寧夏中部干旱帶主要植物葉性狀變異特征研究
2014-01-02 06:00:20陳林楊新國宋乃平楊明秀肖緒培王興
草業學報 2014年1期
陳林,楊新國,宋乃平,楊明秀,肖緒培,王興
(寧夏大學西北退化生態系統恢復與重建教育部重點實驗室西北土地退化與生態恢復省部共建國家重點實驗室培育基地西部生態與生物資源開發聯合研究中心,寧夏銀川750021)
長期以來,人們習慣采用氣體交換測定指標去理解植物生長過程(忽略了一些植物性狀指標的重要性)[1],但是這種瞬間變化的測定指標(如光合/呼吸速率)是難以深入解釋植物生長的長期適應機理[2]。葉片是植物與外界環境進行物質與能量交換的主要器官,直接影響到植物的基本行為和功能,且隨著氣候、土壤和環境因子的變化而變化[3],葉性狀在特定的環境背景下保持穩定,反映了不同物種在長期進化過程中對其生存環境的適應和競爭能力,還可以反映植物適應環境變化所形成的生存對策[4],因而具有重要的生態學和生物進化意義[5],對植物葉性狀的研究能更好地闡明葉片生理生態對環境變化的響應機理[6]。目前,植物葉性狀的變化與環境的關系成為植物生態學的研究熱點之一[7],應用葉性狀研究植物對環境的適應機理更是生理生態學領域近些年研究中新的突破點[3]。
寧夏中部干旱帶降水量稀少,氣候惡劣,植被稀疏,由于風沙危害和超載過牧等自然和人為因素的共同作用,寧夏荒漠草原土壤沙化、植被退化嚴重[6],總體表現為生物多樣性降低、草地生產力下降、荒漠化不斷擴大和風沙危害日益加劇。近年來實施的“退耕還林(草)”和“封山禁牧”等政策雖然取得了一定成效,但植被建設中仍存在成活率低、保存率低等問題,其中一個重要原因就是對該區植物適應環境的機理研究還不夠深入,這無疑使當前生態環境保護和建設陷入盲目。因此,認清干旱地區,特別是北方農牧交錯干旱地區植物的基本性狀,不但可以反映植物適應環境變化所形成的生存對策,而且可以深入解釋植物對環境的適應機理,對該地區植被重建與恢復具有重要的理論和實踐意義。此外,目前針對該地區不同植物葉性狀的研究并不多見。基于此,本研究以寧夏中部干旱帶的典型區域中在相同生境條件下的人工引入種及野生物種植物為研究對象,從植物葉性狀入手,研究不同種植物葉性狀間的差異,分析葉性狀之間的關系,以及不同科類群、功能類群植物葉性狀間的差異,試圖從植物葉片生理生態的角度揭示該地區植物對環境的適應性,為人工調控促進植被演替、退化生態系統的植被恢復、人工建置草地、選擇合適的植物種進行荒漠化防治等提供理論依據,并為進一步研究該地區的生物多樣性和生境的穩定性保育提供基礎資料。
1 材料與方法
1.1 研究區自然概況
寧夏鹽池縣皖記溝行政村(楊寨子自然村和北王圈自然村),處于寧夏中部干旱帶上,位于鹽池縣城東北約3 km,地貌為鄂爾多斯緩坡起伏高原。氣候特點為干旱少雨,蒸發量大,冬春兩季風大沙多,屬典型的中溫帶大陸性氣候。1954-2010年年平均氣溫為8.46℃,年降水量為276.3 mm,年日照時數為2862.6 h。地表植被屬荒漠草原,沙生特征明顯。土壤以有機質含量低、易沙化的淡灰鈣土和風沙土為主。地表水與地下水資源匱乏。20世紀60年代以來,由于人類活動加劇,土地沙化面積仍不斷擴大[8]。
1.2 研究方法
1.2.1 實驗材料的選擇 選取夏季生長的35種植物(表1)作為研究對象,于植被生長盛期(2012年8月份)一次降雨后的第3~5天,選取完全展開、保持完整且生長良好的成熟葉片(或其著生小枝)作為實驗材料。
1.2.2 植物葉性狀測定方法 考慮到水對植物葉片大小有明顯的影響[9],在計算過程中會影響到結果的準確性,因此,植物葉片吸水實驗參照Limm等[10]的方法,改進后進行操作。將植物葉片(或其著生小枝)剪下,迅速在切口沾涂凝固膠后裝入塑封袋,放入智能移動保溫冰箱(52L/AMC017A),然后帶回實驗室,立即用掃描儀(清華同方A868)掃描成圖片,應用Photoshop計算吸水面積,記為S。吸水實驗的具體步驟是:將掃描葉面積后的材料稱重,記為m1,在黑暗中完全浸沒于去離子水中直至飽和,取出后用干潔的白布將附著在表面的水分吸取后稱重,記為m2,在空氣中放置干燥失水10 min后稱重,記為m3,然后再次浸入去離子水中1 s取出,用干潔的白布將附著在表面的水分吸取后稱重,記為m4,然后將材料放入硬質信封中,105℃殺青30 min,65℃烘干至恒重,記為m5。每一物種重復5~9次。
葉片單位面積吸水量(leaf water uptake content,LWUC,mg/cm2)的計算公式為:
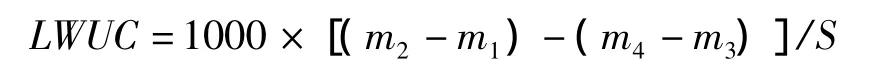
葉片含水量增加率(percentage increase in leaf water content,RW,%)的計算公式為:
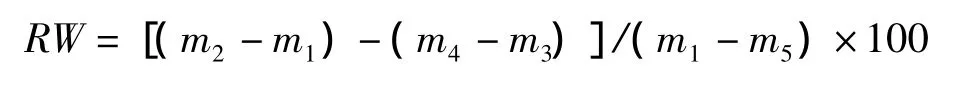
葉片水分飽和虧(leaf water saturation deficit,WSD,%)的計算公式為:
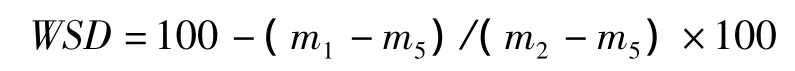
比葉面積(specific leaf area,SLA,cm2/mg)的計算公式為:
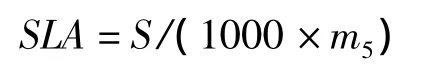
葉片干物質含量(leaf dry material content,LDMC,%)的計算公式為:
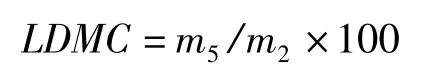
葉片肉質化程度(leaf succulence,Suc,g/cm2)的計算公式為:

葉組織密度(leaf density,LD,g/cm3)的計算公式為:
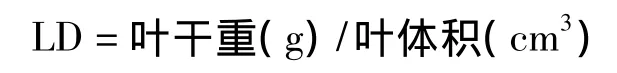
式中,葉體積=葉面積×葉厚度。
葉片厚度(leaf thickness,LT,mm)測定方法為:考慮到野外采樣的可操作性,本研究中隨機取不同葉位9個葉片,用游標卡尺(精度為0.01 mm)測量新鮮葉片的厚度后取平均值。
用便攜式葉綠素儀(USA,SPAD 502DL Plus)對可進行測量植物種的葉片葉綠素含量(chlorophyll content,ChlC,%)進行測定,每一物種重復9次。

表1 植物名錄及功能群分類Table 1 List of the species and their ownership to life forms and functional groups
1.3 數據分析
箱式圖用于多組數據平均水平和變異程度的直觀分析比較,包括最小值、最大值、中位數、兩個四分位數(上四分位數和下四分位數)以及異常值(實心黑圓點)和極端值(*),故本研究采用箱式圖表現不同科類群、功能類群間的差異;采用SPSS 17.0軟件進行單因素方差分析(one-way AVOVA),LSD法檢驗處理組間的差異顯著性。
2 結果與分析
2.1 植物葉性狀的變異特征
表2為研究地植物 LT、Suc、LD、SLA、LDMC、WSD、ChlC、LWUC、RW 的平均值、最值、中值以及變異特征值。按照變異系數,將9種葉性狀劃分為3類:高變異系數(≥70%)、中變異系數(40% ~70%)和低變異系數(≤40%)。35種植物LT和LWUC的平均值分別為0.3605 mm和6.9626 mg/cm2,變異系數高達83.90%和81.10%,其中禾本科的畫眉草LT最小(0.1340 mm),菊科的菊芋LT最大(1.8340 mm),而薔薇科的二裂委陵菜LWUC最小(1.3134 mg/cm2),蒺藜科的白刺LWUC最大(22.2356 mg/cm2)。中變異系數的RW的平均值為34.8357%,變異系數為57.73%,其中豆科的中間錦雞兒最小(8.9829%),豆科的砂珍棘豆最大(104.9063%);LD的平均值為0.2730 g/cm3,變異系數為48.49%,其中菊科的菊芋最小(0.0393 g/cm3),豆科的草木樨狀黃耆最大(0.5999 g/cm3);Suc的平均值為0.0362 g/cm2,變異系數為47.07%,其中禾本科的畫眉草Suc最小(0.0169 g/cm2),蒺藜科的駱駝蓬最大(0.0996 g/cm2);SLA的平均值為0.1582 cm2/mg,變異系數為43.06%,其中禾本科的針茅最小(0.0483 cm2/mg),禾本科的虎尾草最大(0.2963 cm2/mg)。低變異系數中,可測量植物27種ChlC的變化范圍在9.8000%(禾本科的狗尾草)至53.7700%(薔薇科的蘋果樹)之間,平均值為33.8134%,變異系數為39.84%;LDMC的變化范圍在11.2999%(豆科的苦參)至40.7958%(薔薇科的杜梨)之間,平均值為22.5503%,變異系數為34.28%;WSD的變化范圍在14.7490%(禾本科的中亞百草)至57.0748%(豆科的砂珍棘豆)之間,平均值為37.1261%,變異系數為27.61%。
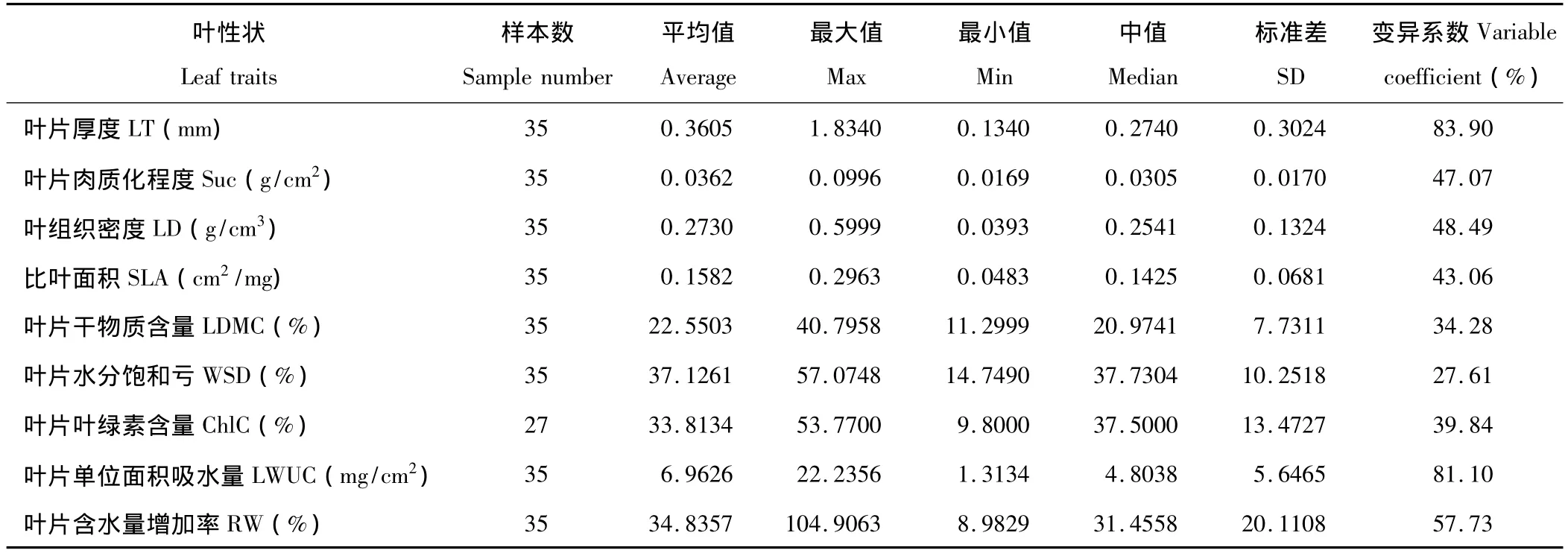
表2 研究地植物葉性狀的變異特征Table 2 Parameters of leaf traits in study area
2.2 葉性狀的相關性分析
表3為植物各葉性狀之間的相關系數及顯著水平。可以看出,LT與LD(r=-0.516,P<0.01)呈極顯著的負相關關系,與 Suc呈顯著的正相關關系(r=0.420,P <0.05);Suc與 SLA(r= -0.563,P <0.01)呈極顯著正相關關系,與 LWUC(r=0.675,P <0.01)呈極顯著正相關關系;LD 與 SLA(r= -0.337,P <0.05)呈顯著負相關關系,與ChlC和LDMC呈正相關關系,并分別達到了顯著水平(r=0.401,P<0.05)和極顯著水平(r=0.542,P<0.01);SLA 與 LDMC(r= -0.468,P <0.01)和 ChlC(r= -0.676,P <0.01)呈極顯著的負相關關系,與 LWUC 存在顯著的負相關關系(r= -0.350,P <0.05);LDMC 與 WSD(r= -0.174,P >0.05)、LWUC(r= -0.328,P >0.05)、RW(r= -0.123,P >0.05)呈負相關,但相關性均不顯著(P >0.05),與 ChlC 呈顯著正相關(r=0.428,P <0.05);WSD 與 LWUC(r=0.395,P <0.05)和 RW(r=0.572,P <0.01)呈極顯著的正相關關系;LWUC 與 RW 呈極顯著的正相關關系(r=0.706,P <0.01),與 ChlC 呈正相關關系,但相關性不顯著(r=0.280,P >0.05);RW 與ChlC 正相關關系不顯著(r=0.164,P >0.05)。
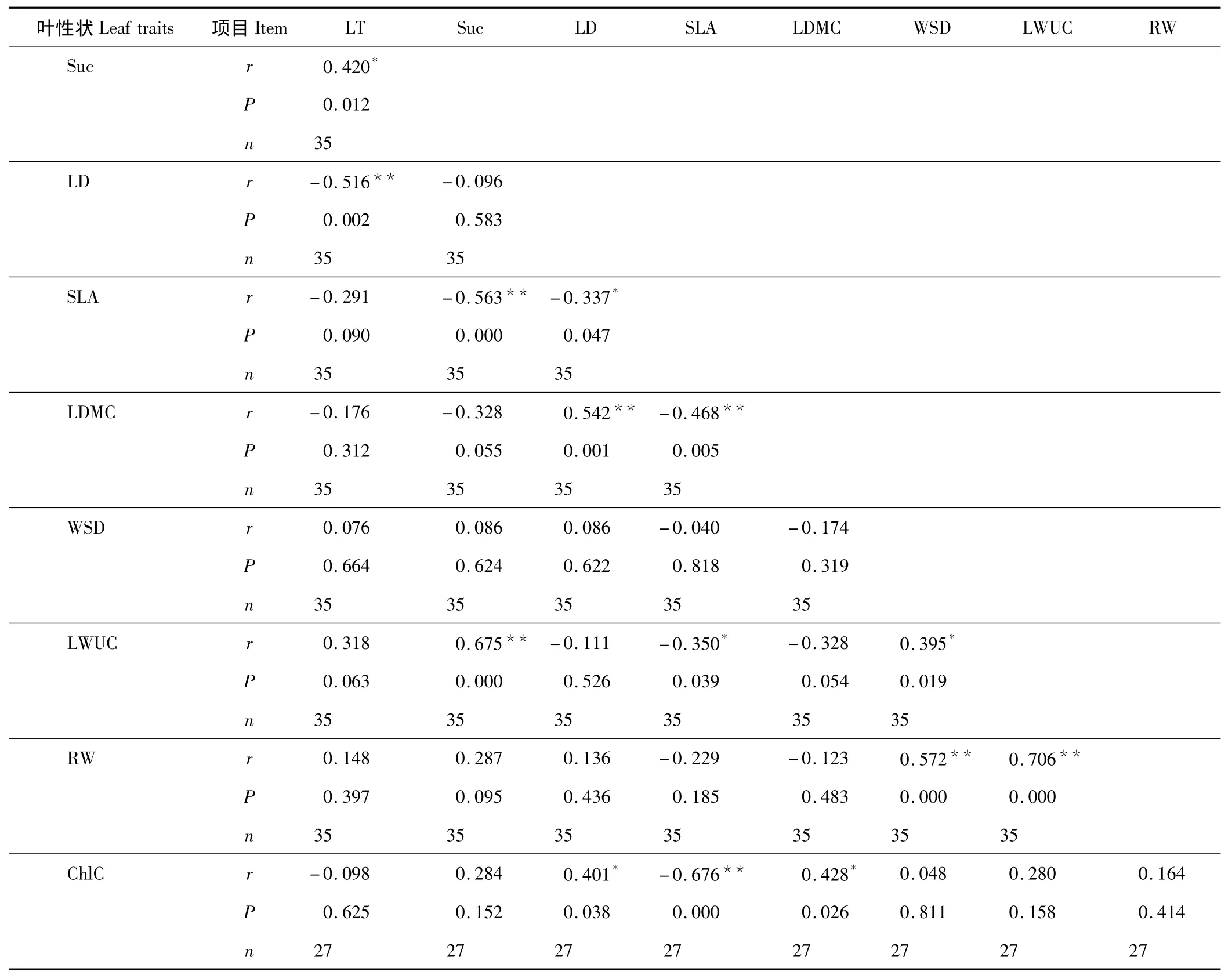
表3 植物葉性狀的相關分析Table 3 Correlation coefficient between six leaf water uptake strategy of 35 plants
2.3 不同科類群植物葉性狀比較
方差分析結果表明(圖1),植物葉性狀LD、LT、Suc、LDMC和LWUC在不同科類群之間存在極顯著差異(P<0.01),但SLA、WSD、ChlC和RW之間則差異不顯著(P>0.05)。薔薇科植物葉片的LD最高,極顯著大于豆科、禾本科、蒺藜科和菊科植物葉片的LD(P<0.01),菊科植物的LD極顯著的小于豆科和禾本科(P<0.01),蒺藜科、豆科和禾本科植物葉片的LD之間則無顯著差異(P>0.05)。菊科植物葉片的LT極顯著的大于豆科、禾本科、蒺藜科和薔薇科植物葉片的LT(P<0.01),而豆科、禾本科、蒺藜科和薔薇科植物葉片的LT之間則無顯著差異(P>0.05)。蒺藜科植物葉片的Suc極顯著高于豆科、禾本科和薔薇科植物葉片的Suc(P<0.01),與菊科植物葉片的Suc無顯著差異(P>0.05),而菊科植物葉片的Suc極顯著高于禾本科植物葉片的Suc(P<0.01),與豆科、薔薇科植物葉片的Suc無顯著差異(P>0.05)。薔薇科植物葉片的LDMC極顯著的大于豆科、蒺藜科和菊科植物葉片的LDMC(P<0.01),與禾本科植物無顯著差異(P>0.05),而禾本科植物葉片的LDMC極顯著的大于蒺藜科和菊科植物葉片的LDMC(P<0.01),與豆科植物葉片的LDMC無顯著差異(P>0.05),豆科、蒺藜科和菊科植物葉片的LDMC之間則無顯著差異(P>0.05)。蒺藜科植物葉片的LWUC極顯著的大于豆科、禾本科和薔薇科植物葉片的LWUC(P<0.01),與菊科植物葉片的LWUC無顯著差異(P>0.05),而豆科、禾本科和薔薇科植物葉片的LWUC也無顯著差異(P>0.05)。
2.4 不同功能類群植物葉性狀比較
圖2是對一年生和多年生植物葉性狀的比較。經方差分析結果表明,一年生植物LD、ChlC和LDMC的平均值極顯著低于多年生植物LD、ChlC和LDMC的平均值(P<0.01),而SLA則極顯著的高于多年生植物(P<0.01)。但一年生和多年生植物LT、Suc、WSD、LWUC和RW的平均值之間無顯著差異(P>0.05)。

圖1 不同科類群植物葉性狀比較Fig.1 Comparison of leaf traits among different family groups
3 結論與討論
有學者提出了基于植物性狀的植物群落構建理論:對特定的生境,構成群落的物種雖存在個體差異,但與其他群落物種相比,群落內物種間具有相似的性狀特征,即性狀趨同[11]。但隨著對植物功能性狀與植物群落形成及生態系統功能關系研究的深入,有研究表明,即使在相似生境中,葉片的一些特征仍存在種間差異[12],本研究的結果也證實了這一點(表2),部分葉性狀(LT、LWUC、RW、LD、Suc、SLA)存在著較大的差異(具有中、高變異系數)。此外,本研究中所測植物的各葉片形狀指標的平均值與其他地區研究結果[13]相比有所差異,造成這種現象的原因可能是不同生境植物長期適應生態環境的結果以及與其本身固有的遺傳特性有關[14],也可能與所選物種的不同以及測定季節的差異有關。SLA和LDMC是植物葉片的兩個關鍵性狀,能夠較好地反映植物生長策略及植物利用資源的能力、生物量大小和生態系統功能,在退化荒漠草原地區植物SLA能更好地指示植物對資源的利用[6],本研究中植物葉片SLA和LDMC的平均值均相對偏小,這與本研究區氣候干旱、土壤貧瘠的環境條件有關,反映了該地區植物對有限資源的利用能力、適應性,能很好地適應貧瘠的環境。
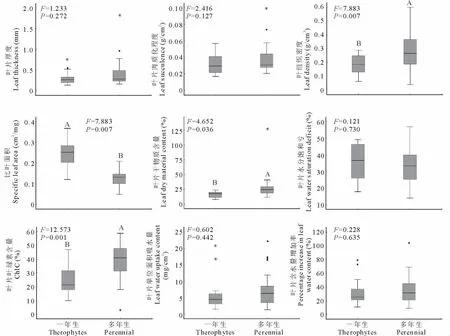
圖2 不同功能類群植物葉性狀比較Fig.2 Comparison of leaf traits between annual and perennials
關于植物葉性狀之間關系的研究已經有許多報道。葉性狀并不是孤立發揮作用的,植物在長期適應環境過程中,通過內部不同功能之間的調整,最終形成一系列適應某種環境的功能性狀的組合[15],它們之間是彼此聯系的。有學者對全球175個樣點2548種植物[3]以及中國草地174個樣點171種植物[16]的葉性狀分析后,研究結果證實了葉性狀之間普遍存在著密切的關系。這是由于植物生長過程中,由于受到物理、生理等的綜合作用,不同的性狀之間表現出一定的相關性,從而反映出植物對環境的趨同適應特征。但是由于植物的葉性狀受環境的影響很大,因此,不同研究區域植物葉性狀之間的關系存在一定的差異,基于寧夏中部干旱帶35種植物葉性狀的測定和分析結果,與鄭新軍等[17]在準噶爾盆地的研究結果不盡相同,也證明了這一點。這些差異通常被解釋為植物遺傳特性[18]或適應環境的結果[19]。幾乎所有的實驗研究結果都表明,植物的SLA和LDMC之間呈負相關關系,隨著LDMC的增加,植物葉片含水率降低,LD增加,從而導致SLA降低[20-21]。但是由于植物葉性狀對環境因子的應對措施不完全一致,不同植物SLA和LDMC在物種水平上的相關性也不盡相同[22]。李玉霖等[23]研究認為,流動沙丘、半固定沙丘植物的SLA和LDMC之間相關性均不顯著,但流動沙丘、半固定沙丘和固定沙丘的所有植物SLA和LDMC之間卻有顯著負相關關系。
一年生草本植物在冬天全部死亡,所以冬天的維持消耗為0;多年生草本植物在冬天僅地下部分能存活,其維持消耗顯然大于一年生草本植物。由于一年生植物和多年生植物在獲取資源的能力以及維持生態系統穩定性方面具有不同的生態功能,而植物的葉片性狀是植物行為和功能的具體體現[19]。研究發現,一年生植物和多年生植物的葉性狀存在較大差異。寧夏中部干旱帶一年生植物的SLA極顯著大于多年生植物的SLA(P<0.01),而LMDC則顯著小于多年生植物(P<0.05),這也驗證了SLA和LMDC之間的顯著負相關關系是很多陸地生態系統類型的普遍特征[24]。因此可以用來推斷彼此,如一年生植物的SLA顯著偏低,則可以推斷一年生植物的LMDC會顯著偏大。
目前,關于不同科類群間植物葉性狀的系統報道尚不多見。本研究發現,植物葉性狀LD、LT、Suc、LDMC和LWUC在不同科類群之間存在極顯著差異(P<0.01),而SLA、WSD、ChlC和RW之間則差異不顯著(P>0.05)。因此可以將LD、LT、Suc、LDMC和LWUC用作不同科類群植物間相比較的研究因子,進而對相關研究提供方便。
最后,有學者提出了一個“全球統一的葉性狀間的比例關系”[25]的假說,然而,目前國內外對葉性狀間的關系在不同空間尺度上的比較研究相對還很少[26],針對不同尺度葉性狀間關系是否有差異,差異的因素是由什么導致的以及葉性狀間關系的相關性是否會隨空間尺度的變化而呈現有規律的變化,都尚未定論。因此,系統開展不同尺度的比較研究是今后研究發展的方向。
[1]李永華,羅天祥,盧琦,等.青海省沙珠玉治沙站17種主要植物葉性因子的比較[J].生態學報,2005,25(5):994-999.
[2]Krne C H.Some often overlooked plant characteristics as determinants of plant growth:a reconsideration[J].Functional Ecology,1991,5:162-173.
[3]Wright I J,Rrich P B,Westoby M,et al.The worldwide leaf economics spectrum[J].Nature,2004,428:821-827.
[4]寶樂,劉艷紅.東靈山地區不同森林群落葉功能性狀比較[J].生態學報,2009,29(7):3692-3703.
[5]張林,羅天祥.植物葉壽命及其相關葉性狀的生態學研究進展[J].植物生態學報,2004,28(6):844-852.
[6]安慧.放牧干擾對荒漠草原植物葉性狀及其相互關系的影響[J].應用生態學報,2012,23(11):2991-2996.
[7]劉旻霞,馬建祖.甘南高寒草甸植物功能性狀和土壤因子對坡向的響應[J].應用生態學報,2012,23(12):3295-3300.
[8]宋乃平,楊新國,何秀珍,等.荒漠草原人工檸條林重建的土壤養分效應[J].水土保持通報,2012,32(4):21-26.
[9]李永華.白刺葉片性狀對人工增水的響應[D].北京:中國林業科學研究院,2010.
[10]Limm E B,Simonin K A,Bothman A G,et al.Foliar water uptake:a common water acquisition strategy for plants of the redwood forest[J].Oecologia,2009,161(1):449-459.
[11]Grime J P.Trait convergence and trait divergence in herbaceous plant communities:Mechanisms and consequences[J].Journal of Vegetation Science,2006,17:255-260.
[12]牛書麗,蔣高明,高雷明,等.內蒙古渾善達克沙地97種植物的光合生理特征[J].植物生態學報,2003,27(3):318-324.
[13]Shipley B,Almeida-Cortez J.Interspecific consistency and intraspecific variability of specific leaf area with respect to irradiance and nutrient availability[J].Ecoscience,2003,10(1):74-79.
[14]白文娟,鄭粉莉,董莉麗,等.黃土高原水蝕風蝕交錯帶不同生境植物的葉性狀[J].生態學報,2010,30(10):2529-2540.
[15]施宇,溫仲明,龔時慧.黃土丘陵區植物葉片與細根功能性狀關系及其變化[J].生態學報,2011,31(22):6805-6814.
[16]He J S,Wang X P,Flynn D F B,et al.Taxonomic,phylogenetic and environmental trade-offs between leaf productivity and persistence[J].Ecology,2009,90(10):2779-2791.
[17]鄭新軍,李嵩,李彥.準噶爾盆地荒漠植物的葉片水分吸收策略[J].植物生態學報,2011,35(9):893-905.
[18]李冰,劉左軍,趙志剛,等.海拔對鈍裂銀蓮花不同花色居群間繁殖特征及繁殖分配的影響[J].草業學報,2013,22(1):10-19.
[19]趙紅洋,李玉霖,王新源,等.科爾沁沙地52種植物葉片性狀變異特征研究[J].中國沙漠,2010,30(6):1292-1298.
[20]張林,羅天祥,鄧坤枚,等.云南松比葉面積和葉干物質含量隨冠層高度的垂直變化規律[J].北京林業大學學報,2008,30(1):40-44.
[21]張靈菲,魏斌,葛慶征,等.高寒草甸恢復演替過程中生產力與植物功能特征關系的研究[J].草業學報,2012,21(6):235-241.
[22]閆建成,梁存柱,付曉玥,等.草原與荒漠一年生植物性狀對降水變化的響應[J].草業學報,2013,22(1):68-76.
[23]李玉霖,崔建垣,蘇永中.不同沙丘生境主要植物比葉面積和葉干物質含量的比較[J].生態學報,2005,25(2):304-311.
[24]Shipley B.Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate:relationship with daily irradiance[J].Functional Ecology,2002,16(5):682-689.
[25]Reich P B,Oleksyn J,Wright I J,et al.Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes[J].Proceedings the Royal of Society(Biological sciences),2010,277:877-883.
[26]劉超,武嫻,王襄平,等.內蒙古灌木葉性狀關系及不同尺度的比較[J].北京林業大學學報,2012,34(6):23-29.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
