PAX6基因多態性與精神分裂癥的關系探討
2014-06-14 09:48:02韓海斌楊建立溫子玉
山東醫藥 2014年12期
韓海斌,楊建立,丁 玉,溫子玉
(1天津醫科大學,天津300070;2天津市精神衛生中心)
精神分裂癥是一種常見的病因尚未完全闡明的精神病,其發生可能與神經發育異常有關[1~3]。目前認為,遺傳因素和某些神經發育危險因素的相互作用,使胚胎期大腦發育過程出現某種神經病理變化,是精神分裂癥神經發育異常的機制之一。PAX6作為PAX基因家族中的一員,定位于人染色體第11號染色體長臂13位點(11p13)。大腦皮層發育期間PAX6缺陷導致成熟期認知缺陷,發育階段PAX6缺失導致皮層、皮層間及皮層內連接嚴重缺損,這可能成為鼠及人類出生后認知缺損的基礎[4]。相關研究表明,PAX6基因多態性可能通過調節發育中或成年的大腦而參與偏執型精神分裂癥的發生[5]。本研究采用病例對照研究方法,探討PAX6基因多態性與精神分裂癥的關系。
1 資料與方法
1.1 臨床資料 患者組:2011~2013年在天津市精神衛生中心治療的首發精神分裂癥患者270例,均為漢族,年齡18~45歲。診斷由2名以上精神科主治醫師獨立完成,符合DSM-IV關于精神分裂癥的診斷標準。排除煙酒及其他精神活性物質濫用者、有其他精神障礙家族史者、有肝腎嚴重軀體疾病者。患者入組前詳細了解本研究目的和程序,并簽署知情同意書。對照組:健康志愿者320例,均為漢族,年齡、性別與患者組具有可比性;一般健康問卷各項目總分<30分;經非結構式臨床訪談排除精神疾病,且家族中無精神疾病和自殺者,個體間無血緣關系;在詳細了解本研究目的和程序后,均簽署知情同意書。
1.2 方法
1.2.1 標本采集與處理 以EDTA作為抗凝劑,取肘靜脈血10 mL充分抗凝處理,冷藏保存,離心后-80℃冷凍。
1.2.2 基因多態性檢測 單核苷酸多態性(SNP)位點:通過HapMap數據庫檢索PAX6基因34 kb區域(覆蓋上游8 kb和下游4 kb),選取其中位于PAX6 的3'非翻譯區(3'-UTR)的 rs3026401、rs1506兩個標簽SNP位點。采用連接酶檢測反應研究SNP的多態性。
采用AXYGEN公司的AxyPrep-96全血基因組DNA試劑盒提取人類基因組DNA。檢測總DNA濃度、純度、完整性,擴增 SNP位點所在片段,準備PCR Master Mix(20 μL體系)。反應條件:95℃變性15 min,94 ℃ 30 s、56 ℃ 1 min、72 ℃ 1 min(35個循環),72℃延伸7 min。反應結束后,取2 μL反應產物在3.0%瓊脂糖凝膠、0.5×TBE中電泳。剩余樣品保存于-20℃。在PCR擴增產物中加入等體積ddH2O稀釋,作為連接反應的模板。連接反應體系(10 μL):在 1.5 mL eppendorf離心管中分別加入 100 μL buffer(10 × )、100 μL Probe Mix、5 μL 連接酶、695 μL去離子水。充分混勻后離心,再取9 μL分裝在200 μL的 PCR反應管中,最后再加入1 μL PCR反應產物。在Perkin-Elmer Gene Amp PCR Systems9600上設置如下程序:95℃變性2 min,94℃ 30 s、50 ℃ 2 min(35 個循環)。取1 μL LDR 連接產物、1 μL ABI GS-500 ROX熒光標記分子量標準、1 μL去離子甲酰胺上樣液混合,95℃加熱變性2 min,冰中驟冷;于5%聚丙烯酰胺和5 mol/L尿素中3 000 V電泳2.5 h,應用Genescantm672軟件進行數據收集、泳道線校正、遷移片段大小測量和校正內在分子量標準,應用Genemapper軟件進行數據分析和基因分型。
1.2.3 臨床癥狀評分比較 治療前采用陽性與陰性癥狀量表(PANSS)評定陽性癥狀、陰性癥狀及一般精神病理因子情況,對患者組SNP rs3026401不同基因型患者的臨床癥狀評分進行比較。
1.2.4 統計學方法 采用SPSS13.0統計軟件。組間基因型和等位基因分布頻率比較采用χ2檢驗,rs3026401各基因型PASS評分比較采用方差分析;使用 SHEsis遺傳分析軟件[6]進行 Hardy-Weinberg平衡、連鎖不平衡(LD)檢驗及單位點和單體型遺傳關聯分析。P≤0.05為差異有統計學意義。
2 結果
2.1 樣本情況 因為檢測過程中樣本損失,患者組8例、對照組2例的rs3026401和(或)rs1506位點數據缺失,故在統計中將其剔除。最終患者組262例、對照組318例納入本研究。
2.2 基因型及等位基因分析結果
2.2.1 Hardy-Weinberg 平衡檢驗結果 兩組的SNP rs3026401基因分布均符合Hardy-Weinberg平衡定律,χ2=0.42、0.37,P 均 >0.05;SNP rs1506 基因分布也均符合 Hardy-Weinberg平衡定律,χ2=2.20、0.08,P 均 >0.05;表明研究樣本具有群體代表性。
2.2.2 Genemapper數據分析結果 患者組各位點的分型截圖見圖1。
2.2.3 兩組 PAX6 SNP rs1506、rs3026401 基因型、等位基因分布情況 患者組SNP rs1506基因型、等位基因分布與對照組比較,差異均無統計學意義(χ2=2.877、1.187,P 均 >0.05);患者組 SNP rs3026401 基因型、等位基因分布與對照組比較,差異均有統計學意義(χ2=11.635、11.044,P 均 <0.05)。見表1、2。
2.2.4 SNP rs1506、rs3026401的LD 分析結果 分析結果顯示,D'=1、r2=0.812,表明這兩個位點之間存在很強的LD。對這兩個位點構成的單倍型進行分析,結果顯示患者組和對照組之間單倍型rs3026401A/rs1506T、rs3026401G/rs1506T 頻率差異具有統計學意義(P均<0.05)。
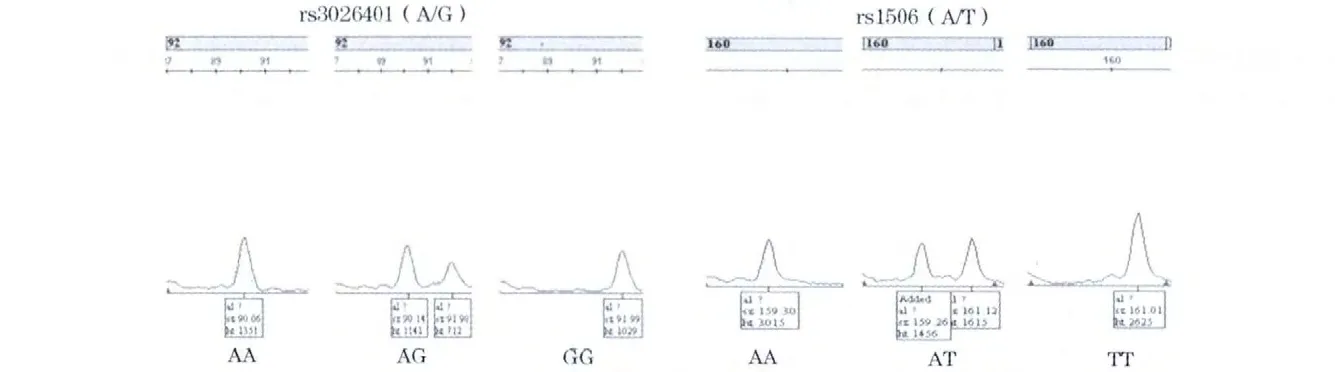
圖1 Genemapper數據分析患者組各位點的分型截圖
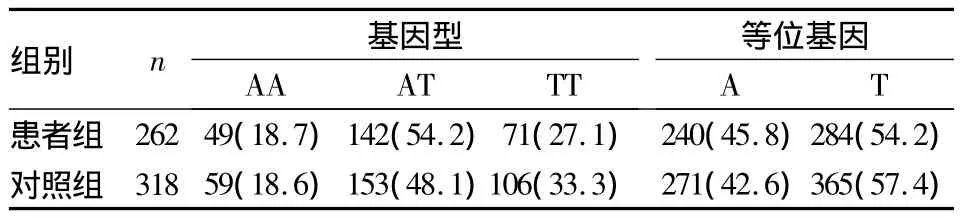
表1 兩組SNP rs1506基因型、等位基因分布[例(%)]
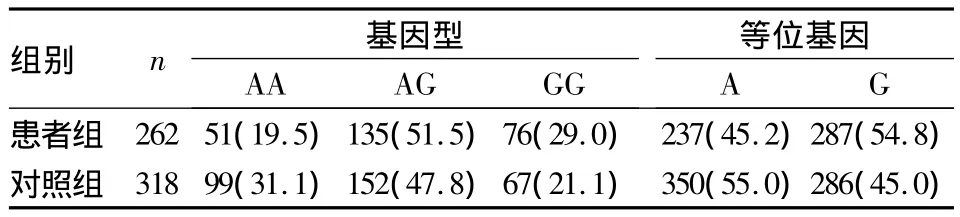
表2 兩組SNP rs3026401基因型、等位基因分布[例(%)]
2.2.5 患者組SNP rs3026401不同基因型患者的臨床癥狀評分比較 患者組SNP rs3026401不同基因型的陽性癥狀評分、一般精神病理因子評分差異均無統計學意義(P均>0.05),而陰性癥狀評分差異具有統計學意義(P<0.05),即 GG>AA、GG>AG(P均 <0.05)。見表3。
表3 患者組SNP rs3026401不同基因型患者的PANSS 評分比較(±s)
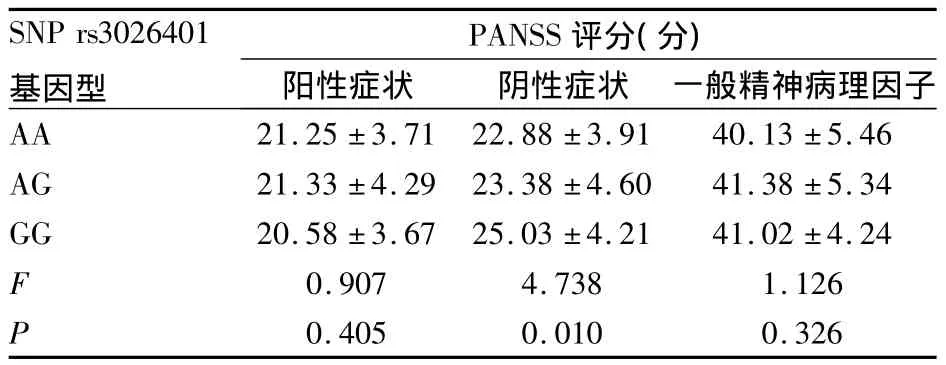
表3 患者組SNP rs3026401不同基因型患者的PANSS 評分比較(±s)
SNP rs3026401基因型PANSS評分(分)陽性癥狀 陰性癥狀 一般精神病理因子AA 21.25±3.71 22.88±3.91 40.13±5.46 AG 21.33±4.29 23.38±4.60 41.38±5.34 GG 20.58±3.67 25.03±4.21 41.02±4.24 F 0.907 4.738 1.126 P 0.405 0.010 0.326
3 討論
精神分裂癥多起病于青壯年,常緩慢起病,病程遷延,有慢性化傾向和衰退的可能。國內外相繼進行的大規模流行病學調查顯示,精神分裂癥患病率為1.4‰ ~4.6‰,發病率為0.16‰ ~0.43‰[7]。精神分裂癥的病因至今尚未明確,現有治療方法效果均不理想[8]。
人與鼠的PAX6基因具有高度同源性。動物實驗表明,PAX6基因在發育過程中的腦和成體腦中都有表達,且對腦發育和腦結構有重要作用[9,10]。此外,有研究表明PAX6參與出生后海馬神經發生的祖細胞的形成和維持[11],而海馬的發生又與精神分裂癥關系密切[12]。動物學研究還發現,PAX6抑制鼠類星形膠質細胞增殖,促進其成熟[13];PAX6雜合鼠(PAX6-/+)表現出沒有松果體及單邊多小腦回等[14]。有研究表明,PAX6基因突變能夠引起人腦結構異常[15]。精神分裂癥的神經發育異常假說認為,由于遺傳因素和某些神經發育危險因素的相互作用,在胚胎期大腦發育過程就出現了某種神經病理變化。研究發現,精神分裂癥患者海馬、額皮層、扣帶回和內嗅皮層有細胞結構紊亂,在這些變化中不伴有膠質細胞增生,可能是由腦發育階段神經元移行異位或分化障礙造成的,破壞了皮層聯絡的正常模式[16]。近1個世紀以來,大量的關于精神分裂癥的家系調查[17]、雙生子研究[18]、寄養子研究[19]、遺傳流行病學研究等結果強烈提示,精神分裂癥與遺傳因素密切相關。現有研究顯示,精神分裂癥并不遵循孟德爾遺傳規律,它更可能是一類由多個、有較小影響和外顯率、相互作用的潛在基因共同作用的結果。由此可見,尋找并定位與精神分裂癥發病有較大關聯的基因位點非常重要。很多研究者致力于尋找與精神分裂癥相關聯的基因多態性的研究,諸如代謝型谷氨酸受體-3基因[20]、精神分裂癥斷裂基因1(DISC1)[21]等的研究。
前脈沖抑制模型(PPI)是研究精神分裂癥的一個重要模型。有關研究顯示,PAX6雜合鼠的PPI明顯受損,同時伴有出生后的神經發育異常[22]。大腦皮層發育期間PAX6缺陷導致成熟期認知缺陷,發育階段PAX6缺失導致皮層、皮層間及皮層內連接嚴重缺損,這成為鼠及人類出生后認知缺損的基礎[4]。作為早期表達的PAX6基因多態性,可能通過調節發育中或成年的大腦而參與偏執型精神分裂癥的發生[5]。同時也有報道稱,PAX6促進精神分裂癥可能是通過破壞維甲酸的信號途徑[23]。mRNA的3'-UTR作為mRNA的一部分,具有miRNA、siRNA的靶位點,miRNA或siRNA結合靶位點后可引起mRNA的降解或抑制翻譯起始,其調控自身mRNA的功能已為人們所熟知。本研究通過檢索HapMap數據庫在PAX6基因3'-UTR區選取其中兩個標簽 SNP rs3026401、rs1506 位點[24],既往也有這兩個位點在其他神經系統疾病中的相關研究[25]。本研究發現,患者組SNP rs1506基因型、等位基因頻率分布頻率與對照組相比差異無統計學意義,而SNP rs3026401基因型、等位基因分布頻率與精神分裂癥均相關,提示PAX6基因rs3026401多態性可能是精神分裂癥的危險因素之一。我們進一步比較了SNP rs3026401不同基因型的PANSS評分情況,結果顯示陽性癥狀評分、一般病理因子評分差異無統計學意義,而陰性癥狀評分存在統計學差異,其中GG型陰性癥狀評分顯著高于AA、AG型,這可能是PAX6基因rs3026401多態性引起精神分裂癥易感的重要原因;但目前國內外相關文獻對此方面的報道較少,有待進一步研究以驗證。
本研究還存在不足之處,應在標準化診斷分類的基礎上進一步擴大樣本量,通過LD及普通關聯分析的方法在不同人群中重復研究,以進一步明確該位點與精神分裂癥易感性的關系。同時,還應對PAX6基因rs3026401位點在精神分裂癥中的具體功能進行研究,以進一步發展與疾病相關的生物學內表型的分析,這將更有利于深入探討精神分裂癥發生的基礎,為更好的預防和治療精神分裂癥提供理論支持。
[1]Prasad KM,Keshavan MS.Structural cerebral variations as useful endophenotypes in schizophrenia:do they help construct"extended endophenotypes"[J].Schizophr Bull,2008,34(4):774-790.
[2]Shenton ME,Dickey CC,Frumin M,et al.A review of MRI findings in schizophrenia[J].Schizophr Res,2001,49(1-2):1-52.
[3]Kircher TT,Thienel R.Functional brain imaging of symptoms and cognition in schizophrenia[J].Prog Brain Res,2005(150):299-308.
[4]Tuoc TC,Radyushkin K,Tonchev AB,et al.Selective cortical layering abnormalities and behavioral deficits in cortex-specific Pax6 knock-out mice[J].J Neurosci,2009,29(26):8335-8349.
[5]Stober G,Syagailo YV,Okladnova O,et al.Functional PAX-6 gene-linked polymorphic region:potential association with paranoid schizophrenia[J].Biol Psychiatry,1999,45(12):1585-91.
[6]Shi YY,He L.SHEsis,a powerful softw are platform for analyses of linkage disequilibrium,haplotype construc tion,and genetic assoc iation at polymorphismloci[J].Cell Res,2005,15(2):97-981.
[7]Jablensky A.Epidemiology of schizophrenia:the global burden of disease and disability[J].Eur Arch Psychiatry Clin Neurosci,2000,250(6):274-285.
[8]李霞,潘麗,金國東,等.帕利哌酮緩釋膠囊治療青少年精神分裂癥的療效及安全性[J].山東醫藥,2011,15(51):96-97.
[9]Stoykova A,Fritsch R,Walther C,et al.Forebrain patterning defects in Small eye mutant mice[J].Development,1996,122(11):3453-3465.
[10]Engelkamp D,Rashbass P,Seawright A,et al.Role of Pax6 in development of the cerebellar system[J].Development,1999,126(16):3585-3596.
[11]Maekawa M,Takashima N,Arai Y,et al.Pax6 is required for production and maintenance of progenitor cells in postnatal hippocampal neurogenesis[J].Genes Cells,2005,10(10):1001-1014.
[12]Jun H,Mohammed Qasim Hussaini S,Rigby MJ,et al.Functional role of adult hippocampal neurogenesis as a therapeutic strategy for mental disorders[J].Neural Plasticity,2012(2012):1-20.
[13]Sakurai K,Osumi N.The neurogenesis-controlling factor,Pax6,inhibits proliferation and promotes maturation in murine astrocytes[J].J Neurosci,2008,28(18):4604-4612.
[14]Mitchell TN,Free SL,Williamson KA,et al.Polymicrogyria and absence of pineal gland due to PAX6 mutation[J].Ann Neurol,2003,53(5):658-663.
[15]Free SL,Mitchell TN,Williamson KA,et al.Quantitative MR image analysis in subjects with defects in the PAX6 gene[J].Neuroimage,2003,20(4):2281-2290.
[16]Stefan MD,Murray RM.Schizophrenia:developmental disturbance of brain and mind[J].Acta Paediatr Suppl,1997,86(422):112-116.
[17]Bladessarini RJ.Schizophrenia[J].N Engl J Med,1977,297(18):988-995.
[18]Saha S,Chant D,Welham J,et al.A systematic review of the prevalence of schizophrenia[J].PloS Medicine,2005,2(5):413-433.
[19]Macdonald AW,Schulz SC.What we know:findings that every theory of schizophrenia should explain[J].Schizsophrenia Bulletin,2009,35(3):493-508.
[20]潘成英,岳偉華,盧天蘭.代謝型谷氨酸受體-3基因多態性和精神分裂癥臨床癥狀關聯研究[J].中國心理衛生雜志,2009,23(7):528-532.
[21]孫偉,閆俊,王力芳,等.精神分裂癥斷裂基因1(DISC1)多態性與精神分裂癥的關聯研究[J].中國心理衛生雜志,2009,23(8):590-594.
[22]Maekawa M,Takashima N,Matsumata M,et al.Arachidonic acid drives postnatal neurogenesis and elicits a beneficial effect on prepulse inhibition,a biological trait of psychiatric illnesses[J].PLoS One,2009,4(3):1-9.
[23]Lamantia AS.Forebrain induction,retinoic acid,and vulnerability to schizophrenia:insights from molecular and genetic analysis in developing mice[J].Biol Psychiatry,1999,46(1):19-30.
[24]Hammond CJ,Andrew T,Mak YT,et al.A susceptibility locus for myopia in the normal population is linked to the PAX6 gene region on chromosome 11:a genomewide scan of dizygotic twins[J].Am J Hum Genet,2004,75(2):294-304.
[25]Jiang B,Yap MK,Leung KH,et al.PAX6 haplotypes are associated with high myopia in Han chinese[J].PLoS One,2011,6(5):1-10.
猜你喜歡
初中生學習指導·提升版(2023年8期)2023-09-12 10:26:19
保健醫苑(2022年1期)2022-08-30 08:39:40
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
保健醫苑(2022年4期)2022-05-05 06:11:14
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
基層中醫藥(2020年10期)2020-02-13 15:45:52
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
獸醫導刊(2016年6期)2016-05-17 03:50:35
中國衛生標準管理(2015年18期)2016-01-20 09:27:01
