。作為顯示腸道健康水平標準的雙歧桿菌和腸桿菌比值(B/E值)與同齡亞成年和成年組恒河猴相比,在孕期恒河猴有明顯的降低。
[1] Vaughan EE, Schut F, Heilig HG, et al.Amolecular view of the intestinal ecosystem[J].Curr Issues Intest Microbiol, 2000, 1:1-12.
[2] 朱偉云 姚文, 毛勝勇.變性梯度凝膠電泳法研究斷奶仔豬糞樣細菌區系變化[J].微生物學報, 2003, 43:503-508.
[3] Muyzer G, De Waal EC, Uitterlinden AG.Profiling of complexmicrobial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA[J].Appl Environ Microbiol, 1993, 59:695-700.
[4] Walker WA.Role of nutrients and bacterial colonization in the development of intestinalhost defense[J].J Pediatr Gastroenterol Nutr, 2000, 30(Suppl 2):2-7.
[5] Walter J, Hertel C, Tannock GW, et al.Detection of Lactobacillus,Pediococcus, Leuconostoc, and Weissella species inhuman feces by using group-specific PCR primers and denaturing gradient gel electrophoresis[J].Appl Environ Microbiol, 2001, 67:2578-2585.
[6] Byun R, Nadkarni MA, Chhour KL, et al.Quantitative analysis of diverse Lactobacillus species present in advanced dental caries[J].J Clin Microbiol, 2004, 42:3128-3136.
[7] Mckenna P, Hoffmann C, Minkah N, et al.Themacaque gutmicrobiome inhealth, lentiviral infection, and chronic enterocolitis[J].PLoS Pathog, 2008, 4:e20.
[8] Chen Y, Yang F, Lu H, et al.Characterization of fecalmicrobial communities in patients with liver cirrhosis[J].Hepatology, 2011,54:562-572.
[9] Thompson JR, Marcelino LA, Polz MF.Heteroduplexes inmixed-template amplifications:formation, consequence and elimination by reconditioning PCR[J].Nucleic Acids Res, 2002, 30:2083-2088.
[10] Kumar S, Tamura K, Nei M.MEGA:Molecular evolutionary genetics analysis software formicrocomputers[J].Comput Appl Biosci, 1994, 10:189-191.
[11] Pearson K.On lines and planes of closest fit to systems of points in space[J].Philosophical Magazine, 1901, 2:559-572.
[12] Muller AK, Westergaard K, Christensen S, et al.The effect of longtermmercury pollution on the soilmicrobial community[J].FEMS Microbiol Ecol, 2001, 36:11-19.
[13] Haarman M, Knol J.Quantitative real-time PCR analysis of fecal Lactobacillus species in infants receiving a prebiotic infant formula[J].Appl Environ Microbiol, 2006, 72:2359-2365.
[14] Collins MD, Lawson PA, Willems A, et al.The phylogeny of the genus Clostridium:proposal of five new genera and eleven new species combinations[J].Int J Syst Bacteriol, 1994, 44:812-826.
[15] Bartosch S, Fite A, Macfarlane GT, et al.Characterization of bacterial communities in feces fromhealthy elderly volunteers andhospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecalmicrobiota[J].Appl Environ Microbiol, 2004, 70:3575-3581.
[16] Palmer C, Bik EM, Digiulio DB, et al.Development of thehuman infant intestinalmicrobiota[J].PLoS biology, 2007, 5:e177.
[17] Sela D, Chapman J, Adeuya A, et al.The genome sequence of Bifidobacterium longum subsp.infantis reveals adaptations formilk utilization within the infantmicrobiome[J].Proc Natl Acad Sci USA, 2008, 105:18964.
[18] Koenig JE, Spor A, Scalfone N, et al.Succession ofmicrobial consortia in the developing infant gutmicrobiome[J].Proc Natl Acad Sci USA, 2011, 108(Suppl.1):4578-4585.
[19] Smith RV.Metabolism of drugs and other foreign compounds by intestinalmicroorganisms[J].World Review of Nutrition and Dietetics, 1978, 29:60.
[20] Goldman P.Biochemical pharmacology of the intestinal flora[J].Annu Rev Pharmacol Toxicol, 1978, 18:523-539.
[21] Grundmann O.The gutmicrobiome and pre-systemicmetabolism:current state and evolving research.J Drug Metabol Toxicol[J].2010, 1(2):104-111.
[22] Kennedy PJ, Clarke G, Quigley EM, et al.Gutmemories:towards a cognitive neurobiology of irritable bowel syndrome[J].Neurosci Biobehav Rev, 2012, 36:310-340.
[23] Moore J, Barlow D, Jewell D, et al.Do gastrointestinal symptoms vary with themenstrual cycle?[J].BJOG:An International Journal of Obstetrics & Gynaecology, 1998, 105:1322-1325.
[24] Adeyemo M, Spiegel B, Chang L.Meta-analysis:do irritable bowel syndrome symptoms vary betweenmen and women?[J].Alimentary pharmacology & therapeutics, 2010, 32:738-755.
[25] Altman G, Cain K C, Motzer S, et al.Increased symptoms in female IBS patients with dysmenorrhea and PMS[J].Gastroenterol Nurs, 2006, 29:4-11.
[26] Sokol H, Pigneur B, Watterlot L, et al.Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gutmicrobiota analysis of Crohn disease patients[J].Proc Natl Acad Sci USA, 2008, 105:16731-16736.
