搖晃蛋白引起神經元分枝并促進樹突發育①
2014-03-18 11:41:16段明慧陳樹林趙善廷西北農林科技大學動物醫學院楊陵712100
中國免疫學雜志 2014年2期
關鍵詞:小鼠
盧 習 范 力 邵 虹 段明慧 陳樹林 趙善廷 (西北農林科技大學動物醫學院,楊陵 712100)
哺乳動物的大腦皮質是一個高度有序的復雜結構,從軟腦膜(Pial mater)到白質(White matter)是由不同類型的細胞排列而成的六層結構,每一層的神經元具有相似的形態結構和功能特性,并且在同一時間起源于同一地點[1]。在哺乳動物大腦皮質發育時期,有絲分裂后的神經元從它們的發源地沿著放射狀膠質細胞的突起向軟腦膜遷移,最后會以“由內而外”(Inside out)的方式形成高度有序排列的六層結構[2],即較早出生的神經元最終定位于皮層深處,而晚出生神經元最終占據著淺層區域。神經元遷移缺陷會導致許多神經元疾病,例如無腦回(Lissencephaly)、癲癇(Epilepsy)、智力遲鈍(Mental retardation)和嚴重的學習障礙(Iearning disabilities)[3,4]。神經元遷移依靠不同信號通路的精確調控,其中最具特色的是搖晃蛋白(Reelin)信號通路。
在大腦皮質發育過程中,位于邊緣帶(Marginal zone,MZ)的Cajal-Retzius 細胞(CR cells)分泌的一種細胞外基質蛋白,稱為搖晃蛋白(Reelin)。在大腦皮質發育過程中,搖晃蛋白控制著神經元遷移和大腦皮質的形成過程。在缺失搖晃蛋白的突變體小鼠(搖晃小鼠,Reeler mouse)的大腦皮質、海馬和小腦中神經元的遷移嚴重受阻,最終導致層結構的反轉[5]。
以前研究發現,除了大腦皮質的層結構反轉外,純合子搖晃小鼠海馬中樹突發育異常,排列紊亂[6,7]。樹突分枝朝向紊亂的程度與細胞層異位的程度相一致,表明發育樹突的朝向受位置信號的影響。雜合子搖晃小鼠在沒有出現細胞層異位的情況下仍表現出樹突長度變短,樹突棘(Dendritic spine)密度減小[8],這表明搖晃蛋白的缺失本身可能影響樹突發育和神經元成熟。
本研究通過子宮內電擊轉染的方法,標記遷移中的神經元,觀察到在接近含搖晃蛋白的MZ 附近,遷移神經元開始出現分枝,表現出與在遷移中完全不一樣的表型。通過分析發現搖晃蛋白引起神經元分枝,因此神經元的發育與搖晃蛋白之間有著密切的關系。
1 材料與方法
1.1 實驗動物 本研究工作中使用的動物為孕期E15.5 的野生型C57BL/6J 小鼠。成年C57BL/6J小鼠(2 月齡)購自于西安交通大學實驗動物中心。小鼠首先放置于實驗室鼠房[室溫(22±1)℃,相對濕度50%~60%]適應7 d。按照雌∶雄=2∶1 的比例進行合籠,次日上午檢查陰栓,若見陰栓,當天中午記為E0.5(Embryonic day 0.5)。小鼠出生的當天記為P0(Postnatal day 0)。
1.2 子宮內電擊轉染 子宮內電擊轉染實驗方法參照Saito 等[9]的文獻進行,并做適當修改。
取E15.5 孕鼠,用0.7%戊巴比妥鈉溶液進行腹腔注射(70 mg/kg),麻醉后將其腹部向上置平板上,用碘伏和酒精棉反復交替消毒腹部皮膚,用中間開口的無菌手術墊單覆蓋腹部,沿腹中線剪開皮膚和肌肉,將一側子宮輕輕拉出,找到腦部人字縫,判斷側腦室的位置,以左手固定胎鼠,用玻璃微注射針(圖1A、C 中箭形所示)將pCAG-GFP(Plasmid 11150,Addgene,Cambridge,MA,USA)1.5 μl/μg 注射到鼠胚一側側腦室,其中含0.01% 固綠(Fast green)作為指示劑,用板狀電極(圖1B、D 黑色所示)輕輕夾住胎腦,按照如下參數進行電脈沖刺激:電壓45 V,脈沖時間50 ms,脈沖間隔950 ms,電擊次數5 次。在此過程中,持續往子宮上滴加預熱到37℃的無菌生理鹽水。轉染完畢,將子宮輕輕放回腹腔中,以預熱生理鹽水灌滿腹腔后將肌肉和皮膚分別縫合。手術完畢,將母鼠放回籠中,保溫至清醒,分別飼養至轉染后3 d 或5 d 后,打開母鼠腹腔,取出胚胎,若出生則取新生小鼠灌注取材。
1.3 熒光免疫組織化學染色 取出的胚胎大腦在4%多聚甲醛中固定3 d 后,用4%瓊脂包埋,振蕩切片機切片,厚度50 μm,于pH7.4 的0.1 mol/L 磷酸緩沖液(Phosphate buffer,PB)中保存,隨后進行免疫熒光染色。
熒光免疫組織化學染色采用“三步法”在24 孔培養板中進行漂片染色,孵育及漂洗過程均在低速搖床上進行。使用的一抗包括:rabbit anti-GFP(1∶1 000;A-11122,Invitrogen,Karlsruhe,Germany),mouse anti-Reelin G10 (1 ∶1 000;MAB5364,Millipore,Billerica,MA,USA);二抗包括goat anti rabbit-Alexa Fluor 488,goat anti mouse-Alexa Fluor 568。用PI(Propidium iodide,2.5 mg/ml;P3566,Invitrogen,Karlsruhe,Germany)或DAPI(4',6-diamidino-2-phenylindole,1 ∶10 000;236276,Roche Molecular Biochemicals,Mannheim,Germany)室溫下復染15 min 以標記細胞核。腦片用防熒光淬滅劑(Fluorescence Mounting Medium,S3023,DAKO,Copenhagen,Denmark)封片,然后在Zeiss LSM 510激光共聚焦顯微鏡下觀察結果并拍照。
1.4 熒光顯微觀察與激光共聚焦顯微拍照 在Nikon 熒光體視顯微鏡下觀察取出的完整的小鼠腦或胚胎腦,確定轉染效果及GFP 表達的腦區。在Zeiss LSM 510 激光共聚焦顯微鏡下主要觀察被GFP 標記上的細胞的形態以及分布的部位。
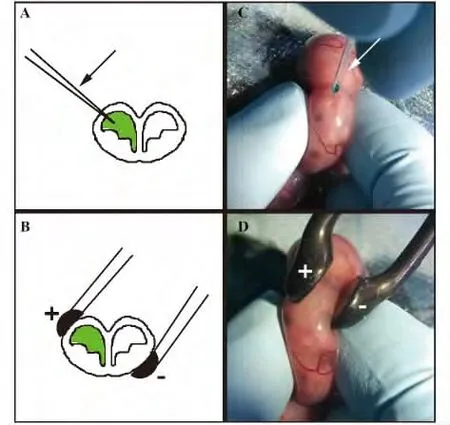
圖1 子宮內電擊轉染示意圖Fig.1 Illustration of in utero electroporation procedure
1.5 細胞形態分析 使用Zeiss LSM510 激光共聚焦顯微鏡(×60,1.4 倍物鏡孔徑)對切片進行z 軸連續掃描拍照,然后通過Zeiss ZEN 2010 軟件依據“最大像素密度”將每一組照片進行疊加成一個平面。通過Neuromantic 軟件描繪出單個神經元的形態和突起。細胞的每根突起的長度可以通過Image J 軟件測量出來。為了分析突起的形態特征,本實驗分析了以下兩個參數:總的分枝長度(Total branch length,TBL)及分枝點(Bifurcation)數目。將從頂突起上伸出的長度大于2.5 μm 的GFP 陽性突起定義為初級分枝,從頂突起上伸出的長度小于2.5 μm 的不穩定絲狀偽足不計入分枝內。將從神經元胞體和頂突起間的收縮部位到第一個分枝點的距離定義為剩余頂突起的長度(Length of leading process)。
1.6 統計學分析 所有數據用統計軟件SPSS17.0進行統計學分析,組間的比較進行單因素方差分析(ANOVA)和Dennet t 檢驗。所有結果以±s 表示,P <0.05 記為數據有顯著性差異。
2 結果
2.1 搖晃蛋白局限性分布于發育中大腦皮質的MZ 用G10 抗體做免疫組織化學染色可以顯示搖晃蛋白在胚胎大腦皮質中的分布位置。G10 抗體是一種單克隆IgG 抗體,可以識別搖晃蛋白N 末端序列上的第164~496 位氨基酸。免疫組化結果顯示,搖晃蛋白主要分布在大腦皮質胞核較為稀疏的MZ區,呈現出一種較好的彌散性的分布模式(圖2 A,A')。在放大圖中可以看到,MZ 中許多細胞顯示出強烈的搖晃蛋白陽性,呈現出典型的CR 細胞特征,水平朝向,帶有粗的突起的雙極形態,胞體為卵圓形或梭形(箭頭)。在MZ 中的CR 細胞之間,檢測到較強的、呈彌散點狀分布的搖晃蛋白分布(圖2 B,B')。
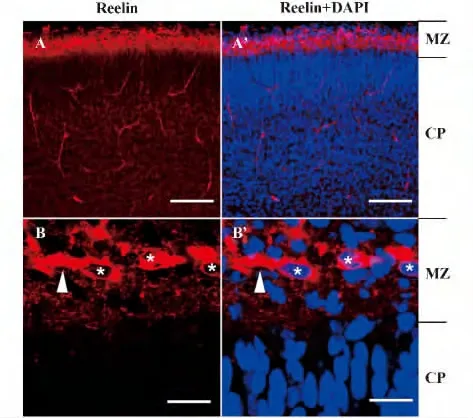
圖2 胚胎期大腦皮質中搖晃蛋白的分布Fig.2 Restricted expression of Reelin in embryonic cerebral cortex
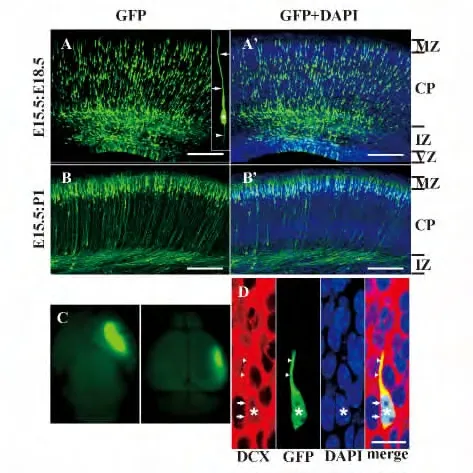
圖3 通過子宮內電擊轉染技術用GFP 標記胚胎大腦中神經元Fig.3 Neurons were labeled by GFP in embryonic brain transfected using in utero electroporation
2.2 利用子宮內電擊轉染技術成功標記遷移神經元 利用子宮內電擊轉染技術,在小鼠胚胎的不同發育時期,將帶有CAG 啟動子的綠色熒光蛋白(Green fluorescent protein,GFP)表達質粒(pCAGGFP)注射入小鼠胚胎側腦室,在一定的電脈沖刺激下,外源質粒進入胚胎側腦室的室帶中的神經前體細胞中,在CAG 啟動子的驅動下,GFP 在轉染中的細胞中表達。
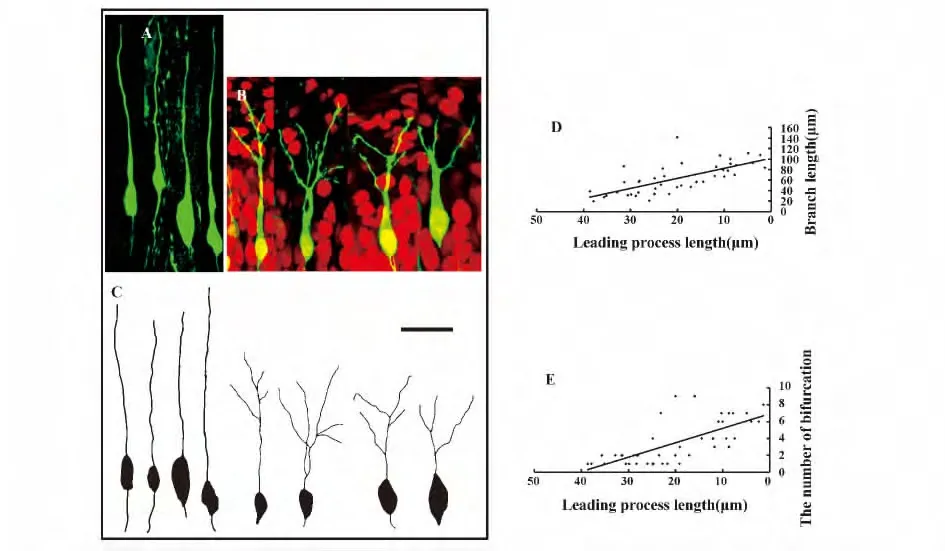
圖4 搖晃蛋白促進神經元的分枝和樹突的發育Fig.4 Reelin promoted branching of neurons and dendritic development
在不同的發育階段,將剝離頭蓋骨的小鼠胚胎完整腦組織或是出生后小鼠腦組織放置于Nikon 熒光體視顯微鏡下,在質粒濃度較高及恰當的轉染條件下可以直接觀察到強烈的GFP 陽性表達(圖3C)。
在E15.5 d 進行子宮內電擊轉染,3 d 后E18.5 d 固定的小鼠胚胎大腦中,可以觀察到在發育中的大腦皮質的皮質板(Cortical plate,CP)、中間帶(Intermediate zone,IZ)和室帶(Ventricular zone,VZ)/亞室帶(Subventricular zone,SVZ)中均有大量的GFP 陽性的細胞被成功標記(圖3A,A')。在CP 中的GFP 陽性的細胞表現出遷移神經元典型的雙極形態特征,具有卵圓形的胞體、一條長、粗的頂突起伸向軟腦膜和一條短、細的尾突起伸向側腦室(圖3 A 插圖)。一些GFP 陽性的細胞已經到達CP 的淺層區域,并且它們的頂突起非常接近或已經侵入MZ 中(圖3A,A')。轉染5 d 后(P1)檢測結果,發現大部分GFP 陽性細胞聚集于MZ 的邊緣,并形成一條致密的帶。它們向MZ 中伸出一些小的分枝(圖3B,B')。GFP 標記出的細胞的形態暗示出它們是神經元。
為了證實這些被GFP 標記的細胞確實是神經元我們進行了雙皮質素(Doublecortin,DCX)免疫組化染色。DCX 是一種微管相關蛋白,被普遍認為是有絲分裂后遷移神經元的標記物[10,11],即能顯示出該神經元未成熟的特征。實驗結果表明,在電擊轉染后第3 天檢測結果,發現在大腦皮質的CP 中大部分GFP 陽性細胞都是DCX 陽性的(圖3D),表明在E15.5 用GFP 質粒轉染,且在3 d 或是更長時間后在大腦皮質中標記出的細胞的確是神經元。
2.3 搖晃蛋白引起神經元產生分枝促進樹突發育 已有研究表明,當遷移神經元的頂突起接觸到CP-MZ 的邊界線時,遷移神經元便將其遷移方式由依賴膠質細胞的整體運動式遷移轉變成胞體位移式遷移的方式,以完成最終的遷移過程。為了研究搖晃蛋白在這一過程中的作用,將遷移神經元按照其所處位置分成兩類:(1)在皮質板中遷移的神經元,神經元的胞體和頂突起都位于大腦皮質板中(圖4A);(2)已接觸到搖晃蛋白的遷移神經元,神經元的胞體接近或接觸到CP-MZ 交界線且其頂突起已經接觸到搖晃蛋白豐富的MZ 區(圖4B)。為了方便統計,利用Neuromantic 軟件對每組中典型的神經元進行示蹤,并畫出其模式圖(圖4C)。
研究發現,頂突起接觸到搖晃蛋白豐富的MZ區域的神經元和頂突起仍在大腦皮質板中的神經元在突起的分枝程度上有明顯的差異。在大腦皮質板中仍然進行整體運動式遷移的神經元的頂突起朝向MZ 區,并沒有出現分枝(圖4A),而當遷移神經元的頂突起剛接觸到搖晃蛋白豐富的MZ 區域時,頂突起的末端開始出現分枝,并且隨著遷移神經元繼續向上遷移,胞體離搖晃蛋白豐富的MZ 區域越近,頂突起的長度越來越短,此時,頂突起上的分枝數目越來越多(圖4 B、D)。遷移神經元的胞體最終會停留在大腦皮質板的最上端而不侵入搖晃蛋白豐富的MZ 區域中,此時,神經元的頂突起完全消失,出現了大量的、繁茂的樹枝狀的突起。
為了對比位于大腦皮質板中的神經元和頂突起接觸到MZ 區域的神經元的形態特征,測量神經元頂突起的長度和其對應的突起的長度并且計數每一個神經元的頂樹突上的分枝點數。大腦皮質板中遷移神經元的頂突起的長度為(65.73±10.88)μm。隨著神經元向上遷移,剩余頂突起的長度越來越短,神經元頂樹突上的分枝的長度越來越長且分枝點越來越多。定量分析結果顯示,神經元的剩余頂突起的長度與總的分枝長度呈極顯著負相關(n=42,r=-0.700,P=2.44E-7;圖4D),且與分枝點的數目呈極顯著負相關(n=39,r=-0.701,P=2.28E-7;圖4E)。
3 討論
3.1 搖晃蛋白促進神經元分枝在神經元遷移中的作用 利用胚胎大腦皮質腦片培養方法來研究神經元遷移模型[12],在頂突起(平均長度為30~50μm)接觸到MZ 后,胞體運動的方式由跳躍性轉向連續性,并且胞體遷移的平均速度由35 μm/h 增加到60 μm/h。遷移神經元的頂突起一旦接觸到搖晃蛋白豐富的MZ,頂突起便開始產生初生的分枝,神經元的遷移方式也會轉變成胞體位移式遷移的形式,這種遷移方式是不依賴于放射狀膠質細胞的[12]。在這個過程中,頂突起越來越短,在搖晃蛋白豐富的MZ 中的分枝越來越多,遷移神經元的胞體也得以快速向上繼續遷移,這種遷移的動力可能來自于搖晃蛋白引起的神經元頂突起的快速的形態學改變而產生的向上的牽引拉力。一方面,這些分枝可以錨定在MZ 中,提供這種類似“引體向上”式的牽引力,為神經元最后通過胞體位移式遷移到達最終位置提供動力;另一方面,當遷移神經元的胞體到達被細胞外基質中搖晃蛋白包圍著的初級分枝點時,分枝點阻止了神經元細胞核在頂突起中向上移動,因此神經元停止遷移。搖晃蛋白引起的頂突起的分枝便起到了終止信號的作用。
3.2 搖晃蛋白對神經元的樹突發育的影響 在本研究中,使用子宮內電擊轉染方法成功將GFP 標記并追蹤正在遷移的神經元,這就使得在體觀察到神經元在不同的發育階段時的形態特征成為可能。結合搖晃蛋白免疫組化染色,神經元在搖晃蛋白陽性的MZ 區出現分枝,可以推論搖晃蛋白促進神經元產生分枝。
之前的研究表明,純合子搖晃小鼠中除了皮層反轉外,另外一個顯著的特征是頂突起和樹突發育受損,朝向紊亂,放射狀膠質細胞的突起發育異常[7]。雜合子搖晃小鼠中沒有出現細胞層紊亂,但是樹突發育仍受到影響[9],說明搖晃蛋白本身可能影響樹突的發育。將重組的搖晃蛋白加到培養基中后可以促進純合子搖晃小鼠海馬神經元的樹突的生長[7]。以前有報道表明搖晃蛋白在大腦皮質和海馬神經元的樹突棘周圍的細胞外基質中聚集[13],在雜合子搖晃小鼠中細胞層并沒有出現異位的情況下小棘的密度仍然減少了,說明搖晃蛋白可以調節生后小鼠樹突的發育[14]。另外,搖晃蛋白可以促進海馬神經元樹突棘的發育,使其變得過度肥大[15]。綜上所述,表明搖晃蛋白可以直接調節神經元的分枝和樹突的發育。
[1]Rakic P.Specification of cerebral cortical areas[J].Science,1988,241(4862):170-176.
[2]Nadarajah B,Parnavelas JG.Modes of neuronal migration in the developing cerebral cortex[J].Nat Rev Neurosci,2002,3(6):423-432.
[3]Ayala R,Shu T,Tsai LH.Trekking across the brain:the journey of neuronal migration[J].Cell,2007,128(1):29-43.
[4]Lambert de Rouvroit C,Goffinet A M.Neuronal migration[J].Mech Dev,2001,105(1-2):47-56.
[5]Curran T,D’Arcangelo G.Role of reelin in the control of brain development[J].Brain Res Brain Res Rev,1998,26(2-3):285-294.
[6]Tabata H,Nakajima K.Neurons tend to stop migration and differentiate along the cortical internal plexiform zones in the Reelin signal-deficient mice[J].J Neurosci Res,2002,69(6):723-730.
[7]Niu S,Renfro A,Quattrocchi CC,et al.Reelin promotes hippocampal dendrite development through the VLDLR/ApoER2-Dab1 pathway[J].Neuron,2004,41(1):71-84.
[8]Liu WS,Pesold C,Rodriguez MA,et al.Down-regulation of dendritic spine and glutamic acid decarboxylase 67 expressions in the reelin haploinsufficient heterozygous reeler mouse[J].Proc Natl Acad Sci USA,2001,98(6):3477-3482.
[9]Saito T,Nakatsuji N.Efficient gene transfer into the embryonic mouse brain using in vivo electroporation[J].Dev Biol,2001,240(1):237-246.
[10]Francis F,Koulakoff A,Boucher D,et al.Double cortin is a developmentally regulated,microtubule-associated protein expressed in migrating and differentiating neurons[J].Neuron,1999,23(2):247-256.
[11]Gleeson JG,Lin PT,Flanagan LA,et al.Double cortin is a microtubule-associated protein and is expressed widely by migrating neurons[J].Neuron,1999,23(2):257-271.
[12]Nadarajah B,Brunstrom JE,Grutzendler J,et al.Two modes of radial migration in early development of the cerebral cortex[J].Nat Neurosci,2001,4(2):143-150.
[13]Rodriguez MA,Pesold C,Liu WS,et al.Colocalization of integrin receptors and reelin in dendritic spine postsynaptic densities of adult nonhuman primate cortex[J].Proc Natl Acad Sci USA,2000,97(7):3550-3555.
[14]Chameau P,Inta D,Vitalis T,et al.The N-terminal region of reelin regulates postnatal dendritic maturation of cortical pyramidal neurons[J].Proc Natl Acad Sci USA,2009,106(17):7227-7232.
[15]Niu S,Yabut O,D’Arcangelo G.The Reelin signaling pathway promotes dendritic spine development in hippocampal neurons[J].J Neurosci,2008,28(41):10339-10348.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34