發酵產物對灰綠曲霉β-葡萄糖苷酶活力的影響
2014-07-26 01:20:54許鑫琦任躍明馬素娟龍敏南陳清西
廈門大學學報(自然科學版) 2014年1期
許鑫琦,任躍明,馬素娟,龍敏南,陳清西*
(1.廈門大學生命科學學院,福建 廈門361102;2.國家藥典委員會 北京100061;3.廈門大學能源研究院,福建 廈門361102)
纖維素是自然界中分布最廣、含量最為豐富的一類多糖,是理想的生物質能源,能夠水解天然纖維素的纖維素酶,是一個復雜的多酶體系.一般認為纖維素酶包含3種主要水解酶[1]:外切葡聚糖酶(CBH,EC 3.2.1.91)、內切葡聚糖酶(EG,EC 3.2.1.4)和β-葡萄糖苷酶(BG,EC 3.2.1.21).纖維素被內切酶和外切酶水解成纖維二糖和低聚寡糖后,由BG將以上產物水解產生葡萄糖.BG普遍存在于植物、微生物和哺乳動物的腸道中,微生物中的木霉(Trichodermakoningii)、青霉(Penincilliumaurantiogriseum)、黑曲霉(Aspergillusniger)以 及 米 曲 霉 (Aspergillus oryzae)等真菌來源的BG利用方便,其酶學性質被廣泛地研究,對纖維素酶的應用起到了有利的支撐.BG在纖維素酶體系內含量少,酶活力低,是制約纖維素降解速率的關鍵酶,因此BG決定了纖維素糖化過程的最終效率.Xin等[2]研究發現,在廢紙產乙醇發酵過程中,加入BG后,促進糖化并降低了內切酶和外切酶所受到的反饋抑制,進而加速纖維素的降解和提高乙醇產量,這說明在纖維素利用體系中,增加BG用量的必要性.近些年,BG 的分子生物學研究不斷深入[3-4],包括木霉及米曲霉和黑曲霉等絲狀真菌,以及極端微生物如嗜熱菌[5]來源的BG也越來越受到關注,BG的克隆表達增加了BG的工業來源,有利于解決纖維素發酵產糖過程中所遇到的瓶頸.除了能源,在其他領域,BG一直受到了國內外學者的緊密關注[6],如食品、天然產物提取、醫療以及日用化工[7]等領域BG也得到了有效的應用.
本文從自行篩選的灰綠曲霉菌株(Aspergillus glaucusXC-9)的纖維素發酵液中,分離純化出BG,研究了乙醇等發酵產物對其催化活性的影響.
1 材料與方法
1.1 實驗材料
灰綠曲霉(Aspergillusglaucus)是由實驗室篩選得到的、能夠降解纖維素的曲霉菌株[8].BG由實驗室分離純化.
1.2 實驗方法
BG的分離純化:灰綠曲霉發酵液先用質量分數為30%(NH4)2SO4沉淀除雜蛋白,再用70%(NH4)2SO4沉淀BG,離心收集沉淀,溶于適當體積的0.02mol/L pH 5.0HAc-NaAc緩沖液中.選用Sephadex G-100凝膠進行柱層析,柱床為1.5cm×65cm,洗脫液為0.02 mol/L pH 5.0HAc-NaAc(含0.1moL/L NaCl),流速為1mL/min,自動收集器收集洗脫液,合并活力峰,接著進行Phenyl Sepharose Fast Flow疏水層析,柱床為1.6cm×20.0cm.疏水層析洗脫 A液為含0.4mol/L NaCl的0.1mol/L pH 7.8Tris-HCl緩沖液,B液為0.1 mol/L pH 7.8Tris-HCl緩沖液;高鹽上樣,A 液沖平后,用200mL A液和200mL B液進行線性梯度洗脫,洗脫流速0.5mL/min,自動收集器收集樣品,每管收集5mL.檢測洗脫液的蛋白濃度和酶活力,合并活力峰.進行native-PAGE電泳,考馬斯亮藍染色,檢測確定所制備的BG為電泳單一純.
酶活力測定參考文獻[9]:50μL酶液加入200μL 0.2mol/L pH 3.6HAc-NaAc緩沖液和250μL 1%(質量分數)水楊素底物,60℃水浴45min后,再加入2mL 3,5-二硝基水楊酸試劑(DNS)顯色液終止反應.沸水浴5min后立即冷卻,在酶標儀上測A540吸光值.通過標準曲線查得葡萄糖含量.以產生的還原糖量表示酶活力.酶活力定義為在上述條件下每分鐘水解底物產生1μg葡萄糖所需要的酶量.
發酵產物對BG抑制作用研究:在BG測活力體系里加入不同量的發酵產物(乙醇,乳酸和乙酸)后,按BG測活力方法測定活力,3次重復試驗,以發酵產物濃度對BG剩余活力作圖,考察抑制BG活力的濃度效應.
發酵產物抑制機理測定:在測活力體系里,在一定發酵產物濃度下,改變 BG 質量濃度(0.1,0.2,0.3,0.4和0.5μg/mL),按測活力方法測定 BG 的酶活力,用酶活力(μmol/min)對酶質量濃度(μg/mL)作圖,得到一組直線,根據直線的位置關系判斷各發酵產物抑制機理.
發酵產物抑制類型判斷:向測活力體系中加入不同體積的發酵產物,在每個濃度溶液中改變底物的濃度,測定酶促反應的初速率(v),以1/v對1/[cS]雙倒數作圖,分析比較效應物下BG的表觀米氏常數Km和最大反應速率Vmax,根據雙倒數曲線圖判定發酵產物抑制類型,再以Vmax或Km對發酵產物濃度作圖,測定3個發酵產物的抑制常數KI.
含乙醇的反應體系中BG最適反應溫度的變化及BG分子內源熒光測定:向BG測活體系加入不同量乙醇,在不同溫度下測定酶活力,分析比較乙醇體積分數與BG反應溫度的關系.取一定量酶分別在不同體積分數乙醇溶液(0%、12%、16%、18%、20%,pH 3.6)中60℃水浴15min,冷卻后在VARIAN紫外/可見分光光度計檢測BG內源熒光圖譜,熒光發射峰掃描范圍300~405nm,激發波長為280nm.
2 實驗結果
2.1 發酵產物對酶活力的影響
在BG最適反應條件下(pH 3.6,60℃),向測活體系中加入不同體積的乙酸、乙醇和乳酸,研究不同濃度發酵產物對酶催化水解底物的活力影響.酶剩余活力與之依賴關系結果見圖1.隨著各種產物濃度的增大,酶活力迅速下降.乙醇、乙酸、乳酸對酶活力有不同程度抑制,抑制強度依次為乳酸>乙酸>乙醇,其半抑制濃度 (IC50)分別為0.09,0.34和4.4mol/L.
2.2 發酵產物對酶活力的抑制機理
為判斷以上3種物質對BG的抑制機理是否為可逆抑制,分別在含不同濃度乙酸、乙醇和乳酸的測活力體系中,固定底物水楊素濃度為5.0mmol/L,改變酶的加入量,測定不同濃度發酵產物對酶活力的影響.以酶活力對加入酶量作圖,都得到了一組通過原點的直線(圖2),說明三者對BG的抑制作用都表現為可逆過程,即發酵產物抑制了酶分子的活性,并未減少有活力的酶量,從而降低BG的催化效率.
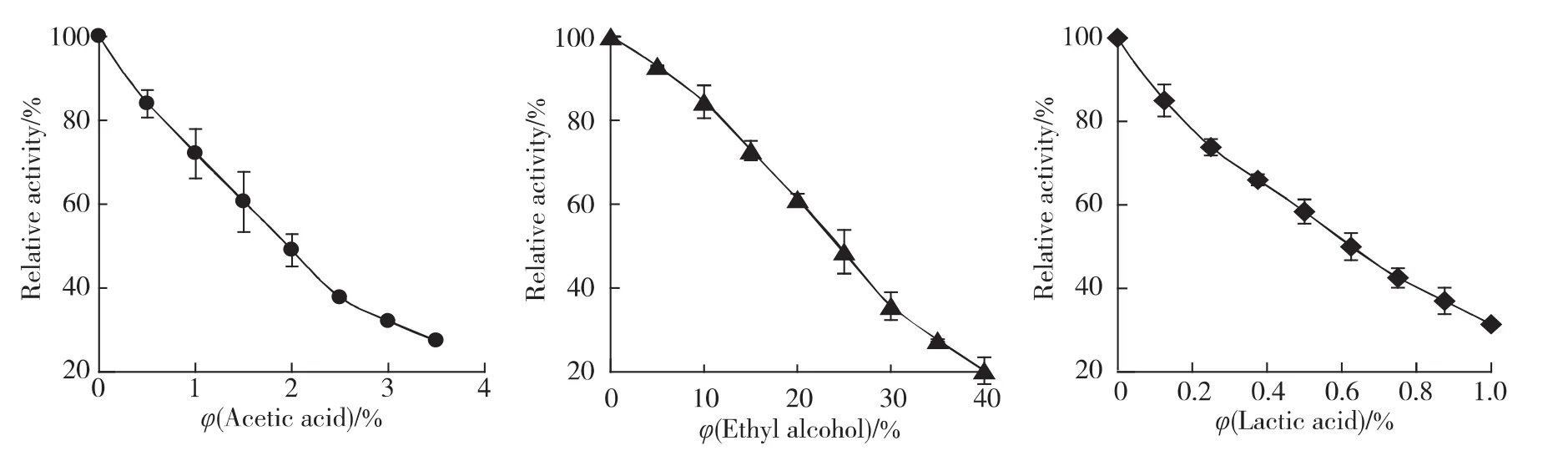
圖1 3種發酵產物對BG活力效應Fig.1 Inhibition of fermention products on BG
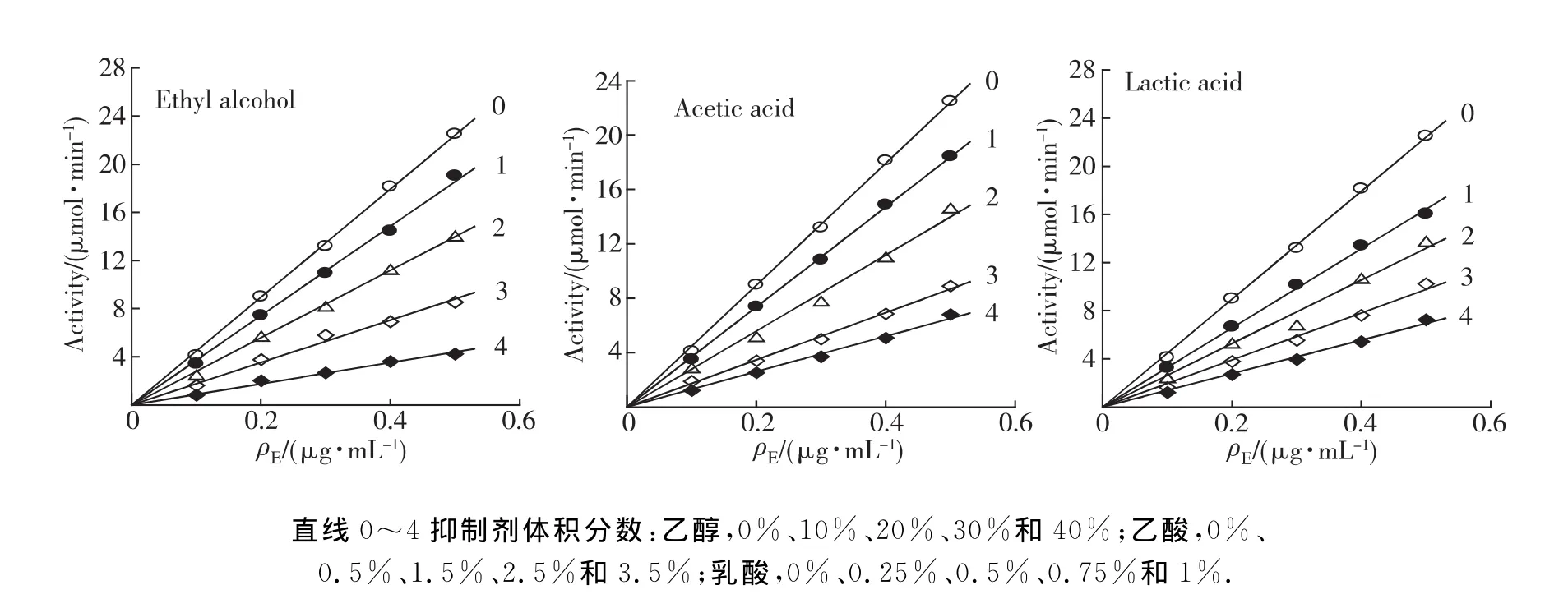
圖2 3種發酵產物對BG活力的影響為可逆抑制Fig.2 Reversible inhibition of three fermention products on BG
在BG最適反應體系中(pH 3.6,60℃),加入不同體積的乙醇、乙酸或乳酸,在每個濃度的溶液中改變底物的濃度,測定酶促反應的初速率(v),以1/v對1/[cS]作圖,結果如圖3~5所示.雙倒數作圖均得到一組相交于橫軸的直線(圖3(a),4(a)和5(a)).說明乙醇、乙酸及乳酸對BG不影響米氏常數(Km),只影響最大反應速率(Vm),抑制機理表現為非競爭性類型,可以同時與游離酶(E)和結合酶(ES)結合,且結合常數相同.以不同濃度效應物下測定的1/Vm對效應物濃度作圖為一條直線,直線的斜率為1/(Vm·KI)(圖3(b),4(b)和5(b)),從而求得發酵產物對BG 的抑制常數分別為乙醇26% (4.25mol/L)、乙酸2% (0.315 mol/L)和乳酸0.7% (0.083mol/L).
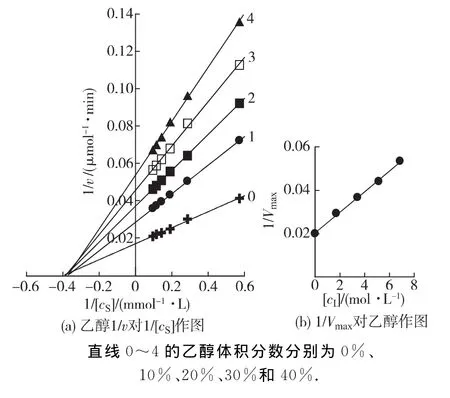
圖3 在不同體積分數乙醇的反應體系中BG水解水楊素Lineweaver-Burk曲線Fig.3 Lineweaver-Burk plots for the hydrolysis of salicin by BG in different concentrations of ethanol
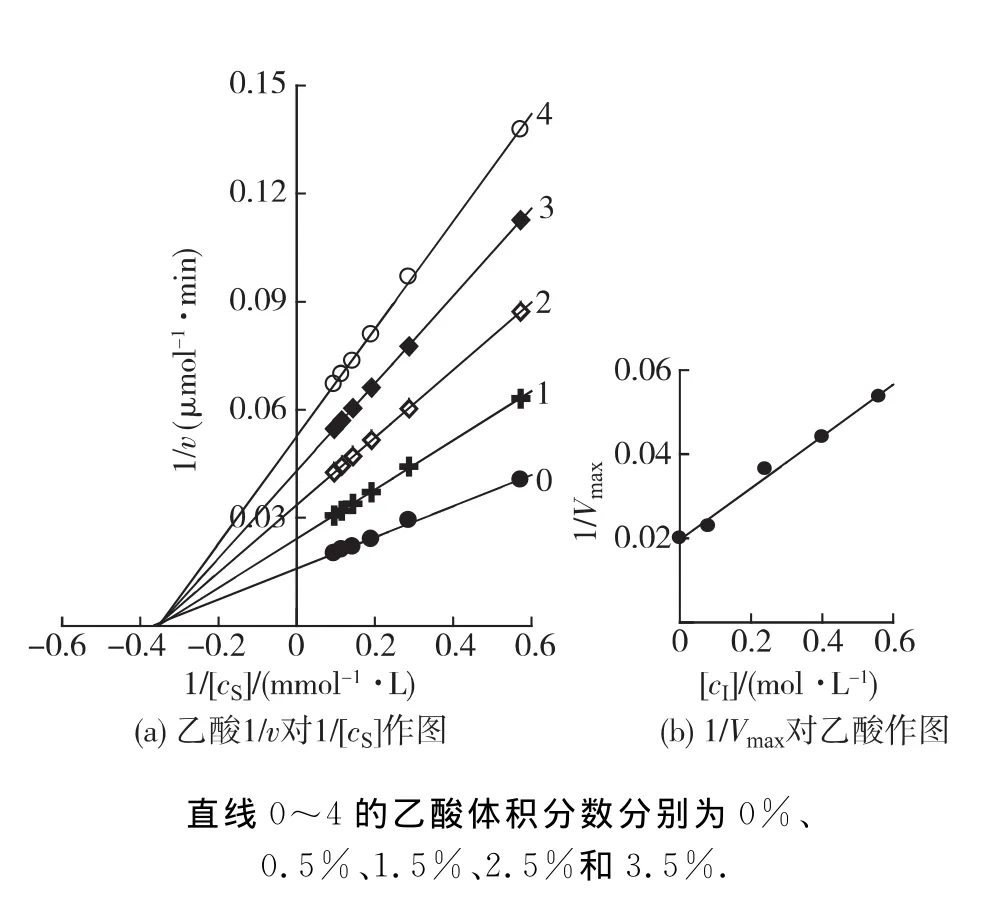
圖4 在不同體積分數乙醇的反應體系中對BG水解水楊素Lineweaver-Burk曲線Fig.4 Lineweaver-Burk plots for the hydrolysis of saliciny BG in different concentrations of acetic acid
2.3 乙醇對BG反應溫度和酶分子內源熒光光譜影響
測定了不同體積分數乙醇對BG催化反應溫度的影響,結果如圖6所示.隨著乙醇濃度的增加,BG催化活力隨之下降的同時,BG水解底物的最適溫度逐漸減小,無乙醇存在下為60℃,當乙醇升至20%(體積分數)時,BG的最適溫度已降到了50℃,說明乙醇體積分數的增加可降低BG最適反應溫度.當反應體系溫度小于55℃,乙醇低于16%時,BG酶活力都高于無乙醇情況下的活力.
最適反應條件(60℃,pH 3.6)保存15min后,BG在不同體積分數乙醇溶液的熒光發射光譜變化結果見圖7.在280nm激發下,天然酶內源熒光在338 nm處有特征發射峰,當乙醇體積分數小于12%時,BG內源熒光強度隨乙醇的增加而下降,未發生紅移;當乙醇體積分數超過12%并繼續增加時,其強度隨著乙醇體積分數上升而增強,當乙醇體積分數達20%時,熒光強度增加了50%,但未發生紅移,酶分子高級結構還未被完全破壞.

圖5 在不體積分數乳酸的反應體系中BG水解水楊素Lineweaver-Burk曲線Fig.5 Lineweaver-Burk plots for the hydrolysis of salicin by BG in different concentrations of lactic acid
3 討 論
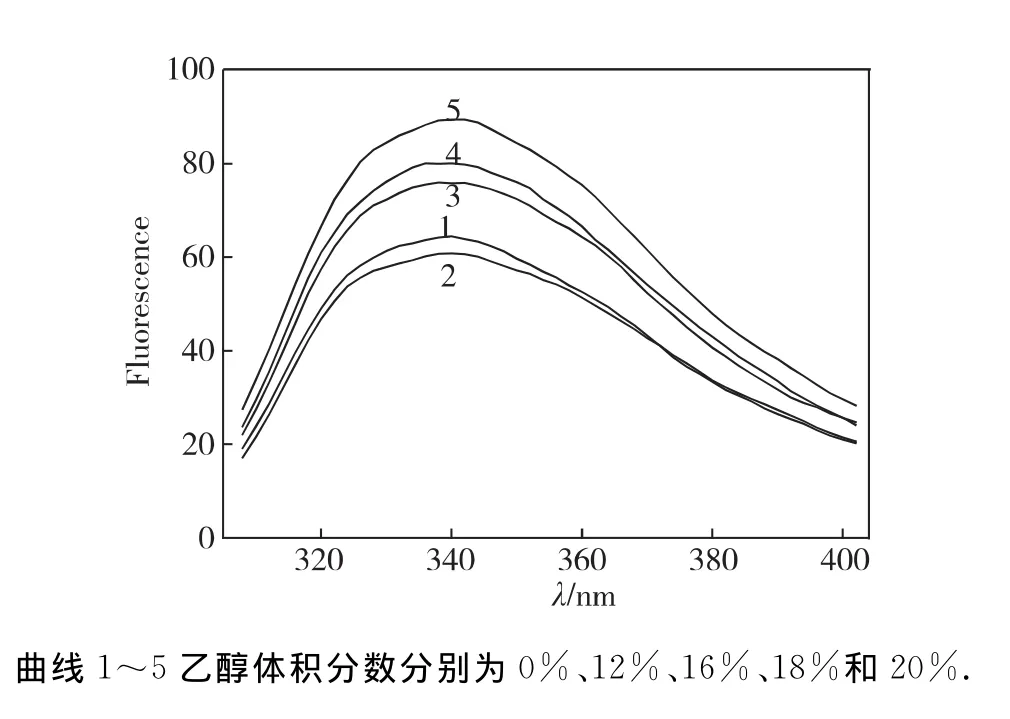
圖7 pH3.6不同體積分數乙醇溶液60℃處理15min后BG熒光發射光譜Fig.7 Fluorescence emission spectra of BG after incubating in different concentration of ethanol solutions(pH3.6)for 15min
本文研究發現發酵產物乙醇、乙酸、乳酸對BG活力有不同程度抑制,其IC50分別為4.4,0.34和0.09 mol/L,抑制強度依次為乳酸>乙酸>乙醇,乙醇IC50明顯大于前兩者,表明該菌株的BG更適合于纖維素發酵產乙醇,而且同Aspergillusoryzae[10]產BG相比,乙醇耐受性更高.三者抑制BG均表現為可逆非競爭性抑制,一方面說明發酵產物以非共價鍵與酶蛋白中的必需基團結合,除去發酵產物后而可使酶重新恢復活性,另一方面,證明乙醇等3種物質不結合在酶活性中心的底物結合部位上,而是結合在其以外的必需基團上,所以并不降低酶對底物的親和力,但能阻止酶的催化作用.3種物質具有相近的烴骨架,乙醇不像乙酸和乳酸都可解離出H+,降低酶所處環境的pH,只是單純靠氫鍵與酶分子發生反應,故抑制作用較后兩者弱.而且,乳酸含有一個羥基和羧基,綜合了乙醇和乙酸的作用,作用比另兩者更強,體現為IC50比乙醇和乙酸低,BG對其耐受度比乙醇或乙酸低.該結果另一方面驗證羧基化合物比中性羥基化合物更能抑制灰綠曲霉BG活力,故在無氧發酵中應注意控制氧化程度,避免過度產酸.
BG在含乙醇的催化反應體系中,熒光強度顯著增大,即分子構象改變,酶分子在乙醇溶液中的構象變化主要表現為酶分子的主肽鏈從緊密的構象趨于松散狀態,從有序的二級結構變成無規則線團.可能是因為乙醇介入酶分子中,改變酶分子所處環境的介電常數,同時引起肽鏈伸展,構象改變,使得Tyr和Trp殘基所處微環境的極性降低;此外,Tyr殘基形成的氫鍵遭到了破壞,處于淬滅狀態的Tyr殘基熒光得以恢復,使其量子率增加,于是熒光強度增大[11].當乙醇體積分數為20%時,熒光強度顯著增加,但吸收峰波長沒有觀察到顯著的紅移,說明酶分子的整體構象尚未完全變性,此時BG還有較大活力.BG在不同量乙醇和不同反應溫度的酶活力變化,可能是乙醇和溫度對酶分子綜合作用的結果.一方面酶的最適反應溫度是活性中心活化與分子變性共同作用的結果,一定量乙醇存在時,由于乙醇作用,隨著溫度上升,酶分子失效加速,故BG合適反應溫度比無乙醇時更低.另一方面,溫度較低(<55℃)時,若乙醇體積分數較低(<20%),乙醇分子使酶蛋白內的次級鍵減弱,酶分子活性中心更加舒展,柔性增加,底物與酶結合更容易,酶催化效率相應地增加;但如果乙醇體積分數較高(20%),乙醇對酶分子的破壞作用占主導,即使溫度降低,酶活力仍然不及前者.
葡萄糖是纖維素酶水解纖維素的最終產物,但在真菌培養或者工業生產中,葡萄糖常常會在適宜的條件下繼續發酵產酸或乙醇,這些發酵產物會對纖維素酶產生反饋抑制,影響酶的活力,以上研究為發酵進程的控制,控制發酵工藝提供了理論基礎.
[1]Sang J H,Yong J Y,Hyen S K.Characterization of a bifunctional cellulase and its structural gene [J].J Biol Chem.,1995,270(43):26012-26019.
[2]Xin Z,Yinbo Q,Peiji G.Acceleration of ethanol production from paper mill waste fiber by supplementation withβ-glucosidase[J].Enzyme Microb Technol,1993,15:62-65.
[3]Bhatia Y,Mishra S,Bisaria V S.Microbialβ-glucosidases:cloning,properties,and applications[J].Crit Rev Biotechnol,2002,22(4):375-407.
[4]蔣西蘭,李文利.纖維素乙醇基因工程研究進展[J].中國生物工程雜志,2009,29(7):127-133.
[5]陳衛紅,李多川.嗜熱子囊菌光孢變種β-葡萄糖苷酶的基因克隆、表達及酶學性質的研究分析[J].應用與環境生物學報,2009,15(4):549-554.
[6]Michael E H,Mark F R,Charles E W.Cellulase for commodity products from cellulosic biomass[J].Curr Opin Biotech,1999,10:358-364.
[7]潘利華,羅建平.β-葡萄糖苷酶的研究及應用進展[J].食品科學,2006,27(12):803-806.
[8]徐昶,龍敏南,鄔小兵,等.高產纖維素酶菌株的篩選及產酶條件研究[J].廈門大學學報:自然科學版,2004,43(1):107-111.
[9]Ma S J,Leng B,Xu X Q,et al.Purification and characterization ofβ-1,4-glucosidase fromAspergillusglaucus[J].Afr J Biotechnol,2011,10(84):19607-19614.
[10]葉海梅.米曲霉FZ58β-葡萄糖苷酶固體發酵優化及酶學特性的研究[D].福州:福建師范大學,2008.
[11]王悅,謝曉蘭,王勤,等.乙醇對南美白對蝦 N-乙酰-β-D-氨基葡萄糖苷酶活力與構象的影響[J].廈門大學學報:自然科學版,2004(增刊1):28-31.
