寧波沿海陸源排污口擬桿菌(Bacteroidetes)分布的特點*
2014-03-19 10:18:12王中華徐茂琴張迪駿何偉娜陳麗萍童茜茜張春丹蘇秀榕
海洋與湖沼 2014年5期
關鍵詞:檢測
王中華 徐茂琴 謝 利 張迪駿 周 君 何偉娜 陳麗萍 童茜茜 張春丹 蘇秀榕
(1. 寧波大學海洋學院 寧波 315211; 2. 寧波城市職業技術學院 寧波 315100;3. 國家海洋局北海環境監測中心 青島 266061)
高通量454焦磷酸測序, 是以DNA擴增的乳膠系統和皮升級(picolilter, 10–12liter)焦磷酸為基礎的測序方法(Margulieset al, 2005), 該技術被廣泛應用于土壤(Roeschet al, 2007)、海洋(Paleniket al, 2009)等環境微生物生態學研究中。擬桿菌門(Bacteroidete)是腸道中數量最大的革蘭氏陰性細菌, 在人和鼠的盲腸內容物中擬桿菌屬占總細菌的20%—40% (Woodet al, 1998)。雖然擬桿菌可以從腸道中分離得到, 但是部分擬桿菌是可以致病的, 而人們對與擬桿菌的認識也是從致病性開始(Wexler, 2012), 作為條件致病菌的擬桿菌, 當其正常的微生態平衡被打破時可引發內源性感染。隨著時間的發展和研究的深入, 人們逐漸發現擬桿菌和其它菌群與宿主之間微妙而又復雜的關系, 對宿主健康起著重要的作用(吳彥彬等, 2007),而在另一方面, 這類長期被環境檢測和公共衛生機構所忽略的微生物逐漸為指示水體所受的糞源污染提供了一條新的途徑(Wadeet al, 2006)。而擬桿菌門還包括另外兩綱, 分別為黃桿菌綱(Flavobacteria)和鞘脂桿菌綱(Sphingobacteria)。黃桿菌綱主要存在于水生環境中, 也會在食物中存在, 多數黃桿菌綱細菌對人無害, 但腦膜膿毒性金黃桿菌(Chryseobacterium meningosepticum)可引起新生兒腦膜炎(Linet al,2010)。而鞘脂桿菌綱在海洋細菌中占有較大比例, 可以降解纖維素(Liet al, 2011)。
本文選取寧波沿海10個主要的陸源排污口, 分別對排污口處和距排污口50m以外的20個站位的水樣進行分析。在454焦磷酸測序基礎上, 首先對各個站點總體情況進行統計分析, 再根據不同采樣月份各站位擬桿菌門, 包括擬桿菌綱、黃桿菌綱和鞘脂桿菌綱檢出頻次的統計, 分析擬桿菌門的分布概況。
1 材料與方法
1.1 樣品采集
選擇寧波市10個陸源排污口, 分別為象山爵溪東塘排污口(S1)、象山水桶岱垃圾場滲透水排污口(S2)、象山石浦水產加工園區排污口(S3)、象山西周工業園區綜合排污口(S4)、象山墻頭綜合排污口(S5)、北侖三山排污口(S6)、寧海西店崔家綜合排污口(S7)、奉化市下陳排污口(S8)、寧海顏公河入海排污口(S9)和余姚黃家埠排污口(S10)(陳麗萍等, 2013)。將5 L的桶消毒后采集各占站點的水樣。樣品收集時間分別為2011年3月、5月、8月、10月。10個站點分別標記為S1—S10, 采樣月份分別標記為M3、M5、M8、M10, 排污口和排污口50m以外水樣分別標記為–0和–50。樣品采集完畢后, 立即運回實驗室, 用0.2 μm微濾膜對污水進行過濾之后進行DNA提取。
1.2 總DNA提取及擴增
利用水體細菌基因組DNA提取試劑盒(Omega Bio-Tek), 按照試劑盒說明書提取基因組DNA, 測定OD260/OD280在1.8—2.0, OD260/OD230高于1.7。利用Primer Premier 5.0設計擴增引物, 其序列如下: P1:5’-CCA TCT CAT CCC TGC GTG TCT CCG ACT CAG-3’; P2: 5’-CCT ATC CCC TGT GTG CCT TGG CAG TCT CAG-3’。反應體系如下: 10×buffer 2.5μL、25 mmol/L MgCl22.5μL、2.5 mmol/L dNTPs 2.0μL、10μmol/L 引物各 1.0μL, 1.0 U rTaq酶、模板DNA 1.0μL, 無菌超純水補足至25μL。PCR擴增條件為:95°C預變性4 min; 94°C變性45 s, 56°C退火45 s,72°C延伸1 min, 30個循環; 72°C延伸 10 min。取23μL PCR產物, 加入5μL 10×loading buffer混勻,1%瓊脂糖凝膠電泳, –20°C冰箱保存備用(周君等,2012)。
1.3 測序與分析
將PCR擴增產物經電泳檢測合格后, 利用新一代高通量測序平臺454 GS FLX Titanium對擴增產物進行測序。利用SPSS 19.0對數據進行統計分析。
2 結果與討論
2.1 454測序數據總體分析
通過454高通量測序平臺對20個站點4個檢測月份80個樣品進行鑒定分析, 共成功鑒定24個門,主要包括變形菌門(Proteobacteria)、擬桿菌門、藍藻門(Cyanobacteria)和后壁菌門(Firmicutes)等(表1), 其中變形菌門在各個月份中一直為優勢菌門, 甚至在5月份達到了4個檢測月份中最高的65.29%。其次為擬桿菌門, 其出現頻次百分比隨著月份的增加而遞增,與之變化趨勢相同的還有梭桿菌門(Fusobacteria)。郭世奎等(2010)在利用實時熒光定量PCR法比較直腸癌患者與正常人的腸道中的擬桿菌屬(Bacteroides)、梭桿菌屬(Fusobacterium)和梭菌屬(Clostridium)時, 發現直腸癌患者的腸道中的三種菌屬均高于正常人, 有共同上升的趨勢, 呈現出致病性的高相關性。另外, 放線菌門(Actinobacteria)、疣微菌門(Verrucomicrobia)、熱袍菌門(Thermotogae)和廣古菌門(Euryarchaeota)等也呈現出相同的變化趨勢, 均為前三個月份遞增, 10月份下降的趨勢。
2.2 擬桿菌門及其綱的分布特點
經過對454測序結果統計分析發現, 擬桿菌門下共鑒定出3個綱(目), 分為為擬桿菌綱(目)、黃桿菌綱(目)和鞘脂桿菌綱(目), 擬桿菌目下共有3個科5個屬,黃桿菌目下共有2個科17個屬, 鞘脂桿菌目下共有4個科5個屬。
四個檢測月份各個排污口擬桿菌綱、黃桿菌綱與鞘脂桿菌綱的分布情況如圖1, 其中8月份細菌檢出頻次較其它三個檢測月份明顯減少, 推測可能和采樣時的水溫有關, 四個檢測月份大部分站點的優勢屬為黃桿菌綱, Liu等(2013)對春天浮游植物暴發后的黃海海域進行了細菌群落結構的分析, 結果顯示黃桿菌綱也是其優勢菌種之一。其次為擬桿菌綱, 鞘脂桿菌綱出現較少, 但在某特定站點特定月份會出現反常現象, 比如5號站點排污口50m處8月份時,鞘脂桿菌綱出現的次數和黃桿菌綱基本相同, 而擬桿菌綱出現極少。
利用R For Windows 3.0 對各個排污口的擬桿菌門出現情況進行聚類分析, 見圖2。結果發現細菌群落結構存在較明顯時空變化, 10個站點擬桿菌門出現的情況大致可以分為兩類, 一類是S1、S3、S9、S5和S2站點, 另一類是S4、S8、S7、S10和S6站點。其中S1與S3站點、S2與S5站和S4與S8站點相似
度最高, 從排污口類型來看, S1與S3站點均為工業性排污口, S4與S8排污口均為排污河, 故推測可能與排污口類型存在較大的聯系。而從采樣月份來看, 相關度最高的為8月份和10月份, 而3月份和5月份也存在一定的相關性, 這表明擬桿菌門的出現情況與季節也有一定的聯系, 但對于所有排污口而言, 菌群結構在不同季節的變化比空間的不同所形成的差異更為顯著。Díez-Vives等(2014)通過對不同時間分布、不同空間分布的擬桿菌進行群落結構分析, 也認為不同的地點和不同的季節會導致擬桿菌群落結構的變化。
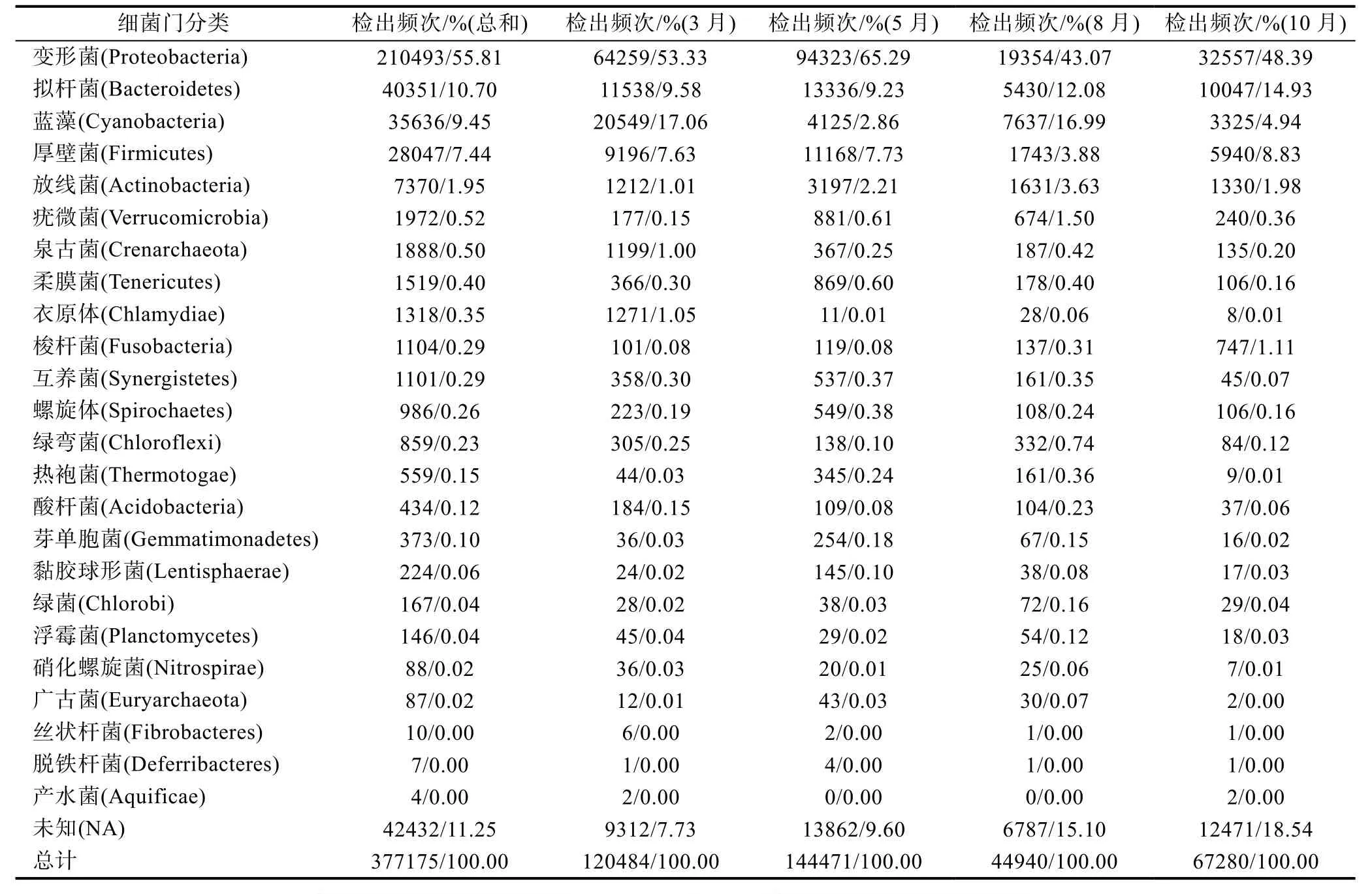
表1 454測序結果總體情況分析Tab.1 Results of the pyrosequencing (454 Life Sciences)
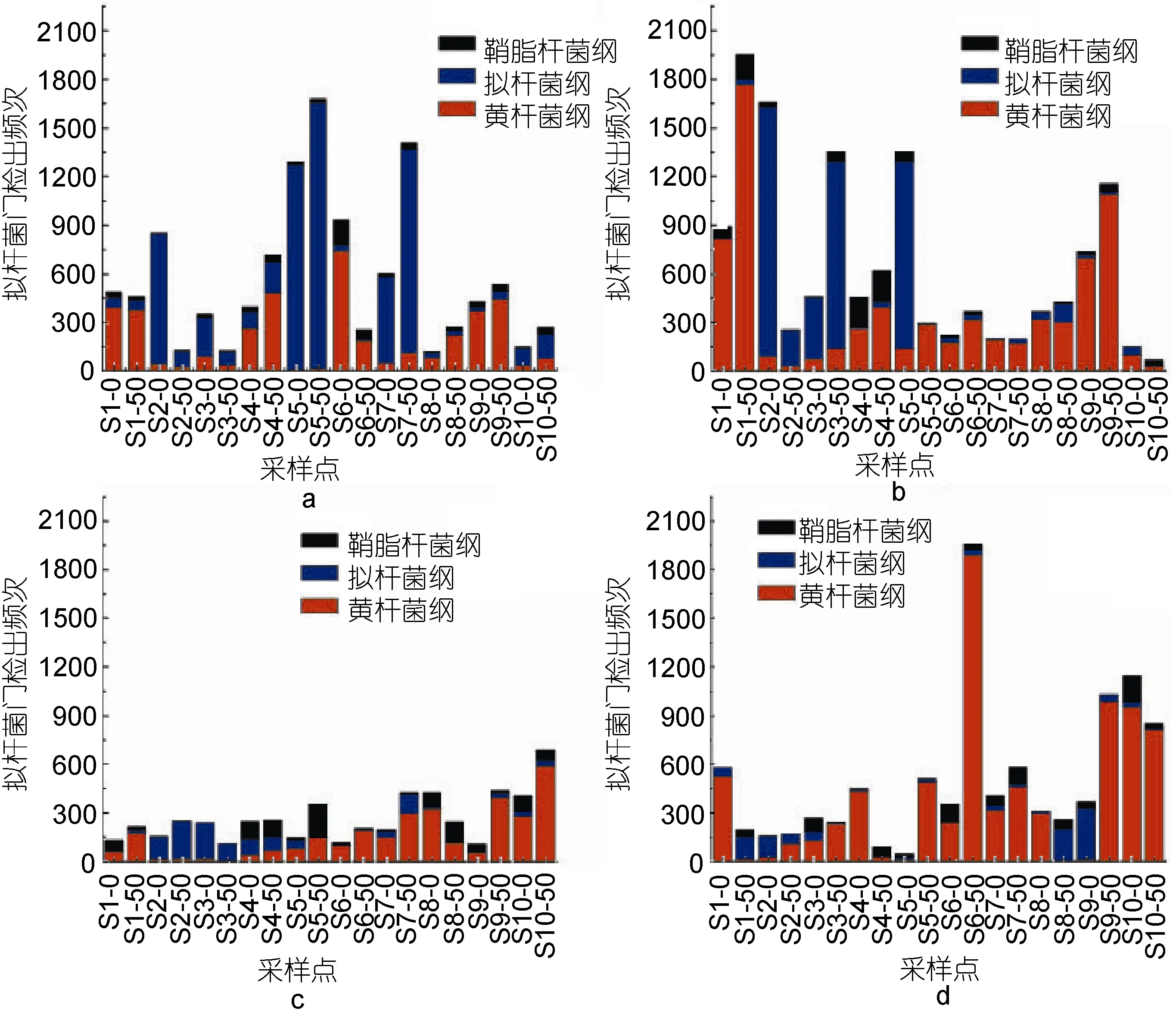
圖1 不同檢測月份不同排污口擬桿菌綱、黃桿菌綱和鞘脂桿菌綱分布情況Fig.1 The distribution statistics of Bacteroidetes, Flavobacteria and Sphingobacteria in different sewage outlets and months a. 3月; b. 5月; c. 8月; d. 10月
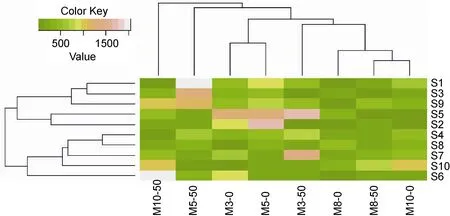
圖2 不同檢測月份不同排污口站位擬桿菌門數量聚類分析Fig.2 The cluster analysis on number of Bacteroidetes in different sewage outlets and months
將排污口和排污口50m處擬桿菌綱、鞘脂桿菌綱與黃桿菌綱出現的次數進行整合, 針對不同類型排污口進行統計分析, 如圖3所示。擬桿菌綱在2個排污口(S5和S7)呈現出隨著月份的推進而大幅下降的趨勢, 而在另外兩個排污口(S2和S3)呈現出先增后降的趨勢, 其余站點較上述四個站點變化幅度較為平緩, 各個月份變化波動很小。傳統的指示菌逐漸暴露出與沙門氏菌等致病菌的相關性差、無法辨識糞源污染(Nobleet al, 2001), 擬桿菌以其在腸道內數量的絕對優勢, 在受污染的水體中不能存活很長時間(約1—2d)、并不能繁殖(Converseet al, 2009), 且通過16S rRNA基因序列的差異能夠分辨糞源的污染來源是人類還是畜類等特點(張薇薇等, 2010), 為指示水體所受的糞源感染提供了一種新的途徑(Wadeet al,2006)。從各個排污口看, S5象山墻頭綜合排污口在3月份糞源污染最嚴重, 隨著時間的推進, 糞源污染逐步減輕, 而S2象山水桶岙垃圾場滲透液排污口在5月份糞源污染較為嚴重。
而黃桿菌綱在不同類型的排污口中的變化幅度雖不同, 但是呈現出較為一致的變化趨勢, 即大部分站點5月份的檢出頻次普遍高于3月份, 10月份的檢出頻次普遍高于8月份, 而8月份較5月份呈現出下降的趨勢, 推測可能和取樣時的水溫有一定關系。從排污口類型看, S2象山水桶岙垃圾場滲透液排污口作為市政排污口, 與其它9個排污口呈現出明顯的不同,黃桿菌綱出現次數很少, 推測可能與排污口污水中的成分有關。
與黃桿菌綱和擬桿菌綱不同, 鞘脂桿菌綱出現頻次較低, 且不同月份之間變化的幅度并沒有之前兩種菌的大, 次數最多的只有S4象山西周工業園區綜合排污口在5月份的381次。由于鞘脂桿菌綱屬于寡營養細菌, 分離和純化十分困難, 和振花等(2011)將北極海水樣品放置于連續透析培養裝置中, 4°C培養3個月之后, 以0.22μm硝酸纖維膜過濾至富集,再將濾膜置于寡營養瓊脂上2°C培養4月, 獲得可培養寡營養細菌。454高通量測序技術的出現, 克服了環境中大多數微生物“存活但不能培養”(viable but nonculturable, VBNC)的特點(段曌等, 2012), 使得大規模的環境基因組研究相繼展開, 大量的新的微生物種群和新的基因得以發現, 體現了454高通量測序較傳統微生物生態學研究方法的優勢。
2.3 普雷沃菌屬及黃桿菌屬分布特點
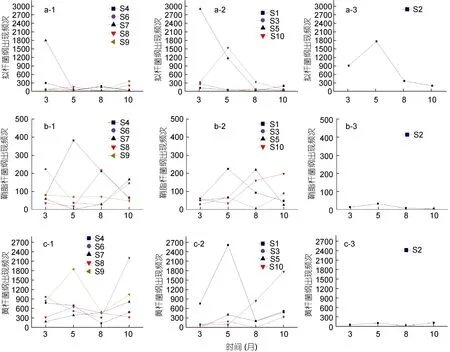
圖3 不同類型排污口擬桿菌綱、鞘脂桿菌綱和黃桿菌綱在不同檢測檢測月份出現情況Fig.3 Appearance of Bacteroidetes, Sphingobacteria and Flavobacteria in different types of sewage outlets during the four months
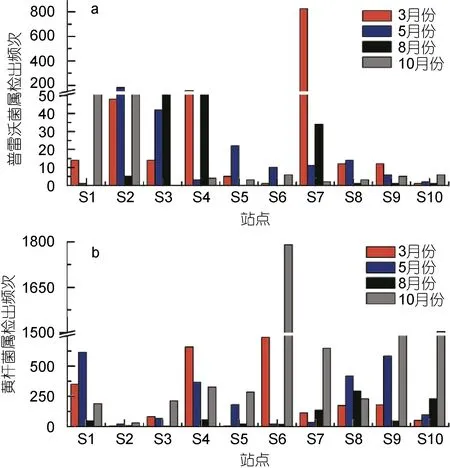
圖4 10個站點4個檢測月份普雷沃菌屬和黃桿菌屬檢出情況Fig.4 The appearance of Prevotella and Flavobacterium in the 10 sites during the four months
對454測序結果的進一步分析發現, 普雷沃菌屬(Prevotellasp.)則為擬桿菌科內除擬桿菌屬外的優勢屬, 而黃桿菌屬(Flavobacteriumsp.)為黃桿菌科內的優勢屬。對10個站點進行分析發現(圖4), S2、S3、S4與S7站點普雷沃菌屬出現頻次相對其余站點較高,其中S7站點在3月份更是高達824次, 這可能與排污口的類型有極大的聯系。普雷沃菌屬是近年來從類桿菌屬中分出的一個新菌屬, 主要集聚于正常人體的口腔、女性生殖道等部位, 組成這些部位的正常菌群僅次于脆弱類桿菌。它是臨床上較常見的一種條件致病菌, 可引起這些部位的內源性感染, 女性生殖道及口腔感染多見, 與結締組織的分解有關(Purusheet al, 2010)。
與普雷沃菌屬相比, 黃桿菌屬出現的頻次明顯較高, S6站點在10月的出現頻次將近1800次。黃桿菌屬在大部分站點的5月與8月的樣品中檢出頻次會遠遠小于3月與10月。柱狀黃桿菌(Flabobacterium columnare)主要危害魚類的鰓組織, 也可引起體表潰亂, 是細菌性爛鰓病的主要病原(黃冠軍等, 2012),同時廣泛存在于自然界及醫院環境中, 主要引起早產兒及新生兒的腦膜炎, 成人則可發生敗血癥及心內膜炎、傷口感染、肺部感染等, 是醫院感染病原菌之一(張德忠等, 2009)。
3 結論
本研究利用454高通量測序技術對寧波沿海2個重點排污口、8個一般排污口的20個站位4個檢測月份的水樣進行分析, 共成功鑒定24個菌門, 在整個454高通量測序結果的背景下, 著重對擬桿菌門及其綱、具有代表性的普雷沃菌屬及黃桿菌屬的分布特點進行了統計分析, 體現了454測序高通量的特點,另外通過454測序檢測到了極難培養的寡營養細菌鞘脂桿菌, 也說明了454測序較傳統微生物生態學研究方法的優勢。
吳彥彬, 李亞丹, 李小俊等, 2007. 擬桿菌的研究及應用. 生物技術通報, 1: 66—69
張德忠, 溫建艷, 周文聰等, 2009. 黃桿菌屬醫院感染特性與多藥耐藥分析. 中華醫院感染學雜志, 19(15): 2040—2043
張薇薇, 楊晶艷, 王 嵬等, 2010. 擬桿菌屬實時熒光定量PCR的建立. 現代預防醫學, 37(17): 3310—3312
陳麗萍, 侯付景, 張迪駿等, 2013. 寧波沿海陸源排污口假單胞菌屬. 海洋與湖沼, 44(4): 926—930
和振花, 楊季芳, 陳吉剛等, 2011. 北極海水中可培養寡營養細菌多樣性. 海洋湖沼通報, 4: 58—67
周 君, 劉 兵, 李春麗等, 2012. 寧波港壓載水浮游植物多樣性的研究. 生物技術通報, 9: 197—201
段 曌, 肖 煒, 王永霞等, 2012. 454測序技術在微生物生態學研究中的應用. 微生物學雜志, 31(5): 76—81
郭世奎, 包維民, 龔昆梅等, 2010. 實時熒光定量PCR法研究結直腸癌患者腸道擬桿菌屬、梭桿菌屬和梭菌屬量的變化.中國微生態學雜志, 22(1): 24—28
黃冠軍, 饒朝龍, 劉衍鵬等, 2012. 柱狀黃桿菌常規PCR檢測體系的建立. 水產科學, 30(11): 689—692
Converse R R, Blackwood A D, Kirs Met al, 2009. Rapid QPCR-based assay for fecalBacteroidesspp. as a tool for assessing fecal contamination in recreational waters. Water research, 43(19): 4828—4837
Díez-Vives C, Gasol J M, Acinas S G, 2014. Spatial and temporal variability among marine Bacteroidetes popula-tions in the NW Mediterranean Sea. Systematic and Applied Microbiology, 37(1): 68—78
Li Y, Lee C G, Watanabe Tet al, 2011. Identification of microbial communities that assimilate substrate from root cap cells in an aerobic soil using a DNA-SIP approach. Soil Biology and Biochemistry, 43(9): 1928—1935
Lin P Y, Chen H L, Huang C Tet al, 2010. Biofilm production,use of intravascular indwelling catheters and inappropriate antimicrobial therapy as predictors of fatality inChryseobacterium meningosepticumbacteraemia. International Journal of Antimicrobial Agents, 36(5): 436—440
Liu M, Xiao T, Sun Jet al, 2013. Bacterial community structures associated with a natural spring phytoplankton bloom in the Yellow Sea, China. Deep Sea Research Part II: Topical Studies in Oceanography, 97: 85—92
Margulies M, Egholm M, Altman W Eet al, 2005. Genome sequencing in microfabricated high-density picolitre reactors.Nature, 437: 376—380
Noble R T, Fuhrman J A, 2001. Enteroviruses detected by reverse transcriptase polymerase chain reaction from the coastal waters of Santa Monica Bay, California: low correlation to bacterial indicator levels. Hydrobiologia, 460(1—3): 175—184
Palenik B, Ren Q, Tai Vet al, 2009. CoastalSynechococcus metagenomereveals major roles for horizontal gene transfer and plasmids in population diversity. Environmental Microbiology, 11: 349—359
Purushe J, Fouts D E, Morrison Met al, 2010. Comparative genome analysis ofPrevotella ruminicolaandPrevotella bryantii: insights into their environmental niche. Microbial Ecology, 60(4): 721—729
Roesch L F, Fulthorpe R R, Riva Aet al, 2007. Pyrosequencing enumerates and contrasts soil microbial diversity. The ISME Journal, 1: 283—290
Wade T J, Calderon R L, Sams Eet al, 2006. Rapidly measured indicators of recreational water quality are predictive of swimming-associated gastrointestinal illness. Environmental Health Perspectives, 114(1): 24—28
Wexler H M, 2012. Pump it up: Occurrence and regulation of multi-drug efflux pumps inBacteroides fragilis. Anaerobe,18(2): 200—208
Wood J, Scott K P, Avgu?tin Get al, 1998. Estimation of the relative abundance of differentBacteroidesandPrevotellaribotypes in gut samples by restriction enzyme profiling of PCR-amplified 16S rRNA gene sequences. Applied and Environmental Microbiology, 64(10): 3683—3689
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
