氮磷限制條件下螺旋魚腥藻產(chǎn)生土嗅素特征研究
2014-08-03 03:20:20賈曉燕儲(chǔ)昭升胡小貞中國環(huán)境科學(xué)研究院湖泊環(huán)境創(chuàng)新基地環(huán)境基準(zhǔn)與風(fēng)險(xiǎn)評估國家重點(diǎn)實(shí)驗(yàn)室北京0002北京科技大學(xué)土木與環(huán)境工程學(xué)院北京00083
中國環(huán)境科學(xué) 2014年4期
賈曉燕,劉 聰,2,儲(chǔ)昭升*,龐 燕,胡小貞 (.中國環(huán)境科學(xué)研究院湖泊環(huán)境創(chuàng)新基地,環(huán)境基準(zhǔn)與風(fēng)險(xiǎn)評估國家重點(diǎn)實(shí)驗(yàn)室,北京 0002;2.北京科技大學(xué)土木與環(huán)境工程學(xué)院,北京 00083)
隨著人類生活水平的不斷提高,對水質(zhì)量的要求越來越高,水體異味已成為研究關(guān)注的重點(diǎn).異味問題廣泛存在于各類淡水水體中,早在1883年就出現(xiàn)了有關(guān)水體異味的報(bào)道[1].已有研究表明了產(chǎn)生水體異味的主要原因[2]是水體富營養(yǎng)化,在富營養(yǎng)化水體中,由于營養(yǎng)物質(zhì)過剩,藻類過度生長,由藻類代謝和腐爛分解產(chǎn)生的異味物質(zhì)是水體中異味物質(zhì)的主要來源.土霉味是淡水水體中存在最廣泛、且最難聞的異味.引起水體土霉味的主要物質(zhì)之一是土嗅素[3-5].螺旋魚腥藻是一種常見的土嗅素產(chǎn)生藻.影響螺旋魚腥藻產(chǎn)生土嗅素的因素主要有溫度[6]、光照[7]和營養(yǎng)鹽[8-9].
目前國內(nèi)關(guān)于水體異味及土霉味的研究主要集中在異味的分布及原因分析[10-12],異味物質(zhì)的檢測技術(shù)[13-14]及水體異味的防治措施[15-16]等方面,開展室內(nèi)模擬實(shí)驗(yàn),分析水體中氮磷含量對藻類生長及產(chǎn)生異味物質(zhì)影響的研究鮮有報(bào)道.本文通過研究在氮限制和磷限制條件下,藻細(xì)胞密度、葉綠素 a的合成以及土嗅素產(chǎn)生量等,探討氮限制和磷限制對螺旋魚腥藻生長、土嗅素生成及釋放的影響,為更好的預(yù)測和控制魚腥藻水華暴發(fā)和預(yù)防由此產(chǎn)生的異味事件提供參考.
1 材料與方法
1.1 藻種及培養(yǎng)
本實(shí)驗(yàn)所用螺旋魚腥藻從河北省秦皇島市洋河水庫分離獲得,在中國環(huán)境科學(xué)研究院藻類培養(yǎng)室進(jìn)行培養(yǎng).采用 M11培養(yǎng)基[17](mg/L:NaNO3100, K2HPO410, MgSO4·7H2O 75,CaCl2·2H2O 40, Na2CO320, Na2EDTA·2H2O 0.1),在溫度 26℃,光照強(qiáng)度 2500lx、光暗比 12h:12h的條件下培養(yǎng).
1.2 培養(yǎng)實(shí)驗(yàn)
本實(shí)驗(yàn)在10L的玻璃瓶中分為2組進(jìn)行,分別為缺氮條件和缺磷條件,每組設(shè)置兩個(gè)平行.在每個(gè)培養(yǎng)瓶中加入8L M11培養(yǎng)基和200mL的洋河水庫底泥濾液,其中氮限制控制氮的濃度為1/10M11培養(yǎng)基,則氮的濃度為 1.647mg/L(N:P=0.93);而磷限制則控制磷的濃度為 1/10M11培養(yǎng)基,則磷的濃度為 0.178mg/L(N:P=92.53),然后用高壓滅菌鍋在121℃下滅菌30min.
藻種在接種前分別在缺氮和缺磷的M11培養(yǎng)基中進(jìn)行3d的饑餓培養(yǎng).在無菌條件下接種至10L玻璃瓶中,接種量為 5×103cells/mL,培養(yǎng)時(shí)曝氣量為2.5L/min.藻細(xì)胞密度和葉綠素a每5d取樣進(jìn)行分析,異形胞和土嗅素含量每3d取樣進(jìn)行分析.
1.3 測定方法
藻細(xì)胞密度和異形胞用浮游植物計(jì)數(shù)板在光學(xué)顯微鏡下測定,每個(gè)樣品計(jì)數(shù)3次.土嗅素采取固相微萃取(美國 Supelco公司)與 HP7890/5975氣相色譜-質(zhì)聯(lián)用儀(美國Agilent公司)進(jìn)行測定[18].用玻璃纖維濾膜(Whatman GF-C)對藻液進(jìn)行過濾后測定胞外土嗅素,用未過濾的藻液測定土嗅素總量.胞內(nèi)土嗅素含量為土嗅素總量減去胞外土嗅素含量.葉綠素a采用玻璃研磨器,丙酮提取,分光光度法測定[19].總氮和溶解性總氮采用過硫酸鉀氧化,紫外分光光度法測定[20];總磷和溶解性總磷采用過硫酸鉀氧化,鉬銻抗分光光度法測定[21].
2 結(jié)果
2.1 螺旋魚腥藻的生長特征
螺旋魚腥藻在磷限制條件下的生長受到抑制,生長遲緩,而在氮限制條件下迅速進(jìn)入一個(gè)相對較快的生長期(如圖 1所示).磷氮限制條件下最大藻細(xì)胞密度分別為 5.48×104,2.26×105cells/mL,后者是前者的4倍.
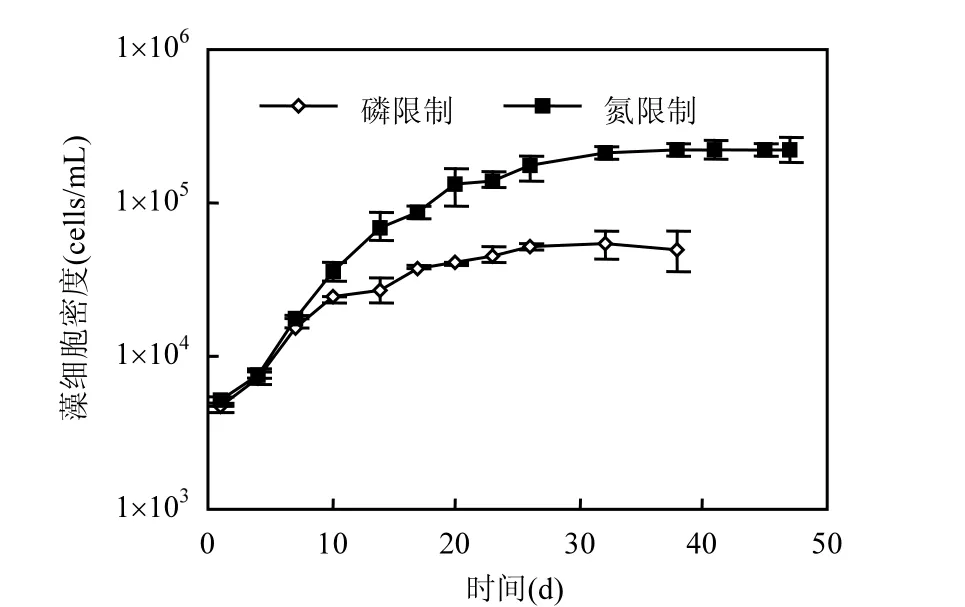
圖1 螺旋魚腥藻的生長曲線Fig.1 Growth curves of Anabaena sp.population

圖2 氮限制條件下異形胞形成的百分比Fig.2 Ratios of heterocyst under nitrogen-limited condition
螺旋魚腥藻在磷限制下幾乎不生成異形胞,在氮限制下則極易生成異形胞.在螺旋魚腥藻培養(yǎng)到第20d后異形胞形成比例為3.5%~4.4%(如圖2所示).
2.2 土嗅素生成量的變化趨勢

圖3 磷氮限制條件下土嗅素濃度的變化趨勢Fig.3 Varietion of geosmin concentration under phosphoruslimited condition and nitrogen-limited condition
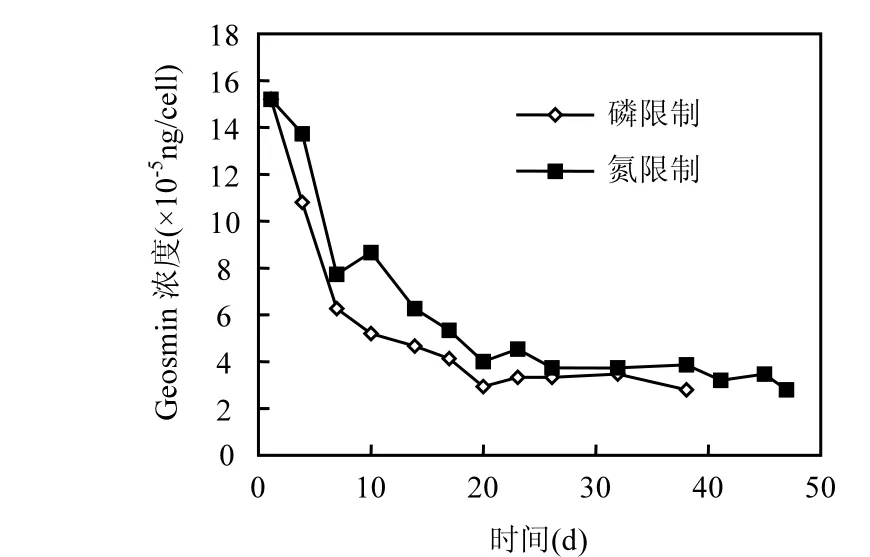
圖4 單位細(xì)胞所含土嗅素濃度的變化Fig.4 Varietion of geosmin concentration in unit cell
在螺旋魚腥藻的整個(gè)生長過程中,其胞內(nèi)土嗅素的含量都遠(yuǎn)遠(yuǎn)大于胞外(圖3).在磷限制條件下,土嗅素的最高濃度為 1.91×103ng/L;而在氮限制條件下,土嗅素的最高濃度為8.64× 103ng/L,遠(yuǎn)遠(yuǎn)高于前者.胞內(nèi)土嗅素的濃度變化與藻細(xì)胞密度的變化趨勢基本一致.
在磷限制和氮限制2種培養(yǎng)條件下(圖4),單位細(xì)胞內(nèi)土嗅素的濃度在培養(yǎng)的前 20d內(nèi)都處于急速下降的趨勢,之后開始趨于平緩,分別維持在 3.18×10-5,3.68×10-5ng/cell左右.
2.3 葉綠素a的含量及與土嗅素含量的關(guān)系
螺旋魚腥藻單位細(xì)胞生成的葉綠素 a含量的變化趨勢(圖 5)表明,在磷限制條件下,每細(xì)胞所含葉綠素 a的濃度在培養(yǎng)的前 7d急速下降,之后基本維持在 1.89×10-3ng/cell;而在氮限制條件下,每細(xì)胞所含葉綠素 a濃度的下降趨勢并不明顯,維持在3.67×10-3ng/cell左右.在氮限制條件下,魚腥藻更易形成葉綠素 a,在磷限制條件下,魚腥藻形成葉綠素 a的過程受到抑制,其值遠(yuǎn)低于氮限制條件下葉綠素a的量.
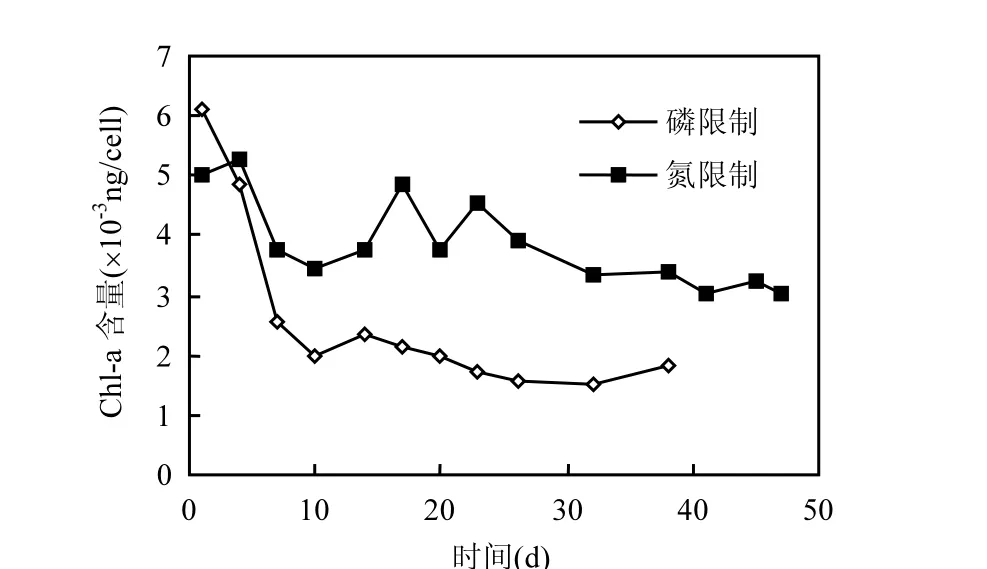
圖5 單位細(xì)胞所含葉綠素a濃度的變化趨勢Fig.5 Varietion of chl a concentration in unit cell
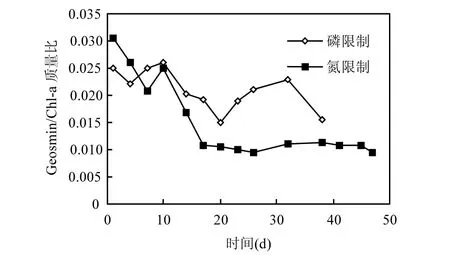
圖6 單位細(xì)胞分泌土嗅素與葉綠素質(zhì)量比的變化趨勢Fig.6 Varietion of geosmin/Chl a ratio in unit cell
魚腥藻單位細(xì)胞產(chǎn)生的土嗅素與葉綠素 a的質(zhì)量比(geosmin/chl a)在磷限制條件下要高于氮限制條件下(圖6).
2.4 培養(yǎng)過程中營養(yǎng)鹽的變化
在磷限制條件下,隨著藻細(xì)胞的生長,藻液中的溶解性總磷(DTP)逐漸降低(由 0.13mg/L降為0.02mg/L)(圖 7).在氮限制條件下,魚腥藻首先利用培養(yǎng)基中的氮源,藻液中的總氮(TN)和溶解性總氮(DTN)都有所下降(圖 7),當(dāng)培養(yǎng)基中氮源不能滿足魚腥藻生長需要時(shí),魚腥藻開始生成異形胞,并進(jìn)行固氮作用.所以隨著藻細(xì)胞密度的增加,藻液中總氮(TN)的濃度有所提高.

圖7 磷限制下水體中磷的變化趨勢和氮限制下水體中氮的變化趨勢Fig.7 Varietion of phosphorus content in water column under phosphorus-limited condition and varietion of nitrogen content in water column under nitrogenlimited condition
3 討論
在磷限制和氮限制條件下,魚腥藻的生長情況不同.Rashash等[22]研究魚腥藻時(shí),發(fā)現(xiàn)其藻細(xì)胞密度在低磷情況下比在低氮情況下明顯減少.這主要是由于螺旋魚腥藻為固氮藍(lán)藻,當(dāng)其處在缺氮條件下時(shí),魚腥藻會(huì)形成具有固氮作用的異形胞,異形胞可以直接固定大氣中的N2(分子態(tài)),形成藻細(xì)胞可以利用的氮素化合物,從而使其生長并不受到很大影響.在本研究中,在氮限制條件下,異形胞形成比率約為 3.8%,螺旋魚腥藻生長未受到明顯影響.在磷限制條件下,螺旋魚腥藻幾乎不生成異形胞且生長受到抑制;所以可以通過控制排入水體中磷的含量,使水體處于磷限制,更能有效避免魚腥藻過度生長.
在生成土嗅素方面,磷限制條件下單位細(xì)胞土嗅素的生成量比氮限制條件下的生成量低約15%.Wu等[23]研究魚腥藻發(fā)現(xiàn)魚腥藻中的異形胞比其他細(xì)胞產(chǎn)生更多的土嗅素,本研究中螺旋魚腥藻在氮限制條件下更易產(chǎn)生異形胞,推測由異形胞產(chǎn)生的土嗅素是螺旋魚腥藻單位細(xì)胞土嗅素的生成量在氮限制條件下高于磷限制條件下的主要原因.
不論是氮限制還是磷限制條件下,螺旋魚腥藻胞內(nèi)土嗅素的含量都遠(yuǎn)遠(yuǎn)大于胞外土嗅素的含量,這說明螺旋魚腥藻在整個(gè)生長過程中,其生成的土嗅素大部分都保留在細(xì)胞內(nèi),只有很少一部分釋放到了水體.劉妍娟等[24]在研究螺旋魚腥藻土嗅素的產(chǎn)生和分布規(guī)律時(shí)報(bào)道螺旋魚腥藻的胞內(nèi)土嗅素占總土嗅素的85%-95%,說明土嗅素在螺旋魚腥藻生長過程中主要分布在藻細(xì)胞內(nèi)部;Jüttner等[25]研究表明在藻細(xì)胞內(nèi)生長旺盛期(約前 4d),細(xì)胞內(nèi)異味化合物濃度不斷增加,98%左右的異味化合物存在于細(xì)胞內(nèi),隨著細(xì)胞的衰老(第4d后),細(xì)胞內(nèi)異味化合物濃度開始下降,而培養(yǎng)液中的異味化合物的濃度卻不斷增加;Rashash等[22]研究得出,在生長的初期,魚腥藻產(chǎn)生的土嗅素大部分保留在了細(xì)胞里,在 20d之后,細(xì)胞內(nèi)和水體中的土嗅素比例相當(dāng);Rosen等[26]研究揭示魚腥藻細(xì)胞的死亡導(dǎo)致了細(xì)胞內(nèi)土嗅素向介質(zhì)中的釋放.本研究中在營養(yǎng)鹽限制條件下,螺旋魚腥藻的生長受到抑制,未達(dá)到衰敗期,故仍處于生成的土嗅素保留在細(xì)胞內(nèi)的階段.基于上述情況,在預(yù)防水體中發(fā)生螺旋魚腥藻引起的嗅味問題時(shí),可以通過過濾去除水體中的過量的藻類,從而減小發(fā)生水體的嗅味問題幾率.
4 結(jié)論
4.1 螺旋魚腥藻在磷限制下生長受到限制,幾乎不生成異形胞;在氮限制下生長未受到明顯影響,平均生成 3.8%異形胞.在磷限制條件下,螺旋魚腥藻產(chǎn)生土嗅素的最高濃度為 1.91×103ng/L;在氮限制條件下,土嗅素的最高濃度為 8.64×103ng/L.
4.2 不論是氮限制還是磷限制,螺旋魚腥藻在整個(gè)生長過程中,其生成的土嗅素大部分都保留在細(xì)胞內(nèi),只有很少部分(0.2%~10.4%)釋放到了水體.
4.3 魚腥藻單位細(xì)胞產(chǎn)生的土嗅素與葉綠素 a的質(zhì)量比(geosmin/chl a)在磷限制條件下要高于氮限制條件下.
[1]Farlow W G.Relation of certain forms of algae to disagreeable tastes and odors [J].Science, 1883,2:333-336.
[2]李 林.淡水水體中藻源異味化合物的分布、動(dòng)態(tài)變化與降解研究 [D].武漢:中國科學(xué)研究院水生生物研究所, 2005.
[3]Izaguirre G, Hwang C J, Krasner S W, et al.Geosmin and 2-methylisoborneol from cyanobacteria In three water supply systems [J].Appl.Environ.Microbiol., 1982,43:708-714.
[4]Li L, Wan N, Gan N Q, et al.Annual dynamics and origins of the odorous compounds in the pilot experimental area of Lake Dianchi [J].China.Wat.Sci.Tech., 2007,55(5):43-50.
[5]Izaguirre G, Taylor W D.Geosmin and MIB events in a new reservoir in southern California.[J].Wat.Sci.Tech., 2007,55(5):9-14.
[6]Zhang T, Li L, Song L, et al.Effects of temperature and light on the growth and geosmin production of Lyngbya kuetzingii(Cyanophyta) [J].Journal of Applied Phycology, 2009,21(3):279-285.
[7]Tsuchiya Y, Matsumoto A.Characterization of Oscillatoria F.Granulata producing 2-Methylisoborneol and geosmin [J].Wat.Sci.Tech., 1999,40(6):245-250.
[8]Ismail M K, Sadoun K K.Environmental and nutritional factors affecting geosmin synthesis by Anabaena sp [J].Water Research,2001,35(5):1209-1218
[9]Rashash D M C, Dietrich A M, Hoehn R C, et al.The influence of growth conditions on odor-compound production by two chrysophytes and two cyanobacteria [J].Chemical Ecology and Biology, 1995,31(11):165-172.
[10]鄧緒偉,陶 敏,張 路,等.洞庭湖水體異味物質(zhì)及其藻類和水質(zhì)的關(guān)系 [J].環(huán)境科學(xué)研究, 2013,26(1):16-21.
[11]李 林,萬 能,甘南琴,等武漢大蓮花湖異味化合物日變化及其相關(guān)因子分析 [J].水生生物學(xué)報(bào), 200731(1):112-117.
[12]齊 敏,孫小雪,鄧緒偉,等太湖中不同形態(tài)異味物質(zhì)含量、相互關(guān)系與其環(huán)境因子關(guān)系的探討 [J].湖泊科學(xué), 2012,26(4):1040-1047.
[13]黎 雯,徐 盈,吳文忠,等.水體中異味化合物定量測定的一種有效方法 [J].分析測試技術(shù)與儀器, 1998,4(2):84-90.
[14]李 林,宋立榮,甘南琴,等.頂空固相微萃取-氣相色譜-質(zhì)譜測定水中異味化合物 [J].分析化學(xué), 2005,33(8):1058-1062.
[15]劉 欣,何 進(jìn),喻子牛.微生物產(chǎn)生的土腥味化合物及其清除方法 [J].中國生物工程雜志, 2005,25(8):35-38.
[16]郭烽武,武周虎,姚 杰,等.水的異味去除技術(shù)研究進(jìn)展 [J].青島建筑工程學(xué)院院報(bào), 2005,26(1):55-58
[17]Kuwabara K.Temperature effect on growth characteristics and competition between cyanobacteria Microcystis sp.and Oscillator sp.: a experimental study using lake simulator [R].日本:第九回世界湖沼會(huì)議, 2002.
[18]梁存珍,王東升,桑義敏,等.飲用水中土霉味物質(zhì)測定方法的優(yōu)化 [J].中國給水排水, 2007,23(10):76-78.
[19]金相燦,屠清瑛.湖泊富營養(yǎng)化調(diào)查規(guī)范 [M].北京:中國環(huán)境科學(xué)出版社, 1990.
[20]GB/T 11894-198 水質(zhì)-總氮的測定-堿性過硫酸鉀消解紫外分光光度法 [S].
[21]GB/T 11893-1989 水質(zhì)-總磷的測定-鉬銻抗分光光度法 [S].
[22]Rashash D M C.The influence of growth conditions on odor-compound production by two chrysophytes and two cyanobacteria [J].Water.Sci.Tech., 1995,31(11):165-172.
[23]Wu J T.Variation of geosmin content in Anabaena cells and its relation to nitrogen utilization [J].Arch.Microbiol., 1991,157:66-69.
[24]劉妍娟,儲(chǔ)昭升,金相燦,等.螺旋魚腥藻土嗅素的產(chǎn)生和分布規(guī)律 [J].中國環(huán)境科, 2009,29(10):1082-1085.
[25]Jüttner F.Biochemistry of biogenic off flavour compounds in surface waters [J].Water Science Technol., 1988,20(8/9):107-116.
[26]Rosen B H, MacLeod B W, Simpson M R.Accumulation and release of geosmin during the growth phases of Anabaena circinalls (Kutz.) Raven horst [J].Water Science and Technology,1992,25(2):185-190.
