地衣芽孢桿菌(Bacillus licheniformis)對鎘脅迫下泥蚶(Tegillarca granosa)的抗氧化酶活性和免疫相關基因表達的影響*
2014-03-19 10:18:18沈靈靈全晶晶
海洋與湖沼 2014年5期
王 稷 李 曄 瞿 洋 沈靈靈 孫 靜 全晶晶
(寧波大學海洋學院 寧波 315211)
近年來, 隨著沿海工業的高速發展, 周邊城鎮生活污水和工業廢水排放入海的增加, 致使灘涂貝類養殖環境受到不同程度的重金屬污染。重金屬鎘(Cd)是廣泛分布于環境的污染物, 通常沉積在河流、灘涂和近岸水體中, 極易被水生生物吸收和蓄積, 并通過食物鏈傳遞至高等生物體內。Cd對人體健康危害嚴重, 尤其損害腎臟、肝臟、心血管系統以及生殖系統(Korte, 1983; Junget al, 2005; Chenet al, 2012; Nairet al, 2012)。
泥蚶(Tegillarca granosa), 俗稱花蚶、血蚶、銀蚶、蚶子等, 富含血紅蛋白、牛磺酸和甜菜堿等生物活性成分, 有補血、溫中、健胃的功效, 且肉味鮮美, 是沿海地區的傳統美食(Baoet al, 2010)。由于泥蚶屬于濾食性動物, 且營底棲生活, 使得它更加容易富集水體中的重金屬等污染物(Junget al, 2012)。調查研究發現寧波地區泥蚶中Cd含量超標, 其質量指數為1.85,各地超標率達100%(尤仲杰等, 2011)。若不解決泥蚶中Cd超標問題, 將直接影響到泥蚶養殖業的可持續發展及泥蚶的食用安全性。因此如何減少Cd在貝類體內殘留或有效脫除Cd已是當務之急, 這對保障我國貝類水產品的質量安全有重要意義。
目前貝類凈化的方法多數針對致病性微生物,對于重金屬污染還未見有效凈化處理技術(葉盛權等,2003)。微生物修復治理重金屬污染具有安全性、經濟性和非破壞性的優點, 是重金屬污染治理的重要研究方向之一。目前, 國內外研究發現一些微生物如細菌、真菌和藻類等對重金屬Cd2+有很強的耐受性和富集能力, 研究的細菌多為假單胞菌屬(Pseudomonassp.)(Chienet al, 2011; Weiet al, 2011)、芽孢桿菌屬(Bacillussp.) (付瑾等, 2011)和皮氏羅爾斯通氏菌屬(Ralstoniasp.) (孫晶等, 2009; Nies, 2000)。其中地衣芽孢桿菌(Bacillus licheniformis), 革蘭氏陽性、兼性厭氧菌, 在生長代謝中能產生多種抗菌活性物質, 可抑制腸道中的致病菌, 常用其制備益生菌, 用于調整腸道菌群失調(周鳴等, 2006; 何英, 2007)。在重金屬污染防治方面, 周鳴等(2006)采用地衣芽孢桿菌死菌體吸附水中的鉻(Cr6+), 最大吸附量為60.5mg/g。
通過活化培養實驗室馴養的耐350mg/L Cd2+的地衣芽孢桿菌, 研究其對Cd2+脅迫下泥蚶存活率、組織中Cd殘留量、抗氧化酶系活性及免疫相關基因表達情況的影響, 進而探討微生物菌劑對貝類Cd污染的防治和脫除的有效性及可行性, 并為提高水產食品食用安全性提供參考。
1 材料與方法
1.1 材料
1.1.1 地衣芽胞桿菌 地衣芽孢桿菌(Bacillus licheniformis)由寧波大學海洋學院實驗室篩選鑒定保存。
1.1.2 泥蚶 實驗用泥蚶(Tegillarca granosa)采自溫州樂清, 平均體重為(8.50±0.23)g, 實驗前洗刷干凈, 清除附著物。
1.1.3 試劑 實驗用硝酸(HNO3)、氯化鎘(CdCl2?2H2O)為分析純, 購自寧波博奧生物工程有限公司);SOD試劑盒、GSH-Px試劑盒、BCA試劑盒購自南京建成生物工程研究所; M-MuLV第一鏈cDNA合成試劑盒由生工生物工程(上海)有限公司提供; SYBR Premix Ex TaqTMⅡ為TaKaRa公司產品。
1.2 方法
1.2.1 泥蚶的處理 泥蚶在20°C、鹽度30的海水中充氣暫養一周, 每天投喂適量餌料, 隔天換水。實驗時泥蚶被隨機分為14組, 每組20枚。根據Cd2+對泥蚶96h的半致死濃度(劉琴, 2008), 將Cd2+在實驗水體終濃度設置為0.437、1.749和6.996mg/L; 每組地衣芽孢桿菌的投喂量分別為6.24×109、1.56×1010、4.50×1011CFU/L (水體中的終濃度)。不加Cd2+,也不投喂地衣芽孢桿菌的為空白組, 而不加Cd2+, 僅投喂4.50×1011CFU/L地衣芽孢桿菌的作為對照組。Cd2+和地衣芽胞桿菌在實驗開始時加入, 此后每次換水時加入相應的量。實驗水體為1L, 每個實驗組做2個平行, 實驗周期為96h。
1.2.2 菌種培養 實驗時將活化的地衣芽孢桿菌接種到普通LB液體培養基中, 搖床培養24h, 菌液經5000r/min離心5min, 菌體沉淀用無菌水制備成3.01×1012CFU/L的菌懸液, 于4°C保存備用。
1.2.3 泥蚶死亡率的統計 每天觀察泥蚶的死亡情況, 96h后統計各組泥蚶的死亡率。
1.2.4 組織中Cd殘留量的測定 每組泥蚶隨機取5枚, 解剖內臟團并搗碎后, 進行微波消解。每0.5g樣品加入5mL HNO3消解試劑。微波消解采用梯度升溫消解程序: 800W從室溫經10min升至120°C, 保持5min, 然后從120°C再經5min升至150°C, 保持5min,最后從150°C經5min升至190°C, 保持15min。冷卻后, 消解液用5% HNO3定容至50 mL。ICP-AES測消解液中Cd含量。實驗中所有器皿使用前均于5%HNO3中浸泡24 h。
1.2.5 泥蚶組織中抗氧化酶活力測定 冰上解剖泥蚶內臟團, 冰凍生理鹽水沖洗干凈, 吸干水分后,準確稱取組織質量, 按質量體積比1︰9加入預冷生理鹽水, 冰上勻漿, 勻漿液稀釋10倍后, 離心(3500r/min,4°C, 10min), 取上清液, 再次稀釋為1%和10%濃度,分別用于總蛋白濃度和SOD、GSH-Px酶活的測定。實驗步驟參照試劑盒說明。
1.2.6 免疫相關基因的表達分析
(1) 總RNA的提取 泥蚶內臟組織, 在液氮中充分研磨, 加入Trizol提取總RNA, 具體步驟參照Trizol說明書。總RNA使用Nanodrop超微量分光光度計進行定量分析, 用1.5%瓊脂糖凝膠電泳確認完整性。
(2) cDNA的合成 上述提取的總RNA, 根據M-MLV使用說明進行第一鏈cDNA的合成, 產物于–20°C保存備用。
(3) Real-time PCR 根據NCBI上的序列信息,采用Primer Premier 5.0軟件設計并合成泥蚶的MT、Ferritin、sHSP基因的熒光定量PCR引物(表1), β-actin作為內參。熒光定量PCR擴增體系為20μL: SYBR Premix Ex TaqTMⅡ(2×), 10μL; PCR Forward Primer(10μmol/L), 1μL; PCR Reverse Primer (10μmol/L), 1μL;cDNA模板, 4μL; ddH2O, 4μL。反應程序為95°C 1min;95°C 10s; 60°C 15s; 75°C 15s, 30至40個循環。以actin為內參, 對得到的各樣品Ct值進行均一化處理, 以正常生長下的泥蚶的相關免疫基因的mRNA的表達量為基準, 應用2–ΔΔCT法確定各個基因 mRNA的相對含量。

表1 用于Real-time PCR的引物序列Tab.1 Sequence of primers used for real-time PCR
2 結果
2.1 地衣芽胞桿菌對泥蚶存活率和鎘殘留量的影響
不同濃度的Cd2+和地衣芽胞桿菌對泥蚶存活率和組織中的鎘殘留量的影響見表2。
Cd2+濃度為0 mg/L時, 4.50×1011CFU/L地衣芽孢桿菌作用下泥蚶的存活率與自然生長狀態下(即空白組)的存活率一樣, 均為100.0%, 表明地衣芽胞桿菌對泥蚶無負面影響。未加地衣芽孢桿菌時, 不同濃度Cd2+的加入對泥蚶存活率的影響具有顯著差異(P<0.05), Cd2+濃度越高, 泥蚶存活率越低。隨著地衣芽胞桿菌投入量的增加, 一定濃度Cd2+脅迫下的泥蚶的存活率逐漸上升。以上數據表明地衣芽孢桿菌的加入對提高Cd2+脅迫下泥蚶的存活率具有顯著的效果。
在養殖水體中分別加入濃度為0.473、1.749和6.996 mg/L的Cd2+, 泥蚶組織中的Cd含量相應地為0.690、1.030和4.470mg/kg (表2)。表明水體中的不同濃度的Cd2+在泥蚶體內引起了不同程度的蓄積。當加入地衣芽孢桿菌后, 泥蚶體內的Cd殘留量發生顯著下降(P<0.05)(表2)。數據證實地衣芽胞桿菌能夠在一定程度上清除泥蚶體內的Cd殘留量。
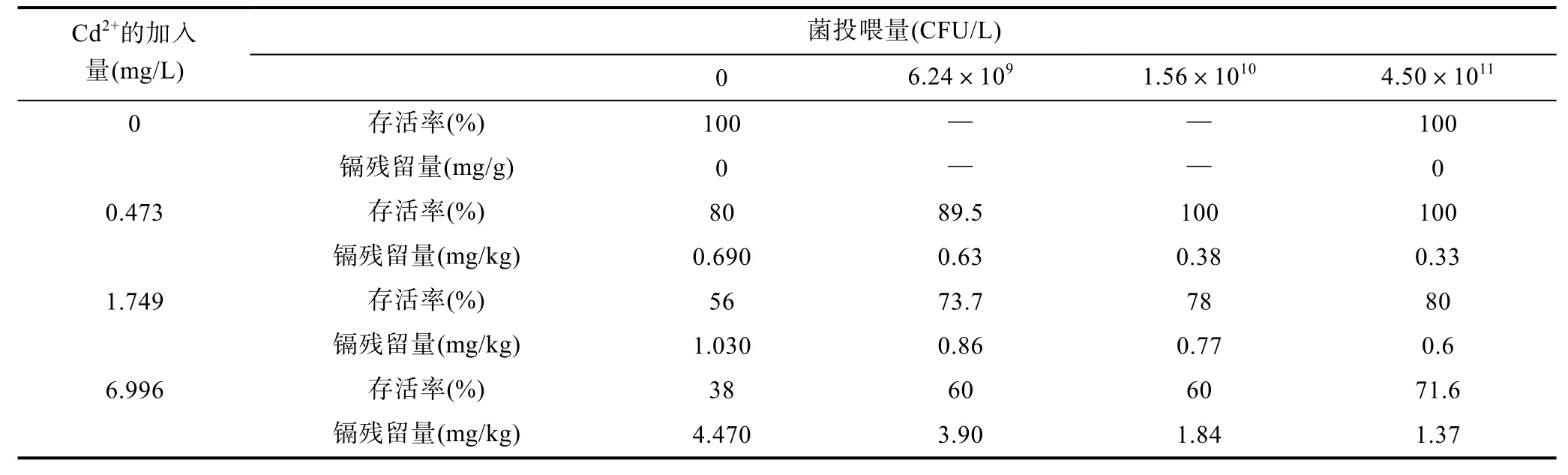
表2 地衣芽胞桿菌對鎘脅迫下泥蚶的存活率和組織中鎘殘留量的影響Tab.2 Influences of B. licheniformis on the survival rates of T. granosa and the Cd concentration in T. granosa exposed to cadmium chloride
2.2 地衣芽胞桿菌對抗氧化酶活性的影響
實驗選擇0.437 mg/L Cd2+脅迫下的泥蚶進一步研究不同濃度的地衣芽胞桿菌對抗氧化酶SOD和GSH-Px活性的影響, 結果見圖1。當暴露于0.437 mg/L Cd2+的泥蚶被投喂不同量(6.24×109, 1.56×1010, 4.50×1011CFU/L)的地衣芽孢桿菌后, 泥蚶組織中SOD活性呈顯著升高趨勢, 且與加入的地衣芽胞桿菌的量呈正相關。地衣芽胞桿菌對GSH-Px活性的影響與對SOD的影響類似, 也表現為隨加入濃度增加而活性升高的趨勢。
2.3 地衣芽胞桿菌對泥蚶體內免疫相關基因表達的影響
同樣, 選擇Cd2+濃度為0.473 mg/L的實驗組泥蚶, 研究不同投喂量的地衣芽胞桿菌對組織中三類免疫相關基因MT、ferritin和sHSP的表達情況, 結果見圖2。隨著地衣芽胞桿菌投喂量的增加, 三類基因的表達量均呈顯著下降趨勢, 其中MT的表達量下降最為顯著, sHSP次之, ferritin表達量的變化最小。表明這三類基因在泥蚶體內起著相似的免疫調節作用, 而地衣芽胞桿菌是通過調控免疫相關基因的表達, 從而在一定程度上提高泥蚶耐受鎘脅迫的能力。
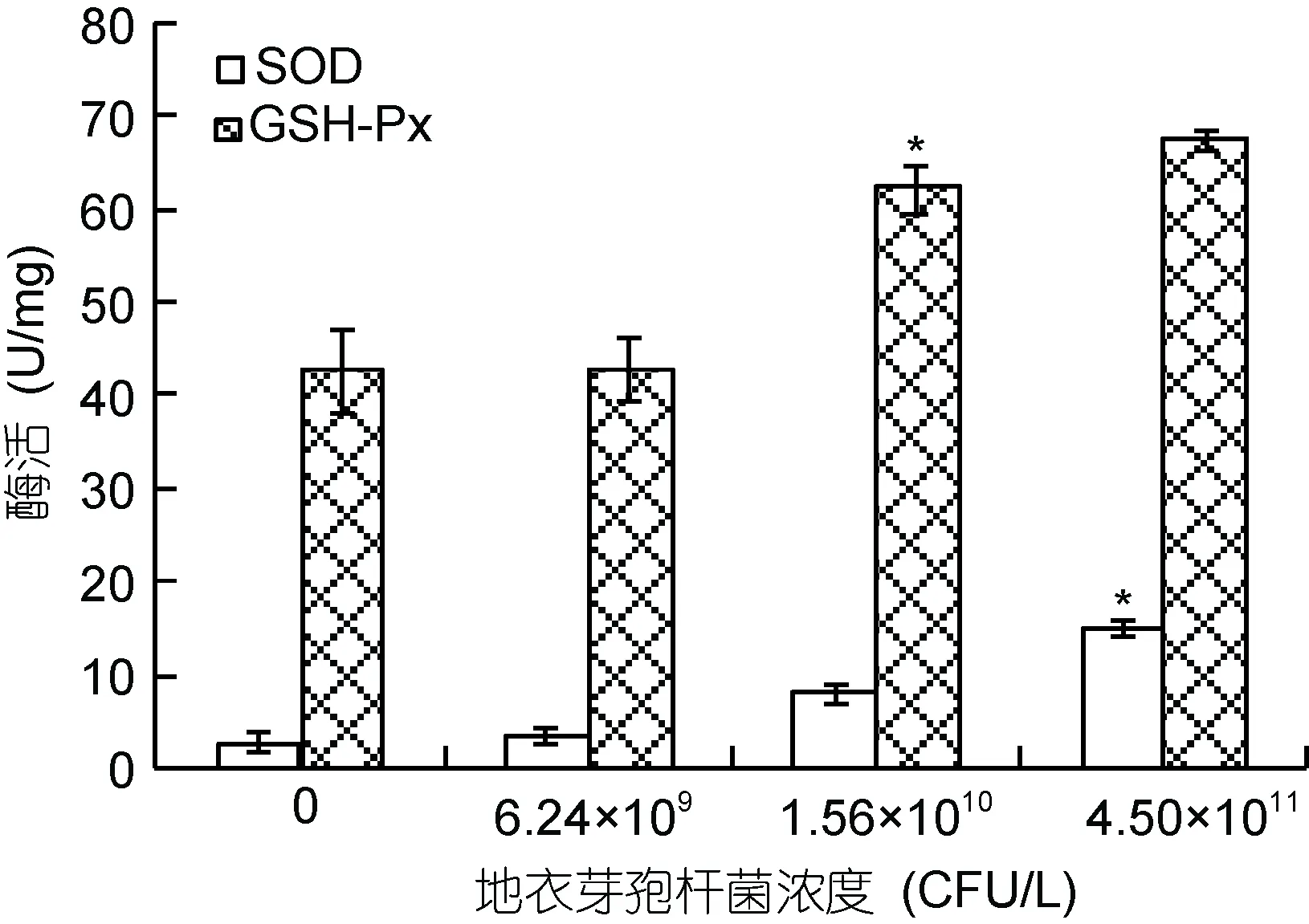
圖1 地衣芽孢桿菌對泥蚶組織中SOD和GSH-Px活性的影響Fig.1 Effects of B. licheniformis on the activities of SOD and GSH-Px in the visceral mass of T. granosa exposed to 0.473mg/L cadmium
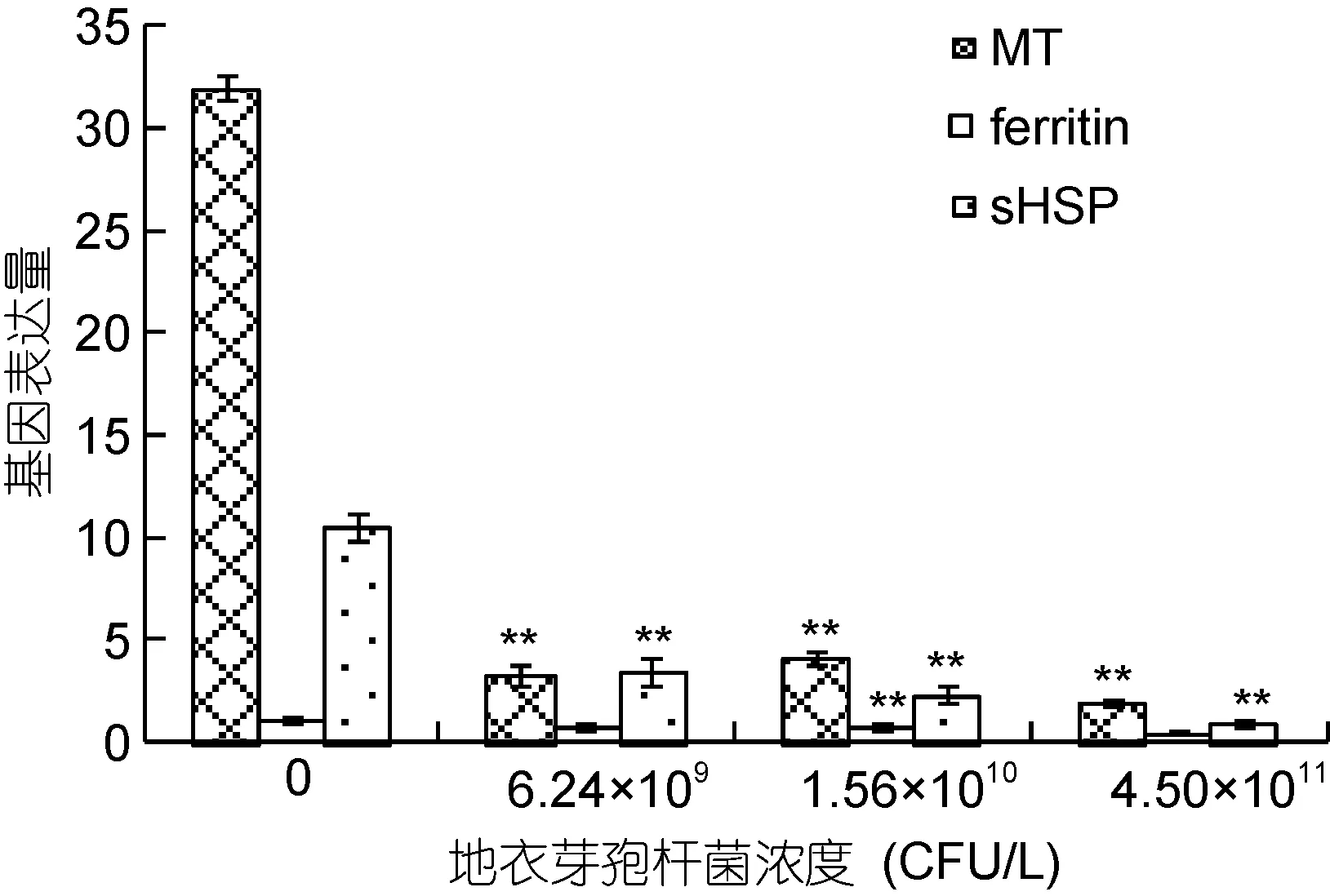
圖2 地衣芽孢桿菌對泥蚶組織中免疫基因表達的影響Fig.2 Effects of B. licheniformis on the expression levels of MT,sHSP and ferritin in the visceral mass of T. granosa exposed to 0.437 mg/L cadmium
3 討論
泥蚶是我國水產養殖中極具經濟價值的貝類,屬灘涂濾食性貝類, 經過鰓不斷吸收溶解水體中的重金屬進入體內。此外, 也可通過體表與水體的滲透交換作用使重金屬進入體內, 并在內臟積累(張傳永等, 2008)。因此泥蚶的生活環境在很大程度上決定了泥蚶體內Cd的含量。Cd在泥蚶體內的累積能破壞泥蚶體內的離子平衡, 改變細胞膜滲透性, 也可能抑制鈉離子-鉀離子(Na+/K+)主動運輸過程, 從而降低ATP的有效性。此外, Cd能抑制泥蚶抗氧化酶系統, 破壞內臟器官酶系統的正常生理功能, 妨礙泥蚶正常代謝(李學鵬等, 2010), 致使泥蚶存活率下降。另一方面泥蚶體內的Cd也容易通過食物鏈進入人體內, 影響泥蚶的食用安全性。
很多微生物包括細菌、真菌、酵母以及藻類, 都具有不同程度的結合重金屬的能力(Dhankharet al,2011)。研究普遍認為微生物的細胞壁是吸收或結合重金屬的主要結構和場所(Pradhanet al, 2007; Fanget al, 2009)。
本實驗將地衣芽胞桿菌投喂給鎘脅迫下的泥蚶,結果使泥蚶的存活率顯著上升(表2), 而體內的Cd殘留量顯著下降(表2), 表明泥蚶對地衣芽胞桿菌的吸收能夠減少Cd在體內的蓄積, 起到保護泥蚶的作用。此結果一方面是因為投放到水體中的地衣芽胞桿菌直接結合了水體中的Cd2+, 從而減少了泥蚶對Cd2+的吸收; 另一方面可能是進入泥蚶體內的地衣芽胞桿菌通過結合體內的Cd, 加速了Cd的排泄。
益生菌已被廣泛地應用在養殖業中, 對提高養殖動物的生長率、免疫力以及對病原菌的抵抗性具有重要作用。大量研究表明飼料中添加益生菌能夠有效調節動物的抗氧化系統, 影響相關免疫基因的表達模式(Bruntet al, 2008; Tovar-Ramírezet al, 2010;Sliva-Aciareset al, 2013)。為此, 本實驗分析了地衣芽胞桿菌作用下, 泥蚶的抗氧化酶活性和相關基因的表達變化情況, 以了解該菌株對泥蚶的保護機制。
有研究發現, 在Cd2+脅迫下的動物體內會產生大量的活性氧(Reactive oxygen species, ROS), 進而耗盡抗氧化酶(Shaikhet al, 1999; Bagchiet al, 2000;Szuster-Ciesielskaet al, 2000)。因此一般認為能夠提高抗氧化酶活性或者減少重金屬吸收的物質都能夠降低Cd2+的誘導毒性(Lacorteet al, 2013)。在本實驗中, 作者發現細菌的投喂量與泥蚶體內的SOD和GSH-Px的活性呈正相關(圖1), 由此推斷, 地衣芽胞桿菌通過增加抗氧化酶活性來減少ROS對泥蚶的傷害作用, 從而降低Cd誘導的毒性。
同時, 研究還發現地衣芽胞桿菌的投喂導致了三類基因MT、sHSP和ferritin在mRNA水平上表達發生顯著下調(圖2)。MT是一種廣泛存在于生物體內的金屬結合蛋白, 可調節生物細胞體內非必需金屬元素的解毒過程; Ferritin是一種高度保守的鐵結合蛋白, 可對機體氧化損傷起保護作用; sHSP是HSP家族中的一員, 也能作用于細胞來抵抗各種應激反應。有研究報告, 當泥蚶受到Cd2+脅迫時, MT、sHSP和ferritin往往是高表達的。在本實驗中, 當水體中加入地衣芽胞桿菌后, 進入泥蚶體內的Cd2+的量減少,可能是導致這三類基因表達下調的原因。事實上, 在很多情況下基因在mRNA水平上的表達和蛋白水平上的表達并不一致(Sekharet al, 2011), 因此, 要闡明地衣芽胞桿菌作用機制, 進一步研究這些免疫基因在蛋白水平上的表達是必不可少的。
尤仲杰, 康飛金, 王建萍等, 2011. 寧波地區灘涂貝類養殖區環境及貝體重金屬含量與評價. 海洋環境科學, 30(4): 508—511
葉盛權, 黃 甫, 2003. 雙殼貝類的凈化技術研究. 食品研究與開發, 24(4): 28—30
付 瑾, 謝學輝, 錢 林等, 2011. 皮氏羅爾斯通氏菌株DXT3-01的耐鎘性能及鎘富集機理. 應用與環境生物學報,17(5): 717—721
劉 琴, 2008. 重金屬Pb、Cd和Cr在泥蚶中的行為研究. 青島: 中國海洋大學碩士學位論文, 16—21
孫 晶, 李 紅, 李雪龍, 2009. 整腸生中地衣芽孢桿菌對幾種常見抗菌藥物的體外敏感性. 中國臨床藥理學雜志,25(3): 246—248
李學鵬, 段青源, 勵建榮, 2010. 我國貝類產品中重金屬鎘的危害及污染分析. 食品科學, 17(31): 457—461
何 英, 2007. 地衣芽孢桿菌膠囊聯合制霉菌素治療兒童真菌性腸炎療效觀察. 中國藥房, 18(11): 852—857
張傳永, 劉 慶, 陳燕妮, 2008. 重金屬對水生生物毒性作用研究進展. 生命科學儀器, 6(11): 3—7
周 鳴, 劉云國, 李 欣, 2006. 地衣芽孢桿菌對Cr6+的吸附動力學研究. 應用與環境生物學報, 12(1): 84—87
Bagchi D, Joshi S S, Bagchi Met al, 2000. Cadmium- and chromium-induced oxidative stress, DNA damage, and apoptotic cell death in cultured human chronic myelogenous leukemic K562 cells, promyelocytic leukemic HL-60 cells,and normal human peripheral blood mononuclear cells. J Biochem Mol Toxicol, 14: 33—41
Bao Y, Lin Z, 2010. Generation, annotation, and analysis of ESTs from hemocyte of the bloody clam,Tegillarca granosa. Fish& Shellfish Immunology, 29(5): 740—746
Brunt J, Hansen R, Jamieson D Jet al, 2008. Proteomic analysis of rainbow trout (Oncorhynchus mykiss, Walbaum) serum after administration of probiotics in diets. Veterinary Immunology and Immunopathology, 121: 199—205
Chen C, Shen W, Huo Let al, 2012. Effects of cadmium on the microstructure and ultrastructure of gill and hepatopancreas inTegillarca granosa. Journal of fisheries of China, 36(4):522—528
Chien C C, Jiang M H, Tsai M R, 2011. Isolation and characterization of an environmental cadmium and tellurite resistant pseudomonas strain. Environmental Toxicology and Chemistry, 30(10): 2202—2207
Dhankhar R, Hooda A, 2011. Fungal biosorption——an alternative to meet the challenges of heavy metal pollution in aqueous solutions. Environmental Technology, 32(5): 467—491
Fang L C, Cai P, Chen W Let al, 2009. Impact of cell wall structure on the behavior of bacterial cells in the binding of copper and cadmium. Colloids and Surfaces A:Physicochemical and Engineering Aspects, 347: 50—55
Jung H B, Yun S T, Mayer Bet al, 2005. Transport and sediment-water partitioning of trace metals in acid mine drainage: an example from the abandoned Kwangyang Au-Ag mine area, South Korea. Environ Geol, 48: 437—449
Jung S L, Lee Y G, Park J Jet al, 2012. Microanatomy and ultrastructure of the foot of the infaunal bivalveTegillarca granosa(Bivalvia: Arcidae). Tissue and Cell, 44(5): 316—324
Korte F, 1983. Ecotoxicology of cadmium: general overview.Ecotoxicol Environ Saf, 7: 3—8
Nair P M G, Choi J, 2012. Effects of cadmium chloride and nonylphenol on the expression of StAR-related lipid transfer domain containing protein (START1) gene in aquatic midge,Chironomus riparius. Comp Biochem Physiol C, 155: 369—374
Nies D H, 2000. Heavy metal-resistant bacteria as extremophiles:Molecular physiology and biotechnological use ofRalstoniasp. CH34. Extremophiles, 4(2): 77—82
Pradhan S, Singh S, Rai L C, 2007. Characterization of various functional groups present in the capsule ofMicrocystisand study of their role in biosorption of Fe, Ni and Cr.Bioresource Technology, 98: 595—601
Sekhar K, Priyanka B, Reddy V Det al, 2011. Metallothionein 1(CcMT1) of pigeonpea (Cajanus cajanL.) confers enhanced tolerance to copper and cadmium inEscherichia coliandArabidopsis thaliana. Environmental and Experimental Botany, 72: 131—139
Shaikh Z A, Vu T T, Zaman K, 1999. Oxidative stress as a mechanism of chronic cadmium-induced hepatotoxicity and renal toxicity and protection by antioxidants. Toxicol Appl Pharmacol, 154: 256—263
Sliva-Aciares F, Moraga D, Auffret Met al, 2013. Transcriptomic and cellular response to bacterial challenge (PathogenicVibrio parahaemolyticus) in farmed juvenileHaliotis rufescensfed with or without probiotic diet. Journal of Invertebrate Pathology, 113(2): 163—176
Szuster-Ciesielska A, Stachura A, Slotwinska Met al, 2000. The inhibitory effect of zinc on cadmium-induced cell apoptosis and reactive oxygen species (ROS) production in cell cultures. Toxicology, 145: 159—171
Tovar-Ramírez D, Mazurais D, Gatesoupe J Fet al, 2010.Dietary probiotic live yeast modulates antioxidant enzyme activities and gene expression of sea bass (Dicentrarchus labrax) larvae. Aquaculture, 300: 142—147
Wei X, Fang L C, Cai Pet al, 2011. Influence of extracellular polymeric substances (EPS) on Cd adsorption by bacteria.Environmental Pollution, 159(5): 1369—1374
