蘋果輪紋病菌的多基因聯合鑒定
2013-09-13 11:47:10賈廣成周增強侯琿王麗朱建蘭
生物技術通報 2013年7期
關鍵詞:分析
賈廣成 周增強 侯琿 王麗 朱建蘭
蘋果輪紋病是蘋果生產中一種重要病害,主要危害蘋果枝干和果實,因其防治難、危害重,是目前我國蘋果生產的重要制約因素之一[1]。ITS基因,EF-1α基因和β-tubulin基因的 cDNA 序列已經從多種真菌中分離并對其生物學功能進行了初步研究[2]。GUADET等將分子系統學方法引入鐮孢菌屬種鑒定,主要通過ITS-rDNA、EF-1α、β-tubulin 等手段對鐮孢菌進行系統學研究。微管是真核生物中普遍存在的結構,與微絲和中間纖維一起組成細胞質中三維網絡結構的骨架系統。微管的組成為微管蛋白,主要的兩種微管蛋白為A和B微管蛋白,A微管蛋白和B微管蛋白形成微管裝配的基本單位-微管蛋白異二聚體。1963年,Ledbetter和Porter最先報道在植物細胞中存在微管結構。在植物病菌中,報道了若干 ITS基因,EF-1α基因和β-tubulin基因[3,4]的相關序列,但對其生物學功能還未進行深入研究[5]。微管蛋白和轉錄因子是真菌基因組主要的基因,在病菌的侵染過程中具有非常重要的作用。開展微管蛋白和轉錄因子的研究將為闡明蘋果輪紋病菌在侵染過程中的致病機理提供理論依據[6]。為了鑒定蘋果輪紋病菌并明確其主要致病基因的存在狀況,從有代表性的蘋果種植區采集并分離10個蘋果輪紋病菌菌株,對引起蘋果輪紋病的病原進行鑒定,并對病原的主要致病基因的存在狀況進行初步研究,旨在達到聯合鑒定蘋果輪紋病菌的目的。
1 材料與方法
1.1 材料
從河南、遼寧和山東蘋果種植區選擇10株菌株。這些菌株采集于發病的枝干和果實,后經單孢或單菌絲分離得到,由中國農業科學院鄭州果樹研究所保存。
1.2 方法
1.2.1 致病力測定 接種材料為富士蘋果一年生枝條,接種地點為中國農業科學院鄭州果樹研究所南園。用 70%乙醇對接種區域表皮進行消毒,待乙醇揮發后取菌餅接于樹皮光滑部位,接后用沾有無菌水的脫脂棉浸縛在菌餅上,隨后立即纏上塑料條保濕。致病力測定試驗設3個小區,每個小區內每個菌株接種1棵蘋果苗,每棵苗接種3個枝,每個枝上接3個點,整個試驗重復兩次。致病力測定的接種時間為2012年6月20日,調查時間為2012年11月10日,調查每個接種點的瘤狀凸起的發生情況,并記錄發病等級,最后計算病情指數。
分級標準如下表1所示。

表1 病情指數分級標準
1.2.2 菌體準備 將菌種接種于PSA斜面培養基上,26℃培養3d,挑菌于100mL液體培養基中,在220r/min的搖床上于26℃培養3-4d,離心收集菌體,用無菌蒸餾水洗滌菌絲3次,保存于-20℃冰箱。
1.2.3 DNA提取 用無菌竹簽挑取菌絲體1g加入研缽中,并加入預先準備好的液氮,研磨至菌落分散至單個細胞,即無成團菌絲體存在。將研磨液吸入1.5mL的滅菌EP管中,加入100μL預熱至35℃的裂解液,并且震蕩搖勻5s,加入400μL的65℃預熱的抽提液,倒置震蕩5s。加入600μL的Tris飽和酚氯仿抽提液,混勻5s,12000r/min離心5min。取離心后的上清液入另一只滅過菌的EP管中,加入等體積的異丙醇離心沉淀。用70%的乙醇再洗滌一次,離心后倒出乙醇,風干EP管,至無酒精氣味后,加入30μL TE溶液(pH8.0)溶解,各菌種DNA樣品取出2μL用于0.8%的瓊脂糖凝膠電泳,用120 V電壓電泳30min后,在320nm紫外燈下觀測。最后存于-20℃的冰箱中備用。
1.2.4 基因擴增[7]采用引物ITS1和ITS4擴增rDNA ITS1+5.8+ITS2片段。采用引物728F和986R擴增EF-1α基因片段。采用引物Bt2a和Bt2b擴增β-tubulin基因片段。
引物 ITS1:5'-TCCGTAGGTGAACCTGCG-3',ITS4:5'-TCCTCCGCTTATTGATAT-3'。
引物 728F:5'-CATCGAGAAGTTCGAGAAGG-3',986R:5'-ACTTG-AAGGAACCCTTACC-3'。
引物 Bt2a:5'-GGTAACCAAATCGGTGCTGCTTTC-3',Bt2b:5'-ACCCTCCAGTGTAGTGACCCTTGGC-3'。
PCR 反應體系為50μL:ddH2O 33.5μL,10×PCR buffer 5μL,dNTP(2.5mmol/L)5μL,Taq 酶(5U/μL)0.5μL,引物11μL,引物21μL,模板4μL(50ng/μL)。反應條件為:94℃預變性 2min;94℃變性 30s,55℃退火30s,72℃延伸 40s,共35個循環;最后 72℃延伸 8min。擴增產物經1%的瓊脂糖電泳檢測后,由上海生工進行雙向測序,拼接好的序列用于序列分析。
1.2.5 序列比對和系統發育樹的構建 測得序列用Sequencing Analysis 5.2(Applied Biosystems)軟件反復進行校對,去除兩端不確定的序列后,在 GenBank中進行 BLASTn比較搜索,下載同源性序列;同時下載 GenBank 中Botryosphaeria 的ITS基因、EF-1α基因和β-tubulin基因序列進行分析。運用 MEGA5分別計算不同種及亞種之間的遺傳距離(p-distance)和堿基差異數目,并構建系統發育樹[2]。
2 結果
2.1 蘋果輪紋病菌株情況蘋果輪紋病菌株情況如表2所示。
2.2 序列相似度比較
將PCR 產物電泳后于紫外透射下觀察,分別在約540bp、440bp與250bp出現一條清晰的cDNA條帶。DNAMAN軟件分析將所測得的ITS基因、β-tubulin基因和EF-1α基因序列通過查詢NCBI網站,對序列進行比對。
病原菌的rDNA-ITS序列測定結果菌株經過測序后獲得約540bp的DNA序列,GenBank 檢索結果(表3)表明,HX11,PY19和ZZ42菌株ITS序列與Botryosphaeria dothidea的同源性達到100%,其它菌株同源性均為99%。
病原菌的β-tubulin基因序列經過測序后獲得約450bp的DNA片段,GenBank 檢索結果(表4)表明,全部菌株β-tubulin基因序列與Botryosphaeria dothidea的同源性達到99%。
病原菌的EF-1α基因序列測定結果菌株經過測序后獲得約250bp的DNA序列,GenBank 檢索結果(表5)表明,HL1、PY19、YC5、YT1和ZZ42菌株EF-1α基因序列與Botryosphaeria dothidea的同源性達到100%,HX11和YC6菌株菌株 EF-1α基因序列與Botryosphaeria dothidea的同源性達到99%,其它菌株同源性均為98%。

表2 蘋果輪紋病菌株在田間的病情指數
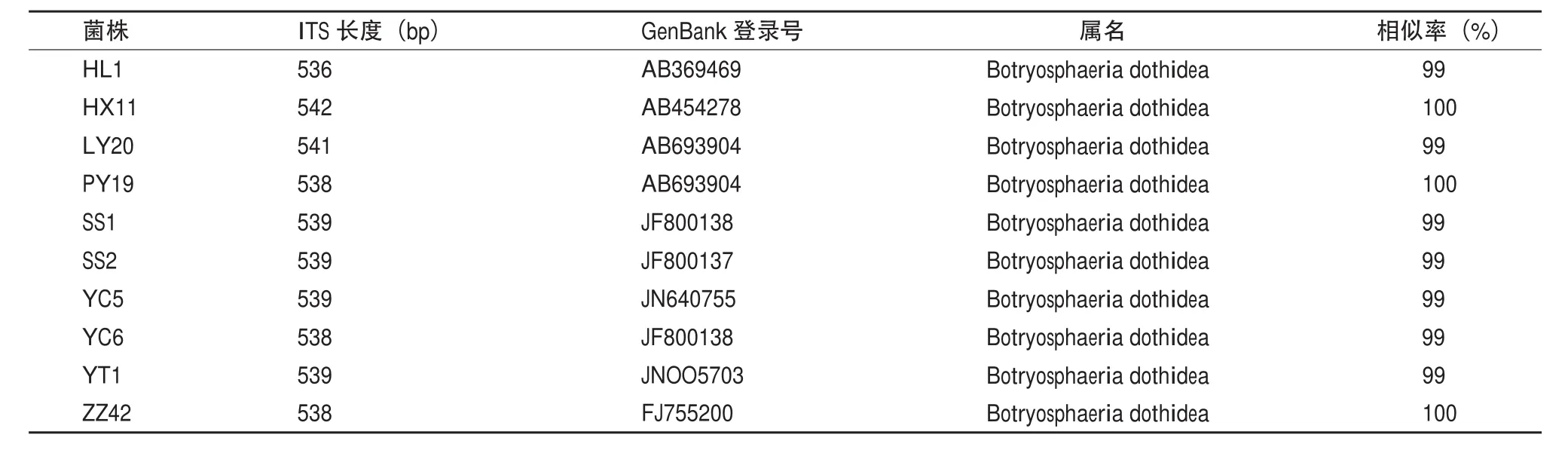
表3 蘋果輪紋病菌株ITS基因序列與GenBank中ITS基因序列相似度比較

表4 蘋果輪紋病菌株β-tubulin基因序列與GenBank中β-tubulin基因序列相似度比較

表5 蘋果輪紋病菌株EF-1α基因序列與GenBank中EF-1α基因序列相似度比較
2.3 菌種的的系統進化分析[8]
各菌種ITS基因、EF-1α基因和β-tubulin基因序列間的遺傳相似性極大,構建分子系統樹,系統樹上各分枝上的數值為經1000次Bootstrap后的置信度。初步探討了各菌種間的遺傳關系。
根據10個供試蘋果輪紋病菌的ITS基因序列和從NCBI上下載的4個其它菌株,利用DNAMAN軟件比對后經MEGA5.0軟件分析構建上述物種的系統進化樹。由系統進化樹(圖1)可以看出,比對為Botryosphaeria dothidea的10個供試蘋果輪紋病菌與同屬的 Botryosphaeria corticis、Botryosphaeria sp、Botryosphaeriamamane和Botryosphaeria iberica可以明顯的區分。系統進化樹與NCBI中比對的結果一致,表明了試驗結果的可信性,可以確定蘋果輪紋病菌的病原菌為Botryosphaeria dothidea。
10個供試蘋果輪紋病菌的β-tubulin基因序列和從NCBI上下載的3個其它菌株,利用DNAMAN軟件比對后經MEGA5.0軟件分析構建上述物種的系統進化樹。由系統樹(圖2)可見,所有菌株可分成兩大支,其中一支為HL1、YC6、SS2、LY20、PY19、YC5、ZZ42, 另 一 支 為 SS1、YT1、HX11、JN607129、JQ659299,DQ008335, 這 兩 支之間的親緣關系也很近。但在SS1、YT1、HX11、JN607129、JQ659299,DQ008335 這支中 SS1、YT1、HX11 和 Botryosphaeria rhodina、Botryosphaeria sp,Botryosphaeria corticis也是有發育上的分化。
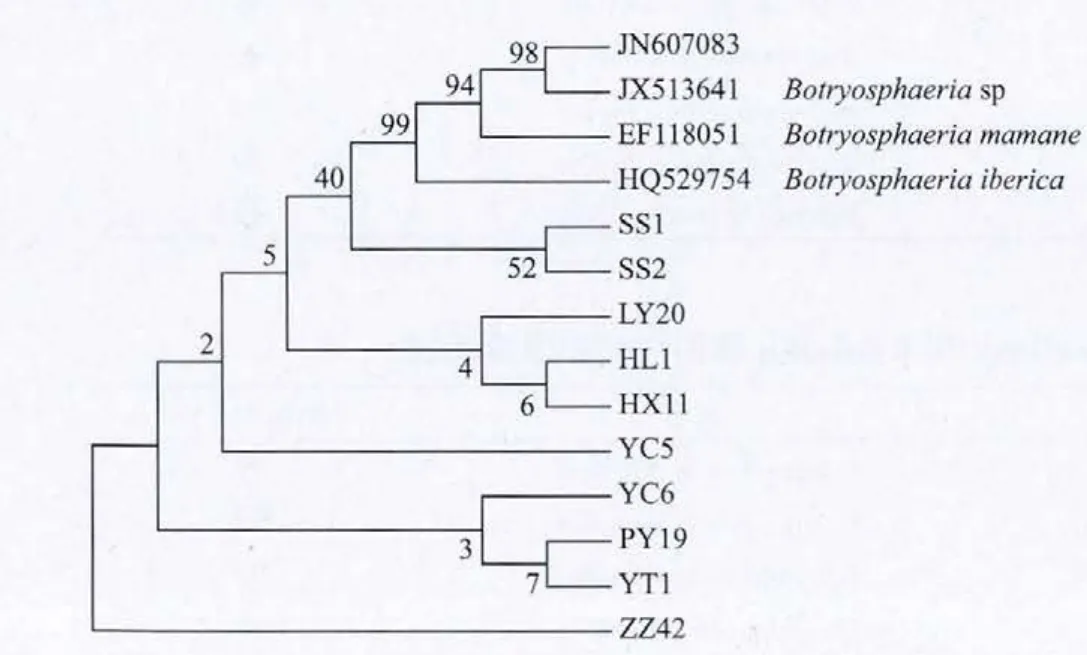
圖1 蘋果輪紋病菌ITS基因堿基序列構建的系統發育樹
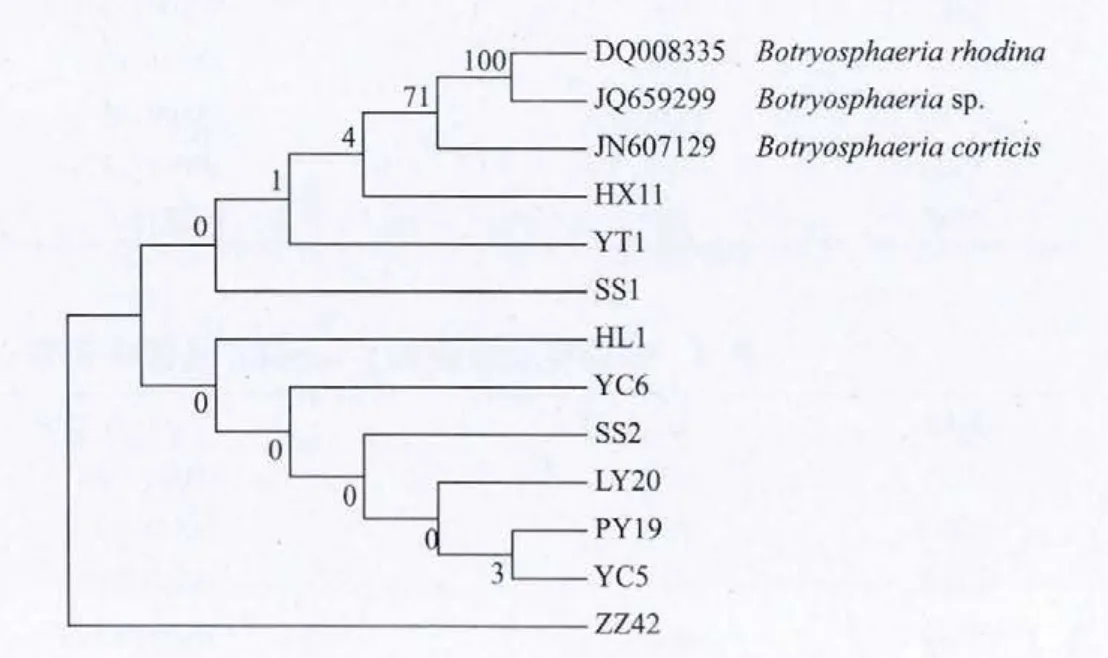
圖2 蘋果輪紋病菌β-tubulin基因堿基序列構建的系統發育樹
根據10個供試蘋果輪紋病菌的EF-1α基因序列和從NCBI上下載的3個其它菌株,利用DNAMAN軟件比對后經MEGA5.0軟件分析構建上述物種的系統進化樹。由系統進化樹(圖3)可以看出,比對為Botryosphaeria dothidea的10個供試蘋果輪紋病菌與同屬的 Botryosphaeria corticis、Botryosphaeria sp、Botryosphaeriamamane和Botryosphaeria iberica無法區 分。SS1、SS2和 HX11與 Botryosphaeriamamane的親緣關系更近。
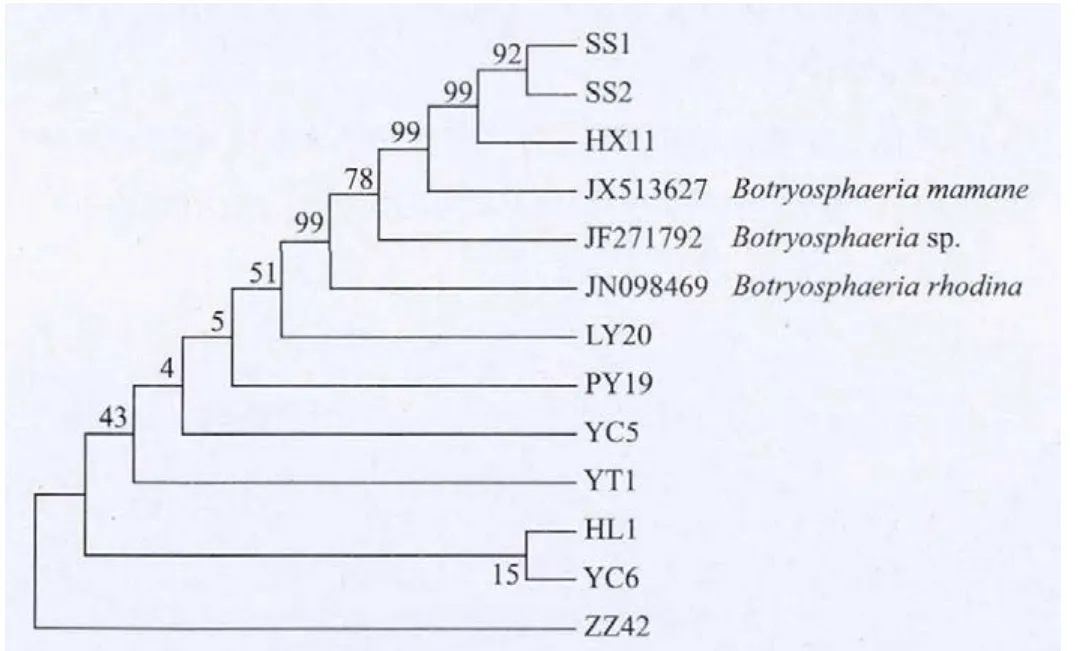
圖3 蘋果輪紋病菌EF-1α基因堿基序列構建的系統發育樹
3 討論
PCR技術是模擬自然界DNA復制過程,從而快速準確得到更多DNA樣本[9]。基因檢測是用于DNA序列的測定,基因表達譜鑒定和基因突變體的檢測和分析[10]。本研究中使用了DNAMAN和MEGA兩種軟件對蘋果輪紋病菌的遺傳關系進行分析,并且得到了相似的結果,說明以上兩種軟件均可使用。
隨著微管的發現,其諸多重要的生物學功能也被發現[11]。研究表明,微管與維持細胞形態、細胞的有絲分裂、細胞內物質運輸、細胞壁構建等植物重要的生理活動有著密切的聯系[12]。隨著分子生物學技術的發展,多種真核生物中的微管蛋白基因被擴增出來,微管已經成為生物學領域研究的熱點。微管蛋白基因核酸及氨基酸序列的比較分析研究表明,微管蛋白是生物進化中最為保守的蛋白質之一。植物中的微管蛋白高度保守。在植物細胞中引入不同植物物種的微管蛋白,外源微管蛋白可以被組合進入植物微管列陣,卻不影響其功能[13]。本研究克隆得到的10條β-tubulin基因序列和Botryosphaeria dothidea中的高相似性,同樣證明了蘋果輪紋病菌中B微管蛋白的高度保守性。目前,基于β-tubulin基因在真核生物中廣泛存在的特點,以及β-tubulin基因的植物系統發育學研究已取得的成績,β-tubulin基因將在蘋果輪紋病菌的系統發育和親緣關系研究中具有十分重要的意義。在進行有關核酸序列分析的時候,最好同時用不同的軟件進行分析,可以做到相互印證所分析的結果。
延伸因子EF-1α基因是一種編碼蛋白質的單拷貝核基因,在t RNA轉運到核糖體的過程中發揮作用,在研究系統發育方面具有重要作用,同時,EF-1α基因中也存在一些進化速度比較快的內元,因此它也可以分析低級階元的系統發育,許多學者對其在分子系統學上的應用已經做了大量的嘗試。Caterino等(2000)主張將 EF-1α基因作為分子系統學研究中的標準分子標記[14]。
4 結論
蘋果輪紋病菌的ITS基因序列相似性很高,蘋果輪紋病的病原菌是Botryosphaeria dothidea。證明了EF-1α基因和β-tubulin基因分化趨勢,這與測得的蘋果輪紋病菌田間致病力的分化相對應。EF-1α基因和β-tubulin基因與菌株的致病力有相關性,但EF-1α基因和β-tubulin基因復雜的分子作用機制還需進一步的深入研究。
[1] 蘇平, 周增強, 侯琿, 王麗, 朱建蘭.蘋果輪紋病菌對戊唑醇的敏感性檢測.果樹學報, 2010, 27(1):69-76.
[2] Yli-Mattilat T, Mach RL, Alekhinaia IA.Phylogenetic relationship of Fusarium langsethiae to Fusarium poae and Fusarium sporotrichioidesas and inferred by IGS, ITS, β-tubulin sequences and UP-PCRhybridization analysis[J].International Journal of Food Microbiology, 2004, 95:267-285.
[3] 戴芳瀾.中國真菌總匯[M].北京:科學出版社, 1979:1-1527.
[4] 黃勝雄, 胡尚連, 孫霞, 蔣瑤.慈竹B-tubulin 基因片段的克隆及序列分析[J].福建林業科技, 2009, 36(1):8-10.
[5] 李洪杰, 王曉鳴, 宋鳳景, 等.中國小麥品種對白粉病的抗性反應與抗病基因檢測[J].作物學報, 2011, 37(6):943-954.
[6] 董金皋, 農業植物病理學[M].北京:中國農業出版社, 2001,297.
[7] 王敏.基于PCR技術的基因檢測技術研究[J].生命科學儀器,2008, 6(3):34-36.
[8] 張琪, 薛慧良, 徐金會, 等.黑線倉鼠CRH 基因的編碼序列及系統進化分析[J].生物技術通報, 2012(10):88-94.
[9] 鄒志文, 陳芬, 余麗萍, 夏斌.幾種常見桔園螨類Wolbachia基因檢測與序列分析[J].生態學雜志, 2011, 30(3):547-551.
[10] 張羽, 王勝寶, 馮志峰, 張彥清.陜西省水稻種質資源的香味基因檢測[J].西北農林科技大學學報:自然科學版, 2011,39(4):55-59.
[11] 杜予州, 王莉萍, 陸亞娟, 等.美洲斑潛蠅不同寄主種群及地理種群間的β-tubulin 基因序列分析[J].中國農業科學,2008, 41(5):1542-1547.
[12] Snustad DP, Haas NA, Kopczak SD.The small genome of Arabidopsis contains at least nine expressed B-tubulin genes[J].Plant Cell, 1992, 4:549- 5561
[13] Villemur R, Haas NA, Joyce CM.Characterization of four new B-tubulin genes and their expression duringmale flower development inmaize(Zeamays L)[J].Plant Mol1Biol, 1994,24:295-315.
[14] 陳璐, 高增貴, 莊敬華, 等.玉米頂腐鐮孢菌rDNA-ITS和EF-1a基因序列及UP-PCR遺傳多樣性分析[J].沈陽農業大學學報, 2011, 42(1):31-36.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
