宮頸癌組織白介素10和干擾素γ基因表達及其啟動子區甲基化狀態研究
2014-02-10 00:50:12楊艷艷金玉蘭
中國全科醫學 2014年23期
馬 冬,楊艷艷,趙 力,金玉蘭,王 洋,李 鷗
宮頸癌患者機體內免疫微環境的變化已受到人們重視[1],其中輔助性T細胞(T helper,Th)亞群Th1/Th2的變化在惡性腫瘤免疫逃逸中十分重要[2]。組織特異性DNA甲基化譜是基因組的一個顯著特征,近年來研究表明,基因甲基化異常與宮頸癌變有關[3],因此制定宮頸癌Th1、Th2相關細胞因子DNA甲基化譜對宮頸癌的早期診斷有重要意義。本研究通過檢測宮頸癌患者宮頸組織中Th1/Th2型細胞因子干擾素γ(IFN-γ)和白介素10(IL-10)基因啟動子區甲基化狀態及轉錄活性,旨在探討DNA甲基化與基因表達的關系及其在宮頸癌中的作用。
1 資料與方法
1.1 一般資料 選取2011年3月—2012年5月在唐山市工人醫院住院部就診的宮頸癌患者43例為宮頸癌組、宮頸上皮內瘤變(CIN)患者62例為CIN組,另選取同期行全子宮切除的子宮肌瘤患者43例為對照組。其中宮頸癌組年齡46~59歲,平均(49.8±5.2)歲;病理類型均為鱗癌,根據WHO標準分級,高分化鱗癌7例,中分化鱗癌23例,低分化鱗癌13例;根據FIGO(2000年)臨床分期標準,I期27例,Ⅱ期13例,Ⅲ~Ⅳ期3例。CIN組年齡40~55歲,平均(48.6±5.1)歲;宮頸上皮內瘤變I期(CIN1)23例,宮頸上皮內瘤變Ⅱ/Ⅲ期(CIN2/3)39例。對照組年齡43~56歲,平均(50.6±5.2)歲。收集三組患者的宮頸組織標本,其中對照組標本來源于子宮肌瘤旁正常宮頸組織,CIN組和宮頸癌組標本分別來源于宮頸活檢和手術治療,均經病理檢查確診。取標本前患者均未行化療、放療或免疫治療,收集后置于-80 ℃冰箱保存。
1.2 方法
1.2.1 DNA和總RNA提取 采用飽和酚-氯仿抽提法提取三組宮頸組織DNA,Trizol法提取總RNA,1%瓊脂糖凝膠電泳鑒定DNA和總RNA完整性。
1.2.2 IL-10和IFN-γ基因啟動子區甲基化檢測 按照EZ DNA Methylation-GoldTMKit試劑盒說明書對DNA進行甲基化修飾及純化。NCBI檢索IL-10(AF418271)和IFN-γ(J00219)基因啟動子區序列,Methyl Primer Express v1.0軟件分析核對參考引物[4-5],亞硫酸氫鹽測序法(Bisulfite sequencing PCR,BSP)檢測IL-10基因啟動子區域為-454~-66,包含6個甲基化位點(-110、-185、-320、-350、-352、-373),上游引物:5′-GTTAGTAAGGAGAAGTTTTGGG-3′,下游引物:5′-CATTCATTAAAAAACCACAAT-3′,擴增片段389 bp。BSP反應體系(50 μl):PCR Master Mix(美國Promega)47 μl,上、下游引物(10 μM)各0.5 μl,DNA模板2 μl。反應條件:95 ℃ 3 min,94 ℃ 30 s、55 ℃ 30 s、72 ℃ 30 s(30個循環),72 ℃ 5 min。甲基化特異性PCR(MSP)檢測IFN-γ基因啟動子區甲基化,甲基化引物為:上游5′-AAGAGTTAATATTTTATTAGGGCGA-3′,下游5′-TAAACTCCTTAAATCCTTTAACGAT-3′;非甲基化引物為:上游5′-TGAAGAGTTAATATTTTATTAGGGTGA-3′,下游5′-TAAACTCCTTAAATCCTTTAACAAT-3′,PCR產物片段156 bp。MSP反應體系(20 μl):PCR Master Mix 18 μl,上、下游引物(10 μM)各0.5 μl,DNA模板1 μl。反應條件:95 ℃ 3 min,94 ℃ 45 s、50 ℃ 60 s、72 ℃ 45 s(35個循環),72 ℃ 5 min。PCR產物經1.5%瓊脂糖凝膠電泳后觀察條帶。甲基化結果判斷:僅出現甲基化引物擴增154 bp片段產物判別為甲基化,僅出現非甲基化引物擴增156 bp片段產物判別為非甲基化,甲基化引物及非甲基化引物均擴增出條帶為部分甲基化。
1.2.3 宮頸組織中IL-10和IFN-γ基因mRNA的表達 采用實時熒光定量PCR(RT-PCR)檢測三組患者宮頸組織中IL-10和IFN-γ基因的mRNA表達水平[6]。RT-PCR反應:按M-MLV反轉錄酶系(美國GIBCO)說明書操作,加入100 ng隨機引物和0.5~5.0 μg總RNA及緩沖劑(buffer)混勻后,37 ℃孵育1 h,70 ℃滅活15 min,cDNA保存于-20 ℃備用。20 μl PCR反應體系:12 μl buffer,上、下游引物各1 μl,cDNA產物2 μl,Taq酶1 μl,加超純水至20 μl混合。反應程序:93 ℃變性1 min,55 ℃退火1 min,72 ℃延伸1 min,共進行30個循環。
1.2.4 轉錄因子結合位點序列分析 轉錄因子的結合DNA序列位點的分析采用TFSEARCH軟件、promoter scan軟件和在線AliBaba 2.1分析工具。把目的序列粘貼在在線軟件提供的粘貼框,按執行按鈕即可,具體網址如下:http://mbs.cbrc.jp/research/db/TFSEARCH.html;http://www-bimas.cit.nih.gov/molbio/proscan/index.html;http://www.gene-regulation.com/pub/programs/alibaba2/index.html。

2 結果
2.1 三組宮頸組織IL-10和IFN-γ mRNA的表達 三組IL-10和IFN-γ mRNA水平比較,差異均有統計學意義(P<0.05),其中宮頸癌組和CIN組IL-10 mRNA水平均高于對照組,IFN-γ mRNA水平均低于對照組,差異有統計學意義(P<0.01,見表1)。
2.2 三組IL-10和IFN-γ基因啟動子區甲基化分析 三組IL-10基因啟動子區-320、-350、-352、-373位點甲基化率比較,差異均無統計學意義(P>0.05);三組IL-10基因啟動子區-110、-185位點甲基化率比較,差異均有統計學意義(P<0.05),其中宮頸癌組和CIN組IL-10基因啟動子區-110位點甲基化率均高于對照組,宮頸癌組IL-10基因啟動子區-185位點甲基化率低于對照組,差異均有統計學意義(P<0.05,見表2)。不同組織提取DNA的IFN-γ基因啟動子甲基化狀態見圖1。宮頸癌組IFN-γ基因啟動子甲基化率為34.9%(15/43),CIN組IFN-γ基因啟動子甲基化率為14.5%(9/62),對照組IFN-γ基因啟動子甲基化率為4.6%(2/43)。三組IFN-γ基因啟動子甲基化率比較,差異有統計學意義(χ2=14.250,P=0.001),其中宮頸癌組IFN-γ基因啟動子甲基化率高于CIN組,CIN組高于對照組,差異均有統計學意義(χ2=5.970,P=0.015;χ2=12.390,P<0.001)。
Table1 Comparison of the level of IL-10 and IFN-γ mRNA among the 3 groups
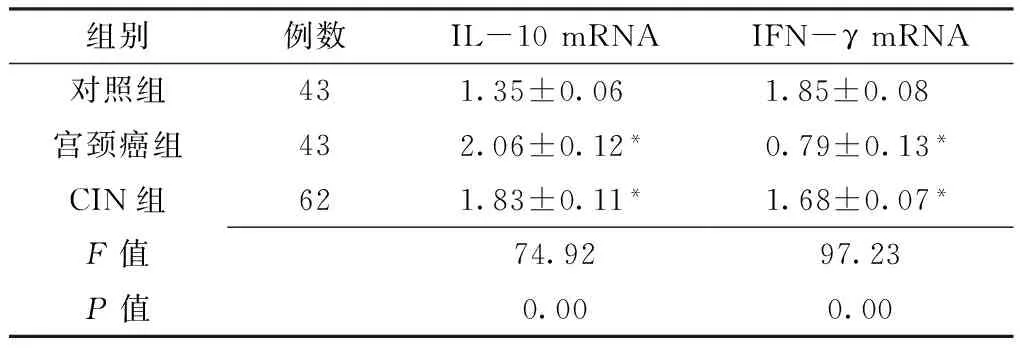
組別例數IL-10mRNAIFN-γmRNA對照組431.35±0.06 1.85±0.08 宮頸癌組432.06±0.12*0.79±0.13*CIN組621.83±0.11*1.68±0.07*F值74.9297.23P值0.000.00
注:與對照組比較,*P<0.01;IL-10=白介素10,IFN-γ=干擾素γ
表2 三組IL-10基因啟動子區位點甲基化率比較〔n(%)〕
Table2 Comparison of methylation rates of IL-10 gene promoter among the 3 groups
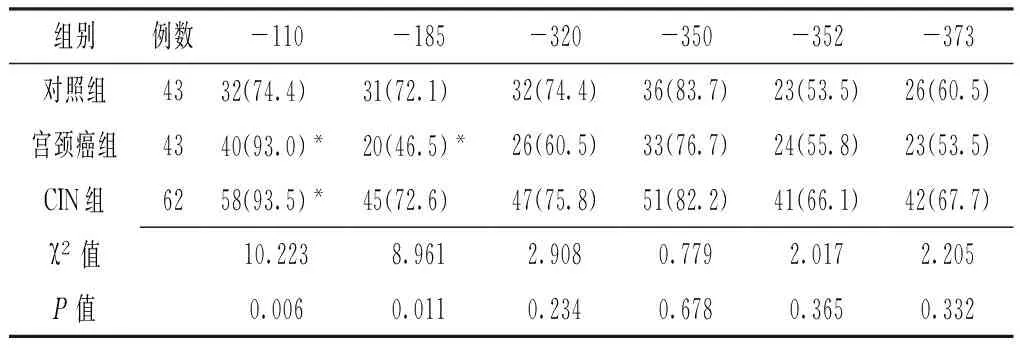
組別例數-110-185-320-350-352-373對照組4332(74.4) 31(72.1) 32(74.4)36(83.7)23(53.5)26(60.5)宮頸癌組4340(93.0)*20(46.5)*26(60.5)33(76.7)24(55.8)23(53.5)CIN組6258(93.5)*45(72.6) 47(75.8)51(82.2)41(66.1)42(67.7)χ2值10.2238.9612.9080.7792.0172.205P值0.0060.0110.2340.6780.3650.332
注:與對照組比較,*P<0.05
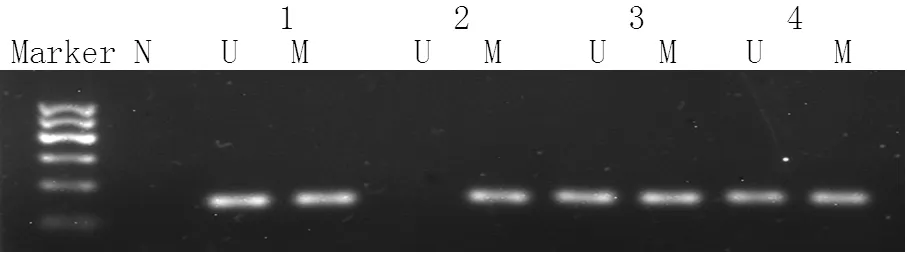
注:N=空白對照,U=非甲基化,M=甲基化,1=對照組,2=宮頸癌組,3=CIN1,4=CIN2/3
圖1 MSP檢測三組IFN-γ基因啟動子甲基化狀態
Figure1 Methylation of IFN-γ gene promoter by MSP among the 3 groups
2.3 甲基化對IL-10基因轉錄活性的影響 通過在線軟件TFSEARCH發現IL-10基因啟動子區可能包含ER、HSF、HSF或AP-4、CCAAT-box、STAT3或Ets-1共5個轉錄因子結合域(見圖2)。

圖2 IL-10基因啟動子區與轉錄因子結合位點預測
Figure2 Prediction of IL-10 gene promoter and transcription factor binding sites
3 討論
機體的抗腫瘤作用是以Th1介導的細胞免疫為主,而Th1向Th2漂移,將造成免疫抑制狀態,機體的抗腫瘤免疫將受到嚴重干擾[7]。IFN-γ是Th1分泌的主要細胞因子,且可誘導前體T細胞向Th1分泌,是抗腫瘤免疫中較早出現的細胞因子;IL-10是一種有助于下調免疫反應的Th2型細胞因子,具有多種抑制抗腫瘤免疫應答的作用。IL-10升高,不利于抗腫瘤免疫。本研究結果顯示,宮頸癌患者IL-10表達水平明顯增強,IFN-γ表達水平明顯降低,呈現出典型的Th2漂移現象,這可能是宮頸癌患者免疫監視功能低下的主要原因之一。
DNA甲基化主要在轉錄水平抑制細胞因子基因的表達[8],CpG島甲基化與其所在基因的表達呈反比,即CpG島甲基化程度越高,基因表達水平越低,反之則越高。本研究對IL-10和IFN-γ基因的甲基化狀態進行分析,結果表明,宮頸癌患者IL-10基因啟動子區-185位點甲基化率較低,而IFN-γ甲基化率異常增高,與Viana等[6]研究牙髓炎及Son等[4]研究乳腺癌相關報道一致,說明DNA甲基化導致基因表達異常,造成免疫功能低下,失去對癌細胞的免疫監視和清除。
DNA甲基化引起基因轉錄抑制的機制主要有3種[9],即直接作用模式(結合位點DNA甲基化導致相應轉錄因子不能識別與結合)、間接作用模式(序列特異性的甲基化DNA結合蛋白與啟動子區甲基化CpG島結合,募集一些蛋白,形成轉錄抑制復合物,從而阻止轉錄因子與啟動子區的靶序列結合,進而影響基因轉錄)和改變染色質結構模式(DNA的甲基化致使染色質結構改變,抑制基因表達)。前期研究發現,宮頸癌組織IL-10基因CpG島的甲基化率較低[10],且Szalmás等[11]在鱗狀上皮細胞中發現IL-10基因的轉錄活性與轉錄因子STAT3、Sp1、Ets-1和CCAAT-box有關。因此,本研究采用BSP進一步明確精確位點的甲基化對IL-10基因轉錄活性的可能影響。通過DNA序列的轉錄因子結合位點預測,發現IL-10基因啟動子區可能包含ER、HSF、HSF或AP-4、CCAAT-box、STAT3或Ets-1共5個轉錄因子結合域,其中-110、-185位點的甲基化率在宮頸癌組和對照組中具有顯著差異,影響CCAAT-box、STAT3和Ets-1與之結合。有趣的是兩位點甲基化率水平高低在宮頸癌組和對照組中正好相反,-110位點的甲基化上調可能影響轉錄因子STAT3或Ets-1相關功能,而-185位點甲基化下調可能影響轉錄因子CCAAT-box的結合活性增強,進而影響IL-10基因在宮頸癌中的表達水平升高。
綜上所述,IL-10基因啟動子區域-110、-185位點甲基化狀態使其在宮頸組織中的分泌表達增高,IFN-γ甲基化率異常增高,導致其在宮頸組織中的分泌表達減少,宮頸癌患者處于Th2極化狀態,與宮頸癌的發生有關。表觀遺傳對細胞因子基因表達的調控除DNA甲基化外,還包括組蛋白乙酰化[12]和染色質結構變化[13]等方式,但其在宮頸癌發生中的可能作用將是筆者下一步研究的重點,這將為診斷和免疫治療宮頸癌的研究提供科學參考。
1 馬翠,張為遠.宮頸癌與免疫功能狀態[J].中國婦幼保健,2009,24(25):3608-3611.
2 陳遠崇,陳竹雨.結直腸癌患者血清Th1/Th2細胞因子水平變化及其臨床意義研究[J].中國全科醫學,2011,14(12):4136-4138.
3 Szalmás A,Kónya J.Epigenetic alterations in cervical carcinogenesis[J].Semin Cancer Biol,2009,19(3):144-152.
4 Son KS,Kang HS,Kim SJ,et al.Hypomethylation of the interleukin-10 gene in breast cancer tissues[J].Breast,2010,19(6):484-488.
5 Bonilla-Henao V,Martínez R,Sobrino F,et al.Different signaling pathways inhibit DNA methylation activity and up-regulate IFN-gamma in human lymphocytes[J].J Leukoc Biol,2005, 78(6):1339-1346.
6 Viana MB,Cardoso FP,Diniz MG,et al.Methylation pattern of IFN-γ and IL-10 genes in periodontal tissues[J].Immunobiology,2011,216(8):936-941.
7 Hedrich CM,Bream JH.Cell type-specific regulation of IL-10 expression in inflammation and disease[J].Immunol Res,2010,47(1/3):185-206.
8 Wilson AG.Epigenetic regulation of gene expression in the inflammatory response and relevance to common diseases[J].J Periodontol,2008,79(8 Suppl):1514-1519.
9 Berger SL.The complex language of chromatin regulation during transcription[J].Nature,2007, 447(7143):407-412.
10 馬冬,蔚巖,王艷,等.IL-10基因啟動子區CpG島甲基化與宮頸癌的相關性研究[J].中國婦幼保健,2013,28(12):1956-1959.
11 Szalmás A,Bánáti F,Koroknai A,et al.Lineage-specific silencing of human IL-10 gene expression by promoter methylation in cervical cancer cells[J].Eur J Cancer,2008,44(7):1030-1038.
12 Wang Z,Zang C,Rosenfeld JA,et al.Combinatorial patterns of histone acetylations and methylations in the human genome[J].Nat Genet,2008,40(7):897-903.
13 Li B,Carey M,Workman JL.The role of chromatin during transcription[J].Cell,2007,128(4): 707-719.
