當歸苯丙氨酸解氨酶基因片段克隆和組織特異性表達分析
2014-03-26 08:38:16雒軍王引權溫隨超李靜張金林夏琦
草業學報 2014年4期
關鍵詞:分析
雒軍,王引權*,溫隨超,李靜,張金林*,夏琦
(1.甘肅中醫學院藥學院,甘肅 蘭州730000;2.甘肅省高校中(藏)藥化學與質量研究省級重點實驗室,甘肅 蘭州730000;3.蘭州大學草地農業科技學院,甘肅 蘭州730020)
當歸(Angelicasinensis)為傘形科當歸屬草本植物,以干燥根入藥,具有補血活血,調經止痛,潤腸通便等功效,為臨床常用藥[1]。甘肅是當歸主產區,年栽培面積和總產量都占全國90%以上,當歸適宜栽培在2000~2800 m海拔的高寒陰濕地帶[2]。目前對當歸栽培模式、當歸生理特征、藥材品質和產量形成進行了許多研究,王慧珍等[3]報道連作當歸比輪作當歸的揮發油含量、葉片光合色素含量和光合特性顯著降低,進而導致產量和品質下降;王田濤等[4]研究發現當歸/大蒜(Alliumsativum)間作能提高當歸產量和優等品比率,并能降低當歸麻口病發病率;邱黛玉等[5]報道當歸種苗的大小影響生長過程中當歸莖葉和根部蛋白質、游離氨基酸、可溶性糖含量的變化,進而影響成藥期抽苔率,但對當歸藥效成分生物合成及其代謝調控的分子生物學機理尚未開展過系統研究。
阿魏酸(ferulic acid,FA)是當歸藥材中含量較高的水溶性活性成分,為《中國藥典》規定的當歸質量控制指標之一[6]。現代藥理研究表明,FA具有明顯抗動脈粥樣硬化,抗血小板凝集和血栓,清除亞硝酸鹽、氧自由基、過氧化亞硝基,抗菌消炎,抗腫瘤,抗突變,增加免疫功能等作用[7]。FA為苯丙烷代謝途徑生成的中間產物之一,其主要生物合成途徑是由苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)催化L-苯丙氨酸(L-Phe)生成反式-肉桂酸(t-CA),肉桂酸-4-羥基化酶(C4H)催化反式-肉桂酸生成對-香豆酸(p-Coumaric acid),對-香豆酸經香豆酸-3-羥基化酶(C3H)作用生成咖啡酸(caffeic acid),再以甲硫氨酸(S-腺苷-L甲硫氨酸)為甲基供體,在咖啡酸-O-甲基轉移酶(COMT)的催化下生成阿魏酸(FA)[8-9]。PAL催化L-苯丙氨酸(L-Phe)生成反式-肉桂酸(t-CA),是苯丙烷代謝的第一步反應,也是其中的關鍵步驟之一[10]。目前有多種植物的PAL已經得到克隆,在藥用植物中有朝鮮當歸(Angelicagigas)[11]、皺波天竺葵(Pelargoniucrispum)[12]、金銀花(Lonicerajaponica)[13]、黃芩(Scutellariabaicalensis)[14]、紫草(Lithospermumerythrorhizon)[15]、黃芪(Astragalusmembrana-ceus)[16]和丹參(Salviamiltiorrhiza)[17]等,但尚未有當歸PAL研究的報道。本研究的重點在于用RT-PCR方法克隆當歸代謝相關PAL片段、進行序列分析,并運用實時熒光定量PCR分析其在當歸不同組織的特異性表達,為進一步獲得當歸PAL全長及其闡明FA的生物合成與制定調控策略提供理論基礎。
1 材料與方法
1.1 材料
當歸植株于2013年8月采自甘肅岷縣茶埠鎮試驗基地(海拔2780m,東經104°06′,北緯34°29′)。田間挖取當歸全株,立即裝入干冰泡沫箱中,于當日帶至實驗室后迅速用去離子水沖洗干凈,并將根、莖、葉組織分離后用液氮迅速冷凍后,保藏于-80℃超低溫冰箱,用作RNA提取材料。大腸桿菌(Escherichiacoli)DH5α菌株由本實驗室保存。
采用的分子生物學試劑主要包括:RNA提取試劑十六烷基三甲基溴化銨(CTAB)、LiCl和聚乙烯吡咯烷酮360(PVP-360)為美國Sigma產品,cDNA 合成試劑盒(TaKaRa,大連),Taq DNA Polymerase(Thermo,美國)、PCR產物回收試劑盒(TransGen,北京)、PCR 產物克隆試劑盒(Promega,美國),DNA Marker(TransGen,北京),SYBR Green熒光定量試劑盒(Promega,美國),其他生化試劑均為進口或國產分析純產品。
1.2 引物設計與合成
通過對GenBank中皺波天竺葵(X81159)、朝鮮當歸 (HM114215)、胡蘿卜(Daucuscarota)(AB435640)和擬南芥(Arabidopsisthaliana)(NM_001203294)等植物PAL編碼基因CDS序列進行同源比對,找出高度保守區段,利用Primer 6.0生物軟件設計一對擴增引物(P1,P2),用于擴增當歸PAL片段。P1:5′-GCTGAACAGCACAATCAAGATGT-3′,P2:5′-GTTAACAGATTGGAAGAGGAGCAC-3′。
參照克隆并測序的當歸PAL片段序列,利用Primer 6.0生物軟件設計一對用于PAL熒光定量PCR檢測的特異引物(P3,P4),擴增長度為119bp。P3:5′-GTGTCAACGGTGAGCTCCAT-3′,P4:5′-GCATCAATGGGTAGGTTGCG-3′。參照當歸Actin片段序列[18],設計一對熒光定量PCR內參基因檢測的特異引物(P5,P6),擴增長度為109bp。P5:5′-TGGTATTGTGCTGGATTCTGGT-3′,P6:5′-TGAGATCACCACCAGCAAGG-3′。以上所有引物均由生工生物工程(上海)股份有限公司合成。
1.3 總RNA的提取
當歸根、莖及葉組織中總RNA的提取參照文獻[19]方法進行,并略作修改。實驗中所用玻璃容器、離心管和去離子水在RNA提取前均用焦炭酸二乙酯(DEPC)處理,以變性滅活RNase。具體操作如下:
取1g材料置于含液氮和少量石英砂的研缽中充分研磨成粉末狀。取約1/10的粉末轉入內含0.9mL 65℃預熱的提取緩沖液的離心管中,提取緩沖液成分包括100mmol/L Tris-Cl、2%CTAB(m/V)、2%PVP-360(m/V)、30mmol/L EDTA、1.5mol/L的 NaCl和2% (V/V)的β-巰基乙醇(β-ME)。充分振蕩混勻后置于65℃水浴鍋孵育20min,期間每5min充分振蕩1次。冷卻至室溫后,加等體積氯仿/異戊醇(24∶1),劇烈振蕩30s,11000r/min、4℃離心10min。水相轉入另一離心管,加約1/3體積10mol/L的LiCl,混勻后置-20℃冰箱30 min。11000r/min、4℃離心10min,沉淀溶解于0.5mL無RNase的去離子水,加0.5mL水飽和酚抽提一次,再加入等體積氯仿/異戊醇(24∶1)抽提1次,11000r/min、4℃離心10min。水相轉入另一離心管,加1/10體積3 mol/L pH 5.4的醋酸鈉,再加1體積預冷的異丙醇,置-20℃冰箱30min。11000r/min、4℃離心10min,沉淀懸浮于500μL無RNase的去離子水配制的70%乙醇。11000r/min、4℃離心10min,棄去上清液,沉淀干燥后溶于50μL無RNase的去離子水中。
將提取到的總RNA在使用和保存之前進行檢測,使用分光光度計(Thermo BioMate 3,美國),根據吸光度值測定所提總RNA的純度(A260/A280的值為1.8~2.0,說明RNA無污染)及濃度;并依照測定出的RNA濃度確定下一步實驗中合成cDNA所需模板的用量,未使用的RNA保存于-80℃超低溫冰箱;采用非變性瓊脂糖凝膠電泳方法判斷總RNA的完整性。
1.4 RT-PCR擴增
按照試劑盒說明書,以當歸葉片總RNA合成cDNA,然后以cDNA為模板進行PCR反應。反應體系:在200μL PCR管中加入下列組分,無菌去離子水35.8μL、MgCl2(25mmol/L)5μL、10×PCR緩沖液5μL、dNTP(2mmol/L)1.2μL、正、反向引物(10μmol/L)1μL、cDNA 0.5μL和Taq DNA polymerase(5U/μL)0.5μL,總體積為50μL。反應程序:95℃預變性3min;95℃變性30s、56℃退火30s、72℃延伸60s,30個循環;最后72℃延伸10min,4℃結束,PCR擴增產物用1.5%瓊脂糖凝膠電泳,用凝膠-化學發光成像系統(Bio-Rad CHEMI DOC XRS,美國)檢測。目的片段回收和純化按照回收試劑盒說明書進行。
1.5 陽性克隆的篩選、鑒定及測序
DNA片段與T載體連接體系包括:T4DNA連接酶的2×連接緩沖液5μL、pGEM-T Easy載體0.5μL、純化的PCR產物1.5μL(值以PCR產物∶載體摩爾比1∶1估算)、T4DNA連接酶0.8μL,無菌去離子水補充至10μL,4℃連接過夜。連接產物全部加入到100μL置于冰上的感受態大腸桿菌DH5α細胞中,冰浴20min;42℃熱激50s;冰浴2min。加入900μL平衡至室溫的液體LB培養基,搖菌1.5h(37℃,150r/min)。取100 μL菌液涂布于含有氨芐青霉素的LB固體培養基上,37℃培養過夜。次日挑選單菌落,進行菌落PCR檢測,陽性克隆接種于含氨芐青霉素的液體培養基培養(37℃,150r/min),培養過夜后裝入菌種保藏管送生工生物工程(上海)有限公司測序。
1.6 序列的生物信息學分析
Blast搜索在NCBI網站上進行,序列的比對、翻譯和作圖等在DNAMAN 6.0生物軟件上進行,序列保守區分析在NCBI網站Conserved Domain Database(CDD)中進行。具體分析參照已經發表的關于基因克隆及序列分析文獻中方法進行[20-22]。
1.7 PAL在當歸植株組織中的特異表達
采用實時熒光定量PCR(Bio-Rad CFX96,美國)測定PAL在當歸植株組織中的特異表達。分別稱取約0.1 g當歸根、莖、葉組織提取總RNA,每個組織重復3次,然后用分光光度計測定濃度,計算并量取總RNA約100 ng。按試劑盒說明書進行cDNA的第一鏈合成,將cDNA用無菌去離子水稀釋至30μL后取1μL cDNA作為熒光定量PCR模板進行熒光定量PCR,每個cDNA模板做3管重復。反應體系:在200μL PCR管中加入無菌去離子水7.4μL、2×qPCR mix 10μL、正、反向引物(10μmol/L)0.8μL、cDNA 1μL,總體積為20μL。反應程序:95℃預變性3min;95℃變性15s、60℃退火30s、72℃延伸30s,40個循環。熒光采集時間在72℃延伸步驟,擴增完成后利用PCR儀自帶的程序進行熔點曲線測定。求取3個平行管的平均Ct值,采用2-△△Ct分析方法對PAL進行相對定量表達分析。
1.8 統計與分析
所有數據采用SPSS 16.0進行統計和分析。根、莖、葉間PAL相對表達量差異性利用ANOVA分析,采用LSD檢驗。
2 結果與分析
2.1 總RNA的提取及檢測
采用非變性瓊脂糖凝膠電泳法對當歸不同組織總RNA進行檢測,結果顯示,28SrRNA和18SrRNA條帶清晰,沒有其他雜質條帶(圖1),說明本試驗提取的葉片總RNA的完整性較好;經分光光度計測定A260/A280平均值為1.98,表明總RNA的純度較高,可用于RT-PCR擴增。
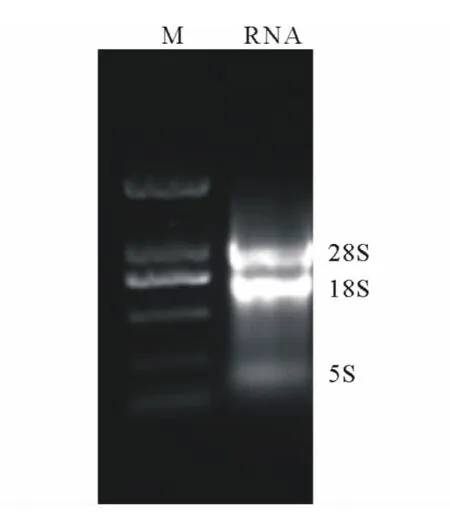
圖1 葉片總RNA非變性瓊脂糖凝膠電泳Fig.1 Nondenaturing agrose gel electrophoresis of total RNA from leaves
2.2 RT-PCR擴增
以當歸葉片總RNA反轉錄所得到的第一鏈cDNA為模板,用PAL擴增引物P1和P2進行PCR擴增。經檢測,擴增產物約在700bp處有1條亮帶(圖2),與理論目的片段大小一致,推測為PAL片段,需要進一步測序鑒定。
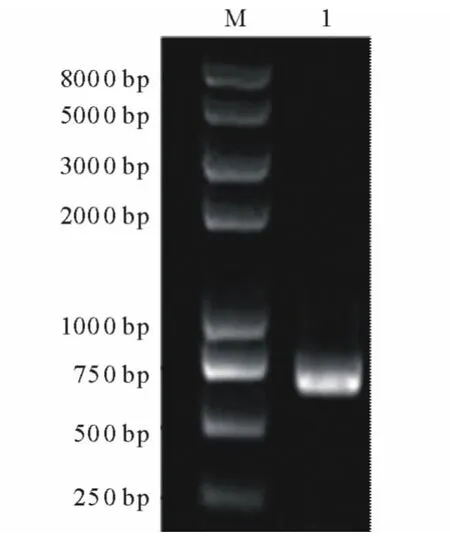
圖2 RT-PCR產物瓊脂糖凝膠電泳Fig.2 Agrose gel electrophoresis of RT-PCR products
2.3 陽性克隆的篩選、鑒定及測序
將回收純化的目的片段連接到pGEM-T Easy克隆載體上,轉化大腸桿菌DH5α,從轉化的平板上隨機挑取5個克隆并進行菌落PCR擴增,經檢測1號和2號克隆擴增出大小約為700bp的條帶(圖3),與RT-PCR結果一致,可能為陽性克隆,然后將陽性克隆進行菌液培養并進行測序,測得一段長度為706bp的序列,將該基因序列在NCBI網站的GenBank數據庫中注冊 (登錄號:KJ000258)。
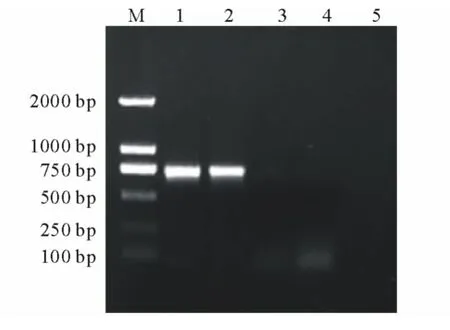
圖3 陽性克隆的PCR鑒定Fig.3 PCR identification of positive clones
2.4 序列分析
對克隆得到的基因片段進行序列分析,其編碼232個氨基酸(圖4)。Blast比對結果顯示,該序列與NCBI基因庫中6個物種的PAL核苷酸序列的相似性在80%以上,與珊瑚菜(Glehnialittoralis)GlPALB基因的相似性最高,為97%,與朝鮮當歸PAL的相似性為88% (表1),表明本研究克隆到的基因片段為當歸PAL片段,將其命名為AsPAL。將推測的當歸PAL片段氨基酸序列和其他植物PAL氨基酸序列進行多重比較(圖5),結果發現完全一致的氨基酸多達179個,占總氨基酸數的77%,與珊瑚菜、皺波天竺葵、胡蘿卜、朝鮮當歸、野山茶(Camelliasinensis)和金銀花氨基酸序列相似性分別達99.14%,98.71%,97.42%,95.28%,86.70%和84.98%,表明克隆片段所編碼的肽段為PAL保守區域,并將其命名為As-PAL。進一步對AsPAL進行保守結構域分析,結果顯示,其包含裂解酶I類超家族 (Lyase_like Superfamily)保 守 結 構 域 (1-44)、PLN02457 結 構 域 (1-223)、TIGR01226結構域(1-224)和pfam00221結構域(1-71),它們均是苯丙氨酸解氨酶的相關結構域,說明AsPAL為當歸PAL的功能區域。
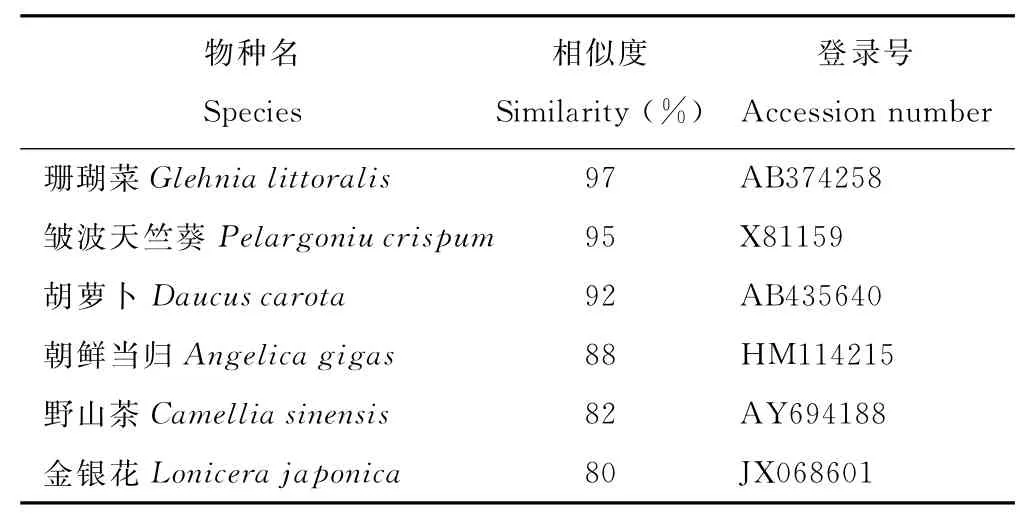
表1 當歸與部分植物PAL片段核苷酸序列相似性比對Table 1 Similarity analysis of PALfragment nucleic acid sequences between A.sinensis and other plant species
2.5 PAL在當歸植株組織中的特異表達
由總RNA的非變性瓊脂糖凝膠電泳結果看出,當歸根、莖及葉組織中總RNA的28S條帶和18S條帶亮度相當(圖6),說明反轉錄總RNA濃度基本相同。實時熒光定量PCR擴增曲線顯示,Actin和PAL的擴增效果良好,Ct值在21~28之間循環;熔點曲線分析也表明擴增曲線具有良好的特異性,表現為單一峰,沒有非特異性熒光峰,Actin和PAL的熔點曲線峰值分別出現在86℃和85℃。將熒光定量PCR的產物進行瓊脂糖凝膠電泳檢測,結果顯示擴增條帶大小約為110bp,與目的大小一致,無引物二聚體條帶(圖7)。
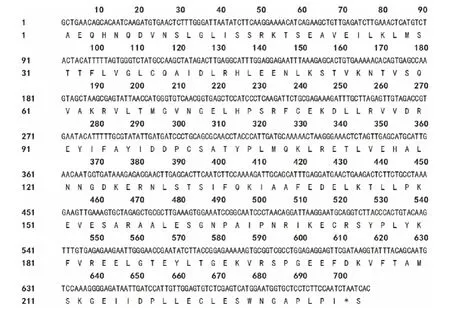
圖4 當歸PAL片段的核苷酸序列及推測的氨基酸序列Fig.4 Nucleic acid sequence and deduced amino acid sequence of PAL fragment fromA.sinensis

圖5 當歸與部分植物PAL片段氨基酸序列相似性多重比較Fig.5 Multiple comparisons on amino acid sequence of PALfragment between A.sinensis and other plant species
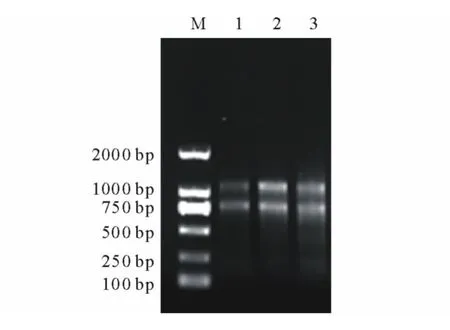
圖6 當歸根、莖、葉總RNA非變性瓊脂糖凝膠電泳Fig.6 Nondenaturing agarose gel electrophoresis of total RNA from root,stem and leaf of A.sinensis
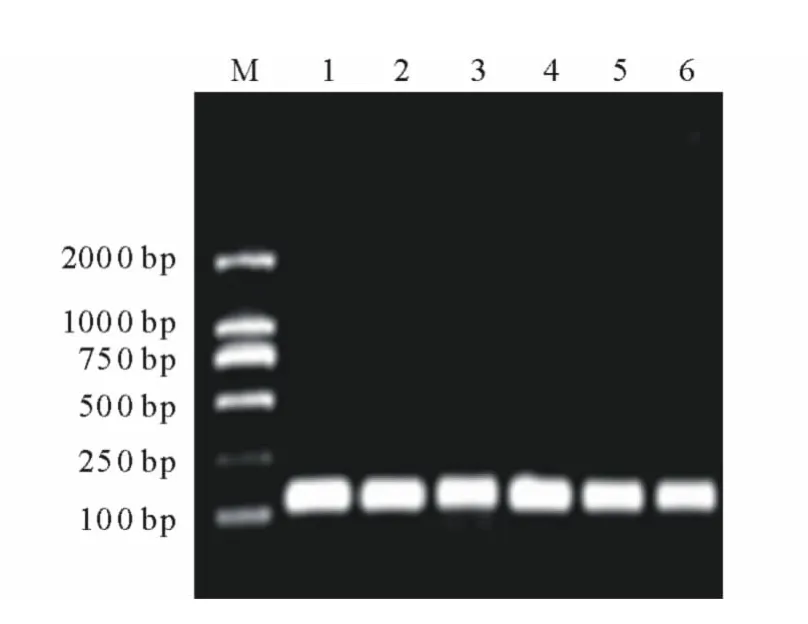
圖7 熒光定量PCR產物非變性瓊脂糖凝膠電泳Fig.7 Nondenaturing agarose gel electrophoresis of products of fluorescence quantitative RT-PCR
采用2-△△Ct方法分析當歸根、莖、葉組織中PAL相對表達量,結果顯示PAL在各組織中均有表達,葉片中表達量最高,其次是莖和根,葉片和莖中表達量分別是根表達量的7.5和2.7倍,三者間差異性顯著(P<0.05)(圖8)。
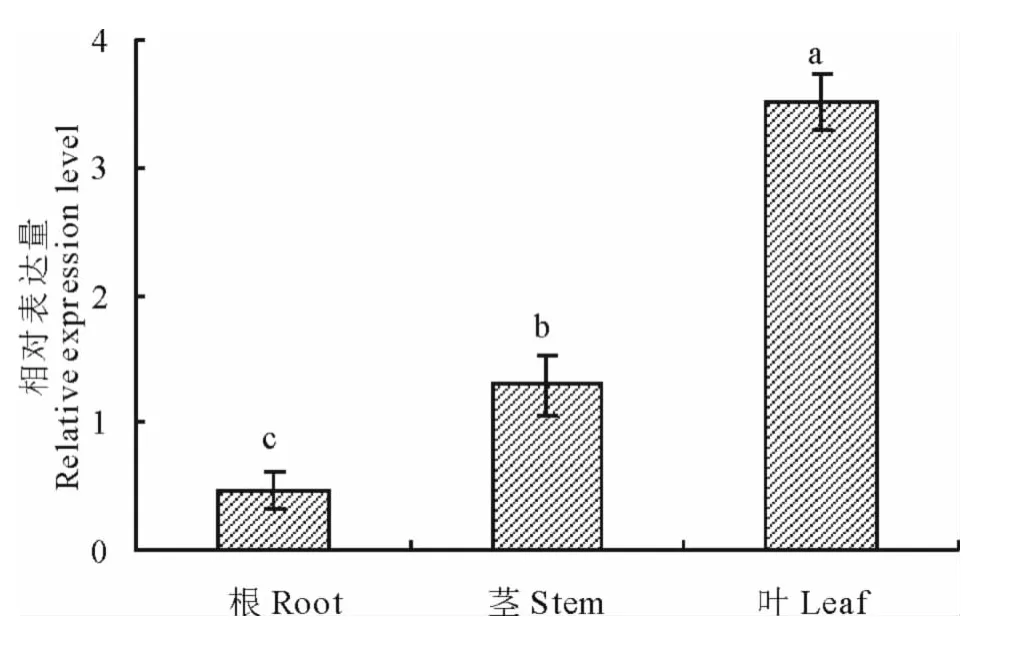
圖8 PAL在當歸根、莖和葉中的相對表達量Fig.8 Relative expression levels of PALin root,stem and leaf of A.sinensis
3 討論
苯丙氨酸解氨酶(PAL)催化的反應是苯丙烷類代謝的第一步反應,其將初生代謝產物苯丙氨酸納入到次生代謝中,成為很多次生代謝產物的前體物質。該酶分布比較廣泛,在所有綠色植物、真菌、細菌、酵母以及藻類中都有[23]。本研究雖然僅擴增的是當歸PAL3′端的保守區片段,但經與GenBank中的其他物種PAL序列比對發現,AsPAL與朝鮮當歸及皺波天竺葵PAL序列同源性非常高,相似性達92%。而與珊瑚菜、皺波天竺葵、胡蘿卜、朝鮮當歸、野山茶和金銀花的同源性也達80%以上,初步推斷AsPAL可能是當歸PAL酶的cDNA片段。由AsPAL保守結構域分析結果初步證實該序列為PAL的保守區域,可能包含著裂解酶I類超家族(Lyase_I_like Superfamily)保守結構域序列。該超家族有裂解酶I家族、組氨酸裂解酶和苯丙氨酸裂解酶,它們催化類似的β消除反應。這些分析結果預示著AsPAL可能是高等植物PAL家族的新成員,它具有較高的結構和功能保守性,這為進一步從基因的組成成分、理化特性、亞細胞定位、功能結構域等方面更全面而準確地揭示當歸阿魏酸生物合成的分子機理奠定了基礎。
苯丙氨酸解氨酶在調控代謝途徑方面具有復雜的作用,Howles等[24]認為,在轉基因煙草(Nicotiana tabacum)中PAL過量表達可提高綠原酸等相關次生代謝產物的含量,而Sewalt等[25]的研究表明,PAL表達抑制的轉基因煙草中木質素等相關次生代謝產物含量會減少。本研究首次從當歸植株中克隆了PAL,通過非變性瓊脂糖凝膠電泳分析,PAL在當歸葉組織中表達量顯著高于根組織,預示著當歸中阿魏酸的生物合成可能是先由葉組織合成,然后由植物傳導系統將其輸送至根部積累。
本研究曾比對朝鮮當歸及皺波天竺葵PAL序列設計簡并引物并進行當歸PAL全長的擴增,但未能獲得有效的擴增結果,這是否與當歸PAL在5′端具有自身特異性有關,還需進一步研究。本研究成果為進一步克隆方法獲得當歸PAL全長,以及對闡明其在當歸阿魏酸生物合成途徑中的功能奠定了工作基礎,同時也對未來通過基因工程手段調控當歸阿魏酸含量提供理論依據。
[1] 國家藥典委員會.中華人民共和國藥典[S].北京:中國醫藥科技出版社,2010:124-125.
[2] 張新慧,張恩和,王惠珍,等.連作對當歸生長的障礙效應及機制研究[J].中國中藥雜志,2010,35(10):1231-1234.
[3] 王惠珍,張新慧,李應東,等.輪作與連作當歸光合特性和揮發油的比較[J].草業學報,2011,20(1):69-74.
[4] 王田濤,王琦,王惠珍,等.連作條件下間作模式對當歸生長特性和產量的影響[J].草業學報,2013,22(2):54-61.
[5] 邱黛玉,藺海明,方子森,等.種苗大小對當歸成藥期早期抽薹和生理變化的影響[J].草業學報,2010,19(6):100-105.
[6] 王婕,趙建邦,宋平順,等.30批當歸中阿魏酸、藁本內酯含量測定[J].中國實驗方劑學雜志,2011,17(16):70-73.
[7] 歐仕益,包惠燕,藍志東.阿魏酸及其衍生物的藥理作用研究進展[J].中藥材,2011,24(3):220-221.
[8] 劉敬,李文建,王春明,等.當歸中有效成分阿魏酸的生物合成及調控[J].中草藥,2008,39(12):1909-1912.
[9] 章霄云,郭安平,賀立卡,等.木質素生物合成及其基因調控的研究進展[J].分子植物育種,2006,4(3):431-437.
[10] Navarre D A,Payyavula R S,Shakya R,etal.Changes in potato phenylpropanoid metabolism during tuber development[J].Plant Physiology and Biochemistry,2013,65:89-101.
[11] Park N I,Park J H,Park S U.Overexpression of cinnamate 4-hydroxylase gene enhances biosynthesis of decursinol angelate inAngelicagigashairy roots[J].Molecular Biotechnology,2012,50(2):114-120.
[12] Appert C,Logemann E,Hahlbrock K,etal.Structural and catalytic properties of the four phenylalanine ammonia-lyase isoenzymes from parsley(PetroselinumcrispumNym.)[J].European Journal of Biochemistry,1994,225(1):491-499.
[13] Yuan Y,Wang Z,Jiang C,etal.Exploiting genes and functional diversity of chlorogenic acid and luteolin biosyntheses inLonicerajaponicaand their substitutes[J].Gene,2014,534:408-416.
[14] Xu H,Park N I,Li X,etal.Molecular cloning and characterization of phenylalanine ammonia-lyase,cinnamate 4-hydroxylase and genes involved in flavone biosynthesis inScutellariabaicalensis[J].Bioresource Technology,2010,101(24):9715-9722.
[15] Yazaki K,Kataoka M,Honda G,etal.cDNA cloning and gene expression of phenylalanine ammonia-lyase inLithospermum erythrorhizon[J].Bioscience,Biotechnology,and Biochemistry,1997,61(12):1995-2003.
[16] Liu R,Xu S,Li J,etal.Expression profile of a PAL gene fromAstragalusmembranaceusvar.Mongholicus and its crucial role in flux into flavonoid biosynthesis[J].Plant Cell Reports,2006,25(7):705-710.
[17] Hu Y S,Zhang L,Di P,etal.Cloning and induction of phenylalanine ammonia-lyase gene fromSalviamiltiorrhizaand its effect on hydrophilic phenolic acids levels[J].Chinese Journal of Natural Medicines,2009,7(6):449-457.
[18] 吳永娜,胡靜,王引權,等.當歸肌動蛋白基因片段的克隆及序列分析[J].中草藥,2012,43(12):2485-2489.
[19] Pandit S S,Mitra S S,Giri A P,etal.A quick method for isolating RNA from raw and ripe fleshy fruits as well as for coisolating DNA and RNA from polysaccharide and polyphenol-rich leaf tissues[J].Journal of Plant Biology,2007,50(1):60-64.
[20] 孔令芳,張吉宇,劉志鵬,等.無芒隱子草SAMS1基因的克隆及干旱脅迫下的表達分析[J].草業學報,2013,22(1):268-275.
[21] 李劍,張金林,王鎖民,等.小花堿茅HKT2;1基因全長cDNA的克隆與生物信息學分析[J].草業學報,2013,22(2):140-149.
[22] 任愛琴,易津,高洪文,等.檸條錦雞兒CkNCED1基因啟動子的克隆及表達分析[J].草業學報,2013,22(2):165-170.
[23] 姜翠翠,陳桂信,潘東明,等.油木奈果實苯丙氨酸解氨酶基因的分離與表達分析[J].西北植物學報,2013,33(3):465-471.
[24] Howles P A,Sewalt V J H,Paiva N L,etal.Overexpression of L-phenylalanine ammonia-lyase in transgenic tobacco plants reveals control points for flux into phenylpropanoid biosynthesis[J].Plant Physiology,1996,112(4):1617-1624.
[25] Sewalt V J H,Ni W,Blount J W,etal.Reduced lignin content and altered lignin composition in transgenic tobacco downregulated in expression ofL-phenylalanine ammonia-lyase or cinnamate 4-hydroxylase[J].Plant Physiology,1997,115(1):41-50.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
