四種草本花卉的染色體核型分析
2013-12-23 05:11:50齊宇晴李新玲張彥妮李鳳
生物技術通報 2013年2期
齊宇晴 李新玲 張彥妮 李鳳
(1.哈爾濱師范大學生命科學與技術學院,哈爾濱 150025;2.東北林業大學園林學院,哈爾濱 150040)
草本花卉多為1-2年生,種類繁多、繁殖容易,同一種花卉又有多種品種和類型,株形、花形豐富多彩,并且病害少,適應性強,所以它們在庭院及城市綠化方面是極有價值的花卉,是美化生活不可缺少的花材[1,2]。本文主要研究了4種草花的核型,其中石竹屬于石竹科石竹屬,是我國傳統名花之一,觀賞期較長,而且含有皂苷、糖類及維生素、生物堿等,營養豐富,既可茶飲又可入藥[3]。觀賞椒又名五色椒,屬于茄科辣椒屬,花期為全年,觀賞椒含有多種氨基酸和維生素,不僅可以在其新鮮時直接食用,還可以腌制成泡菜,因此,好時椒的種植有助于解決北方冬季蔬菜種類少的問題[4]。裂葉花葵是錦葵科花葵屬的草本植物,盛開花徑10 cm左右,是著名的園林觀賞植物[5]。鳳仙花屬于鳳仙科鳳仙花屬,不僅供觀賞,同時又是一種良好的藥用植物。此外,鳳仙花對氟化氫反應靈敏,可以用來監測氟化氫污染[6]。
目前,國內外關于草花的生活習性、繁殖技術、栽培方法、經濟價值等方面的研究報道較多,但對于草花的細胞學研究基礎理論比較薄弱,絕大多數草花的染色體數目及其核型還沒有被廣泛研究。對草花技術開發,特別是增強其適應性、抵抗病蟲害、新品種選育等工作缺少理論支持和發展后勁。染色體是基因的載體,是生物進化發育、遺傳變異的物質基礎[7],對草花染色體的研究可以從細胞水平上了解遺傳變異的規律,研究草花不同科屬種之間在進化上的親緣關系[8],同時,對草花的抗病蟲害育種及新品種選育提供必要的細胞學資料。
1 材料與方法
1.1 材料
供試用的石竹、裂葉花葵、觀賞椒和鳳仙花的種子均來自于東北林業大學園林學院。
1.2 方法
選成熟的種子放在培養皿中加厚濾紙、水與少量的殺菌劑,21℃處理24-36 h,將發芽種子置于8℃下進行48 h低溫處理,芽略有增長,然后將種子轉入22.5-23℃條件下,處理12 h,使芽迅速生長。將上述處理的根尖剪下1 cm左右,立即放入有冰水混合物的試管中處理24 h,再用Carnoy固定液(無水乙醇∶冰醋酸=3∶1)固定24 h,水洗后放入1%的醋酸洋紅中染色。常規壓片法壓片,并在Lecia DM6000B型顯微鏡下觀察,選取圖像清晰、中期染色體分散較好的細胞用Lecia DFC480采集圖像。參照李懋學和陳瑞陽[9]的標準進行染色體計數和測量,得到核型數據;參照Levan等[10]的標準進行染色體類型分析;按照Stebbins[11]的分類標準進行核型分類;核型不對稱系數按Arano[12]的方法計算,比值越大越不對稱。
2 結果
2.1 石竹的染色體數目與核型
選擇30個染色體分散良好的細胞觀察計數,其中29個細胞染色體數目為14條,占細胞總數的97%,因此確定石竹體細胞染色體數目為2n=14(圖1),觀察中未發現染色體有隨體。石竹的7對染色體中有6對是中部著絲點,一對為近中部著絲點,其核型公式為2n=2x=14=12 m+2 sm。染色體相對長度變異范圍為4.13-5.33,最長染色體與最短染色體的比值為1.29,臂比大于2∶1的染色體比率為0,核不對稱系數(AS.K%)為54.84%,根據Stebbins(1971)的分類標準,判定其核型為“1A”型(表1)。
2.2 裂葉花葵的染色體數目與核型
選擇30個染色體分散良好的細胞觀察計數,全部30個細胞染色體數目均為14條,占細胞總數的100%,未發現非整倍性變異現象和多倍體現象,因此確定裂葉花葵體細胞染色體數目為2n=14(圖2)。裂葉花葵的7對染色體中,具近中部著絲點的是第2、3、4、6對;中部著絲點的染色體是第1、5、7對,裂葉花葵的核型中沒有帶有隨體的染色體,其核型公式是2n=2x=14=6 m+8 sm。染色體相對長度變異范圍為5.04-8.62,最長染色體與最短染色體的比值為1.71,臂比大于2∶1的染色體有3對,核不對稱系數(AS.K%)為64.97%,根據Stebbins(1971)的分類標準,判定其核型為“2A”型(表2)。

圖1 石竹的染色體中期相和核型
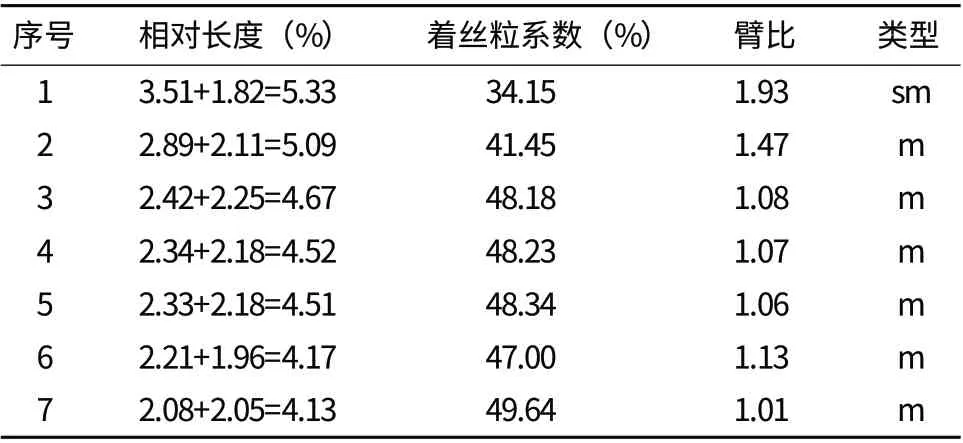
表1 石竹核型參數表

圖2 裂葉花葵的染色體中期相和核型

表2 裂葉花葵核型參數表
2.3 觀賞椒的染色體數目與核型
選擇30個染色體分散良好的細胞觀察計數,全部30個細胞染色體數目均為24條,占細胞總數的100%,因此確定觀賞椒體細胞染色體數目為2n=24(圖3)。觀賞椒的12對染色體均為中部著絲點,其核型公式為2n=2x=24=24 m。染色體相對長度變異范圍為5.24-11.28,最長染色體與最短染色體的比值為2.15,臂比大于2∶1的染色體比率為0,核不對稱系數(AS.K%)為53.30%,根據Stebbins(1971)的分類標準,判定其核型為“1B”型(表3)。

圖3 觀賞椒的染色體中期相和核型

表3 觀賞椒核型參數表
2.4 鳳仙的染色體數目與核型
選擇30個染色體分散良好的細胞觀察計數,其中29個細胞染色體數目為14條,占細胞總數的97%,因此確定鳳仙體細胞染色體數目為2n=14(圖4),觀察中未發現染色體有隨體。鳳仙的7對染色體具中部著絲點的染色體是第1、4、5、6、7對,近中部著絲點的是第2、3對,其核型公式為2n=2x=14=10 m+4 sm。染色體相對長度變異范圍為4.87-10.31,最長染色體與最短染色體的比值為2.12,臂比大于2∶1的染色體比率為0,核不對稱系數(AS.K%)為60.78%,根據Stebbins(1971)的分類標準,判定其核型為“1B”型(表4)。

圖4 鳳仙花的染色體中期相和核型

表4 鳳仙核型參數表
3 討論
石竹、裂葉花葵、觀賞椒及鳳仙花等都是我國北方廣泛種植的草本觀賞花卉,但目前我國對這些草花的研究僅限于生活習性、繁殖技術、栽培方法等方面,而對它們的染色體數目、染色體形態結構特征等方面的研究幾乎沒有。核型是生物體最穩定的細胞學特征之一,與外部形態性狀相比,核型受外界環境因素影響較小,而能保持相對穩定。而且還可以根據核型分析來判斷物種間的親緣關系,推測物種在進化過程中彼此的分化歷程[9]。核型相似程度往往與物種同源性和親緣關系存在一致性,物種間在分類階梯上越遠,它們的核型差異越大[13]。Stebbins[11]認為,在生物的進化過程中,染色體核型是由對稱性向非對稱性演化的,核型對稱性程度越高的生物,其染色體變異越小,進化程度也越低;而非對稱性程度越高的生物,其染色體變異越大,進化程度越高。本研究中4種草花的染色體核型類型分別為1A、2A、1B、1B,是對稱程度較高的染色體組成,因此這4種觀賞花卉在進化上也都是比較保守的。由此,對這些花卉的染色體核型進行研究,不僅對它們的遺傳和育種工作是一個極大的促進,而且對它們的演化、地理分布的研究也具有較大的推動作用。
對染色體核型進行分析的前提是要獲得一張好的染色體制片,而獲得好的染色體片子的基礎是取材,關鍵是對材料進行預處理。預處理成功則整個染色體制片工作便成功了一大半。可以利用藥物進行預處理,如秋水仙素、對二氯苯、8-羥基喹啉等。也可以采取低溫的方法進行預處理。本試驗采用低溫24 h,染色體長度適中,形態良好。在處理的過程中一定要保證冰水混合物的狀態,處理時間過長,染色體高度濃縮,不利于染色體的識別。低溫處理的方法簡便易行、效果良好。
4 結論
本試驗研究結果表明,石竹染色體數2n=14,核型公式為K(2n)=14=12 m+2 sm,其核型為“1A”;裂葉花葵染色體數2n=14,核型公式為K(2n)=14=6 m+8 sm,核型為“2A”;觀賞椒染色體數2n=24,核型公式K(2n)=24=24 m,核型為“1B”;鳳仙染色體數2n=14,核型公式為K(2n)=2x=14=10 m+4 sm,核型為“1B”。此4種草本花卉的染色體不對稱系數(AS.K%)為53.30%-64.97%,屬于對稱程度較高的類型。根據Stebbins[14]有花植物核型進化中“對稱-原始、不對稱-進化”的觀點,可認為該4種草花的核型進化程度較低,屬于比較保守的進化類型。
[1] 劉志虎,陳江,于永武. 草本花卉繁種、制種、引種試驗研究初報[J]. 林業實用技術,2012,3:44-46.
[2] 劉宏濤. 草本花卉栽培技術[M].北京:金盾出版社,1999:2-28,70-161.
[3] 田永久. 石竹的繁殖與栽培管理[J]. 花木盆景:花卉園藝版,2010(9):11.
[4] 余文中,劉崇政,楊紅,等. 觀賞辣椒種質材料主要園藝性狀差異性分析[J]. 湖南農業科學,2010(19):7-9.
[5] 吉醒. 多姿多彩的錦葵科植物[J]. 花木盆景:花卉園藝版,2010(4):2-3.
[6] 潘繼蘭. 鳳仙花高效栽培技術[J]. 花木盆景:花卉園藝版,2010(3):9.
[7] 侯小改,段春燕,劉素云,等 . 中國牡丹染色體研究進展[J]. 中國農業通報,2006,22(2):317-319.
[8] 楊漢民. 細胞生物學實驗[M].第2版.北京:高等教育出版社,1997.
[9] 李懋學,陳瑞陽. 關于植物核型分析的標準化問題[J]. 武漢植物學研究,1985,3(4):297-302.
[10] Levan A,Fredga K,Sandberg AA. Nomenclature for centromeric position on chromosomes[J].Hereditas,1964,52:201-220.
[11] Stebbins GL. Chromosomal and evolution in higher plants[M].London:Edward Arnold,1971.
[12] Arano H. Cytological studies in subfamily Carduoideae(Compositae)of Japan[J]. Botanical Magazine of Tokyo,1963,76:32-39.
[13] 陳瑞陽,宋文芹,李秀蘭,等.中國主要經濟植物基因組染色體圖譜(第3冊)[M].北京:科學出版社,2003:1-52.
[14] Stebbins GL.植物的變異和進化[M].復旦大學遺傳研究所,譯.上海:上海科學技術出版社,1950.
