大亞灣浮游植物DNA指紋及其與群落結(jié)構(gòu)的關系
2014-08-03 03:20:22熊毅俊王朝暉暨南大學生態(tài)學系水體富營養(yǎng)化與赤潮防治廣東普通高校重點實驗室廣東廣州510632
中國環(huán)境科學 2014年4期
關鍵詞:分析
熊毅俊,王朝暉,王 劍 (暨南大學生態(tài)學系,水體富營養(yǎng)化與赤潮防治廣東普通高校重點實驗室,廣東 廣州 510632)
浮游植物是海洋生態(tài)系統(tǒng)的重要組成成分與初級生產(chǎn)者,是魚類和其他經(jīng)濟動物的直接或間接的餌料[1].傳統(tǒng)浮游植物的分類鑒定,主要借助顯微鏡觀察,但該技術需要依靠研究者豐富的鑒定經(jīng)驗,并且許多微小浮游植物在顯微鏡下難以區(qū)分,很容易出現(xiàn)錯誤判斷[2].隨著分子生物技術的迅速發(fā)展,分子生物學方法已被廣泛應用于海洋浮游植物研究[3].
變性梯度凝膠電泳(DGGE)技術是根據(jù)在不同濃度的變性劑中DNA片段解鏈行為的不同而導致電泳遷移率發(fā)生變化,從而將片段大小相同而堿基組成不同的片段分開[4].國外對DGGE技術的應用已日趨成熟,目前研究主要集中在環(huán)境微生物群落結(jié)構(gòu)多樣性分析上,包括物種組成分析,群落相似度比較以及分子進化樹分析等[5-6].DGGE技術在浮游植物群落結(jié)構(gòu)分析上的運用較少,Wang等[7]采用甲藻的特定引物成功分析鑒定了18種實驗室培養(yǎng)甲藻株系;邵娟等[8]利用DGGE技術對常見海洋赤潮藻類及赤潮水樣進行了分析,發(fā)現(xiàn)DGGE結(jié)果與顯微鏡觀察結(jié)果具有較好的一致性;同時DGGE技術也能較好地反映淡水湖泊浮游生物群落[9].此外,利用DGGE技術還可以鑒定某些在光學顯微鏡下難以區(qū)分的超微型浮游生物[10].
大亞灣位于南海北部,是廣東省重要的養(yǎng)殖基地,也是大亞灣核電站和惠州港所在地.由于近些年該海域污染日趨嚴重,營養(yǎng)鹽狀況發(fā)生明顯變化,導致該海域的浮游植物群落結(jié)構(gòu)發(fā)生顯著變化[11].鑒于先前對大亞灣浮游植物群落結(jié)構(gòu)的研究主要集中于傳統(tǒng)方法的觀察研究,對于分子水平上的研究較為有限,僅本課題組于2011年對大亞灣浮游植物 DNA指紋進行了初步分析[12].為了進一步了解DGGE技術在浮游植物多樣性研究中的應用,揭示大亞灣海域浮游植物 DNA指紋,本文采集了大亞灣海域表層水樣,對大亞灣浮游植物進行了顯微鏡定性定量分析,并利用PCR-DGGE技術對浮游植物的 DNA指紋進行了研究,以揭示DGGE技術等分子手段在浮游植物群落結(jié)構(gòu)多樣性分析上的應用.
1 材料與方法
1.1 采樣點的設置和樣品的采集分析
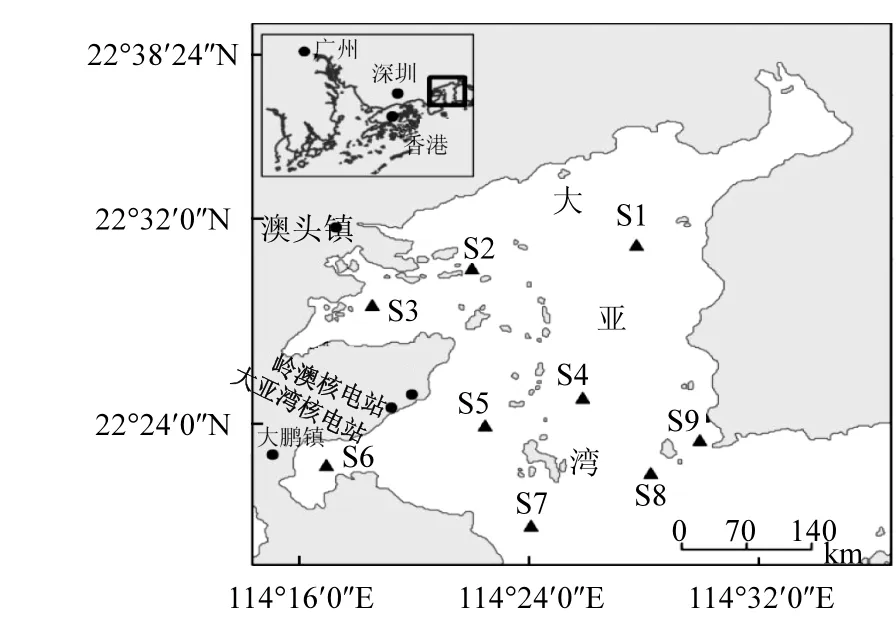
圖1 大亞灣采樣點的設置Fig.1 Sampling stations in Daya Bay
在大亞灣海域設置了 9個采樣點(圖 1),于2012年5月和6月采集表層水樣4L,用1.5%的魯格氏液固定.樣品經(jīng)靜置沉降及濃縮后,用1.2μm孔徑的濾膜(Millipore)抽濾,濾膜于-20°保存待分析.另外采集表層水樣1L,用4%的福爾馬林固定,經(jīng)濃縮后在顯微鏡下對浮游植物進行定性定量觀察.
1.2 浮游植物總DNA的提取
將濾膜冰浴剪碎,置于 1.5mL離心管中,用UNIQ柱式植物基因組提取試劑盒(上海生工)提取總 DNA,用 1%的瓊脂糖凝膠電泳檢測提取效果后,保存于4℃?zhèn)溆?
1.3 PCR擴增
PCR擴增區(qū)域為18S rDNA V3區(qū),用真核生物 V3區(qū)通用引物 F1427GC(5′-CGCCCGCCCG CGCCCCGCGCCCGGCCCGCCGCCCCCGCCC CTCTGTGATGCCCTTAGATGTTCTGGG-3')和R1616(5'-GCGGTGTGTACAAAGGGCAGGG-3')[13]對提取的浮游植物總 DNA 進行擴增,引物由上海生工生物工程技術服務有限公司合成.
PCR反應在Applied Biosystems verityTMPCR儀中進行,反應體系為 30μL,包括3μL 10×反應緩沖液、0.8μL 上、下游引物(10μmol/L)、0.8μL dNTPs(10mmol/L)、0.5μL Taq DNA 聚合酶(5U/μL)、模板量1.5μL,最后以ddH2O補足體積.PCR反應條件為94℃ 1min,94℃變性1min,65℃退火1min(每個循環(huán)降低1℃),72℃延伸1min,10個循環(huán);94℃變性1min,55℃退火 1min,72℃延伸1min,20個循環(huán);最后72℃延伸10min.用1%的瓊脂糖凝膠對PCR產(chǎn)物進行電泳檢測,擴增出來的目的片段大小為 210bp,大小均一,適于DGGE電泳擴增范圍(200~600bp).
1.4 DGGE分析
取 PCR 產(chǎn)物 30μL,在 Bio-Rad公司的DcodeTM通用突變檢測系統(tǒng)對 PCR產(chǎn)物進行DGGE分析.DGGE條件為:丙烯酰胺質(zhì)量分數(shù)8%,變性梯度 30%~70%(100%的變性劑濃度為7mol/L尿素和質(zhì)量分數(shù) 40%去離子甲酰胺),先用200V電壓電泳10min,然后再用 100V電壓電泳 16h,緩沖液為 1×TAE,電泳結(jié)束后用 SYBR GREEN I染色20min,用 Bio-Rad公司凝膠成像系統(tǒng)(GelDoc2000TM)進行拍照.
1.5 數(shù)據(jù)分析
DNA指紋圖譜用Quantity one軟件分析,其中各站點的條帶光密度值強度低于 0.05將被排除,對各站點的條帶進行標記并進行相似度分析,以Pearson系數(shù)0.5作為聚類臨界點,最后采用非加權(quán)組平均法(UPGMA)對圖譜進行聚類分析[14].
顯微鏡觀察結(jié)果用Spss19軟件進行聚類分析,為了減少不同浮游植物物種數(shù)量的差異,對浮游植物細胞密度進行了自然對數(shù)轉(zhuǎn)換.采用組間聯(lián)接的聚類方法,平方Euclidean距離的度量方法對9個不同站點進行聚類.
2 結(jié)果
2.1 DNA指紋圖譜
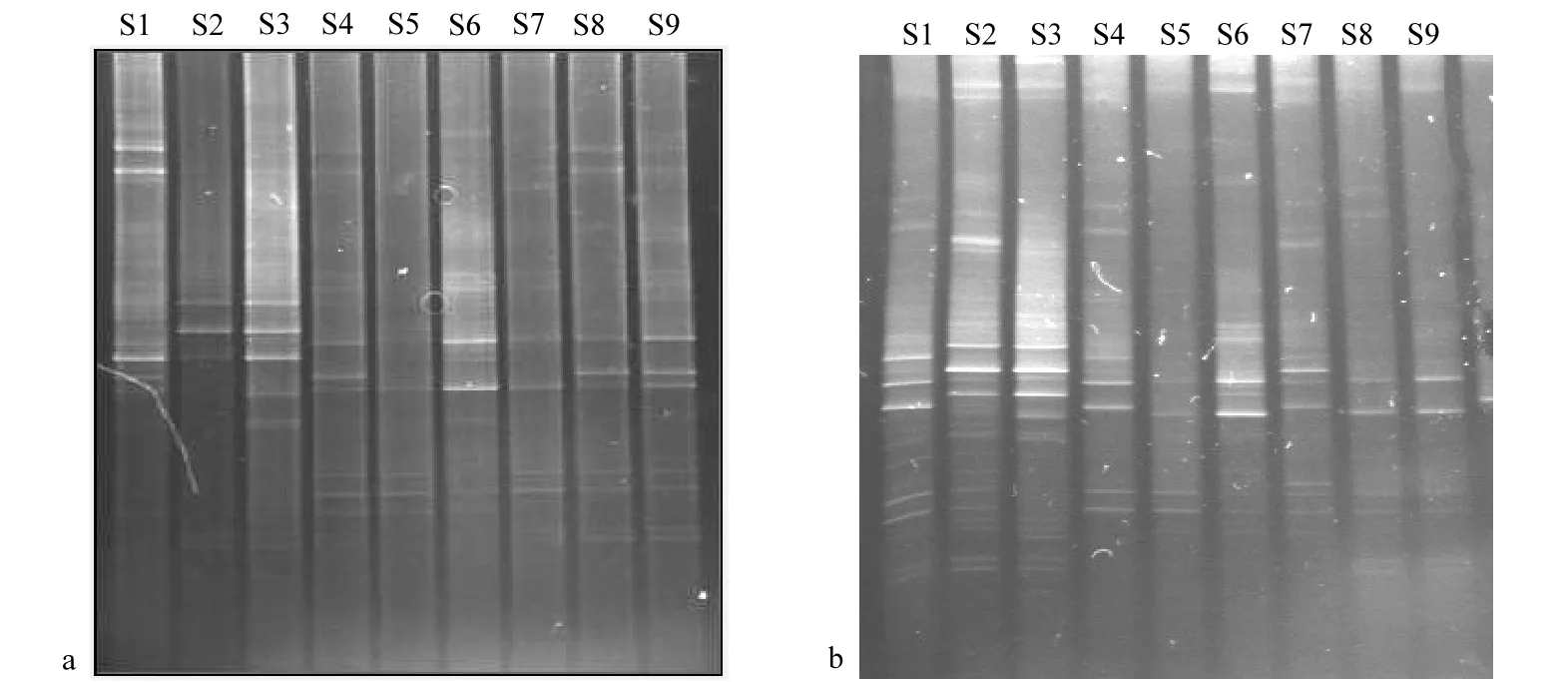
圖2 大亞灣表層浮游植物18S rDNA V3區(qū)DGGE圖譜Fig.2 DGGE fingerprints of 18S rDNA V3 region of phytoplankton in surface layer of Daya Bay
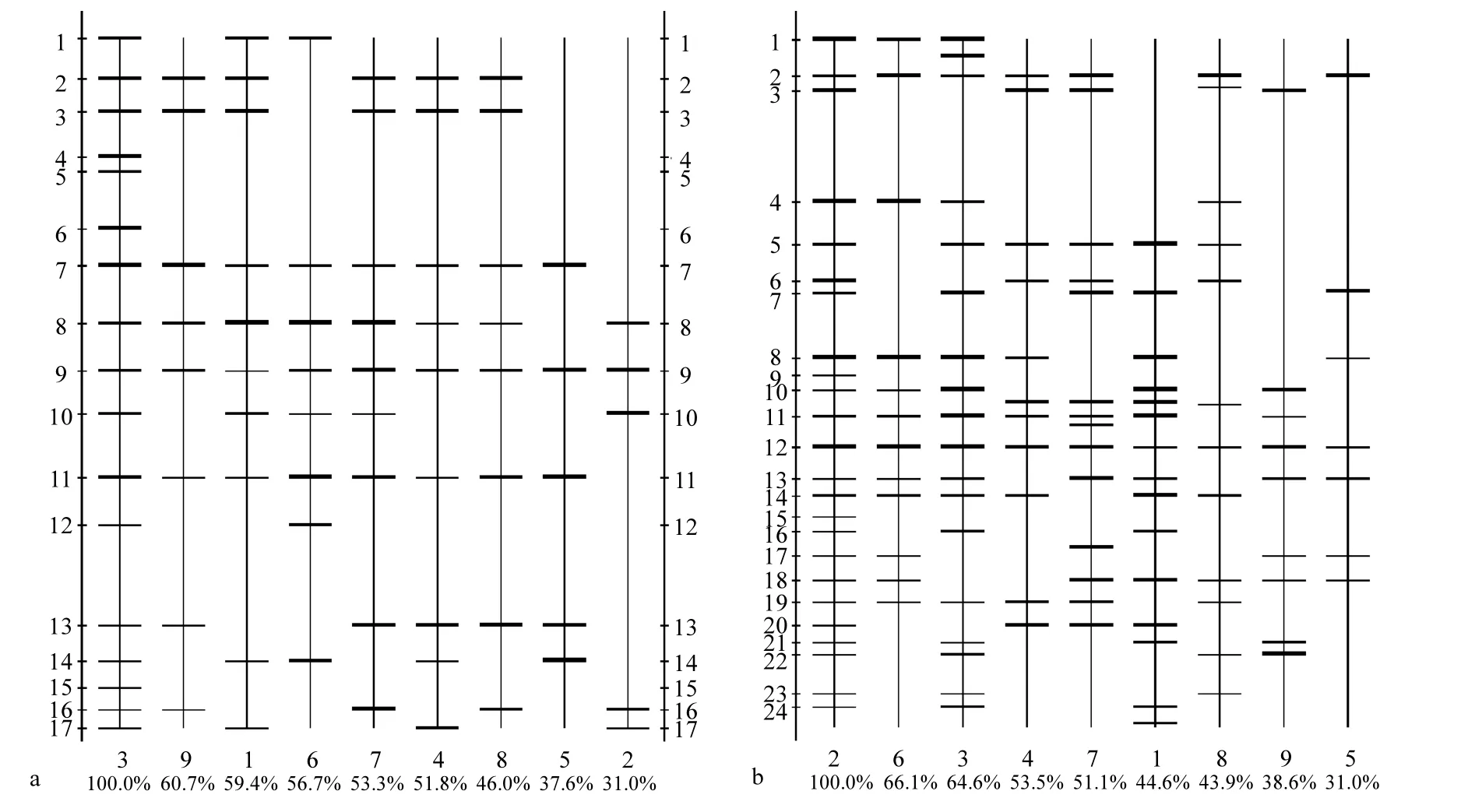
圖3 浮游植物18S rDNA V3區(qū)DGGE圖譜分析帶型圖Fig.3 DGGE profile analysis of 18S rDNA V3 region of phytoplankton
將PCR擴增產(chǎn)物進行DGGE電泳,DGGE電泳圖譜顯示各站點浮游植物的DNA指紋(圖2).較亮的條帶為優(yōu)勢種,而同一水平位置的條帶則代表同一物種.結(jié)果顯示,浮游植物DGGE圖譜條帶清晰,各站點之間優(yōu)勢種既有交叉又有所差異.用Quantity one軟件分析得到的DGGE帶型圖能更清楚顯示出DNA指紋圖譜(圖3),5月份浮游植物DGGE圖譜中共有26條不同條帶,各站點條帶數(shù)為5~17條,S3站點條帶最多,S5和S2站點較少(圖3a).其中條帶9在所有站點中均出現(xiàn),條帶7、8、11在8個站點中出現(xiàn),而條帶2、3、10、13也是常見優(yōu)勢條帶.6月份浮游植物DGGE電泳圖譜共發(fā)現(xiàn) 28條帶,每個站點條帶數(shù)為 7~24條(圖3b),其中S2站點最多,S5站點最少.與5月份相比,6月份優(yōu)勢條帶更為豐富,條帶數(shù)更多.而其中S2站點變化最大,條帶數(shù)由5月份的5條增加至6月份的24條.
2.2 浮游植物群落結(jié)構(gòu)
硅藻是大亞灣浮游植物的主要類群,其次為甲藻(圖4),還存在少量綠藻和藍細菌.5月份和6月份9個站點硅藻分別占浮游植物總數(shù)29.0%~99.4%和 7.9%~99.9%,平均分別為 85.0%和77.1%.5月份和6月份浮游植物平均細胞密度相差不大,分別為3.84×105cells/L 和 6.05×105cells/L.5月份細胞豐度最高的是 S1,為 1.08×106cells/L;S7浮游植物密度較低,為1.17×104cells/L.6月份細胞密度最高的是 S3,為2.06×106cells/L;S5細胞密度最低,為9.7×104cells/L.優(yōu)勢種硅藻包括環(huán)紋婁氏藻(Lauderia annulata)、丹麥細柱藻(Leptocylindrus danicus)、擬菱形藻(Pseudo-nitzschia spp.)、菱形海線藻(Thalassionema nitzschioides)、冰河擬星桿藻(Asterionellopsisglacialis)、中肋骨條藻(Skeletonema costatum)、細弱海鏈藻(Thalassionema subtilis)、角毛藻(Chaetoceros spp.)、小環(huán)藻(Cyclotella spp.)等.優(yōu)勢甲藻主要為小型裸甲藻屬(Gymnodinium spp.)和反曲原甲藻(prorocentrum sigmoides).
5月份反曲原甲藻在S5大量出現(xiàn),在甲藻和浮游植物中的百分比含量分別為92.2%和65.5%.6月份裸甲藻在S3、S7中大量出現(xiàn),該藻在甲藻和浮游植物中的百分比含量分別為 99.4%和91.6%以及99.1%和47.2%.2種甲藻的大量出現(xiàn)導致了甲藻在這些站點的百分比含量明顯高于其他站點,而且在5月份S5以及6月份S3中甲藻的百分比含量均超過了50%.此外,在6月份S8的浮游植物組成與其他站點也具有明顯差別,主要表現(xiàn)在其他藻類百分比的上升,這主要是由于在該樣品中,藍細菌中的色球藻(Chroococcus spp.)的大量出現(xiàn),導致了以該藻為主的其他藻類百分比上升為59.9%.
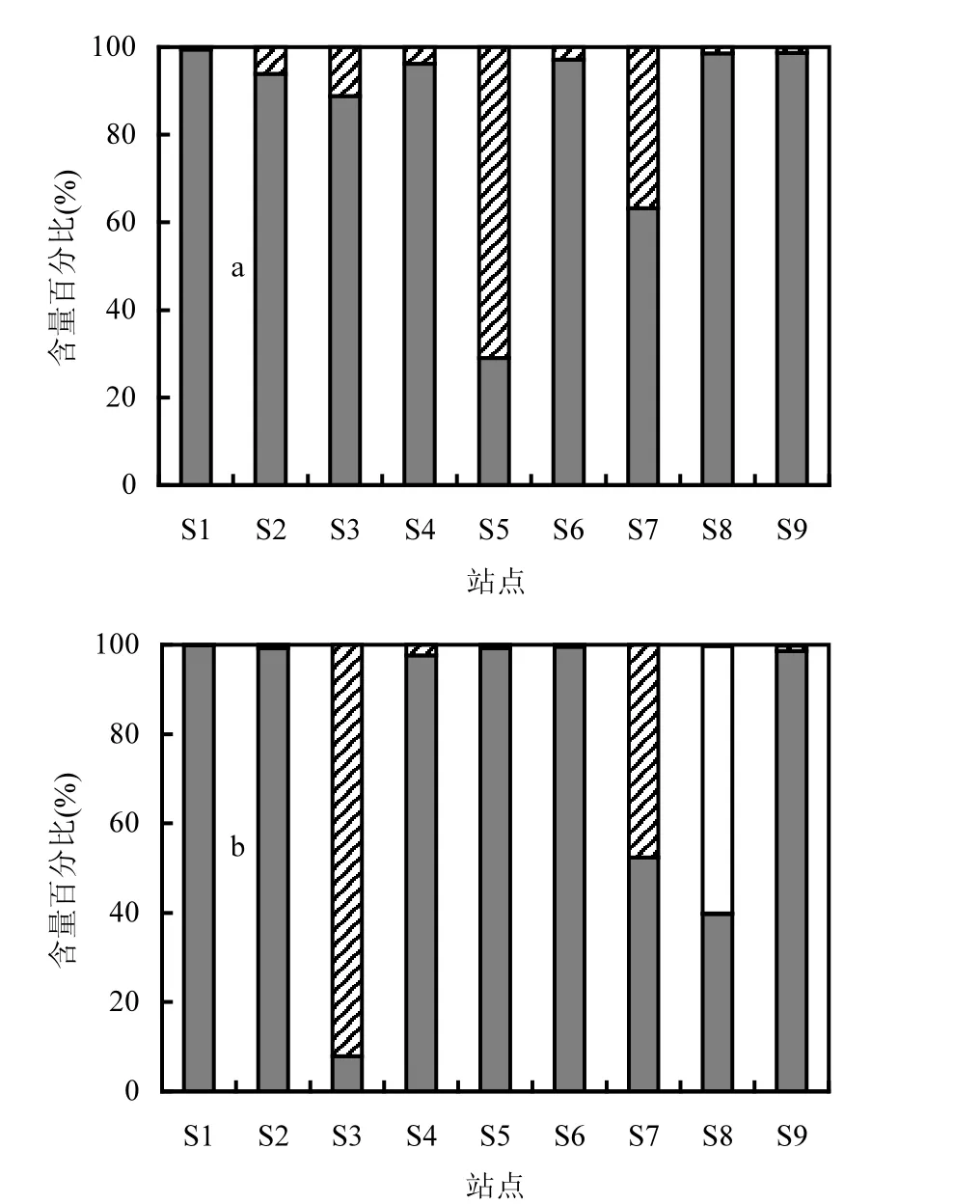
圖4 各站點不同類別浮游植物的數(shù)量百分比Fig.4 Quantitative percentages of different groups of phytoplankton in each station

2.3 浮游植物種類與DNA指紋條帶的比較
圖5可以明顯看出,顯微觀察分析得到的浮游植物種類數(shù)明顯高于DGGE條帶數(shù).在5月份和6月份的2次調(diào)查中,共分析鑒定出浮游植物72種,其中5月份為52種,每個站點鑒定的種類數(shù)為 12~26種;6月份共分析鑒定出浮游植物 54種,每個站點鑒定的種類數(shù)為12~30種.
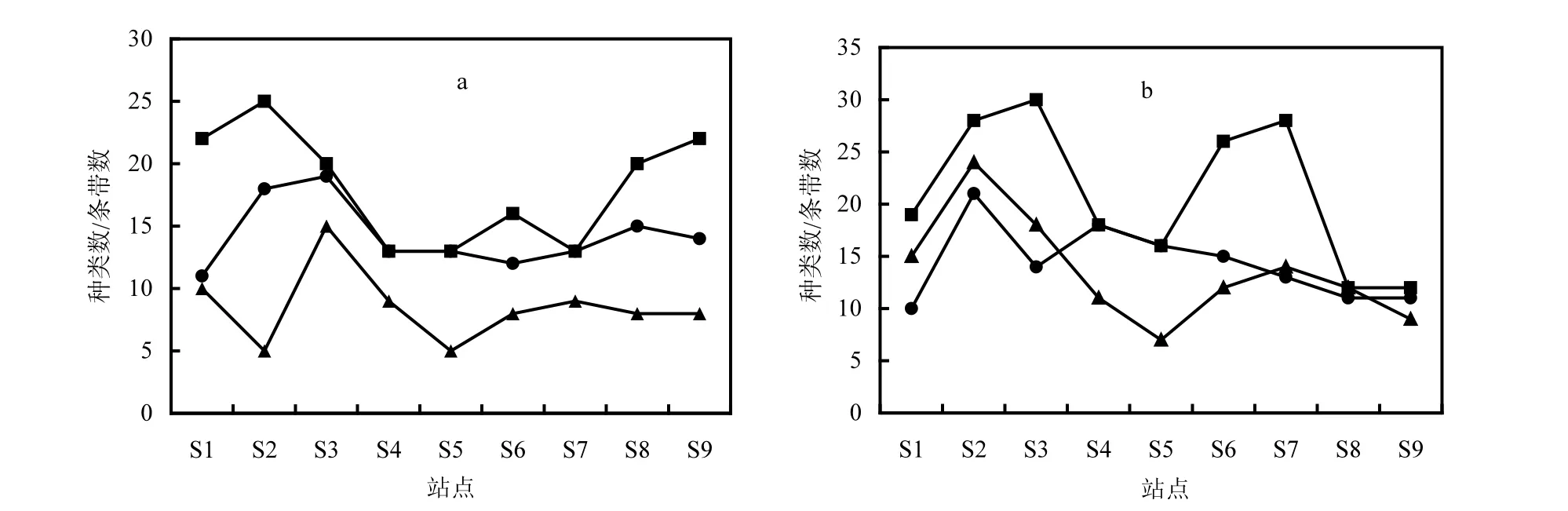
圖5 浮游植物顯微觀察種類數(shù)及DGGE指紋條帶數(shù)的比較Fig.5 Comparison between species number and DGGE fingerprint bands of phytoplankton

為了避免DGGE分析中浮游植物優(yōu)勢種類對稀有種類的屏蔽作用,將百分比含量小于0.1%的種類進行剔除,再與 DNA指紋條帶數(shù)進行比較.結(jié)果顯示,種類數(shù)與 DNA指紋條帶數(shù)變化規(guī)律基本一致(圖 5),說明 DGGE分析能在一定程度上反映浮游植物的種類多樣性.5月份優(yōu)勢種的種類數(shù)都高于DNA指紋條帶數(shù),這是由于5月份樣品中各站點細胞密度最高的優(yōu)勢種均為擬菱形藻(Pseudo-nitzschia spp.),它們的百分比含量均超過 80%,而其他所有非優(yōu)勢種的百分比較低.因此 DNA 擴增過程中,擬菱形藻很有可能對其他密度較低的非優(yōu)勢種存在屏蔽效應.而6月份優(yōu)勢種類數(shù)則與DNA條帶數(shù)相近甚至低于條帶數(shù).一方面是由于6月份浮游植物均勻度較高,優(yōu)勢種相對含量較少,而種類較為豐富,優(yōu)勢種類產(chǎn)生的屏蔽作用也較小;另一方面可能由于顯微觀察較難區(qū)分形態(tài)結(jié)構(gòu)類近的同屬種類,而在DNA指紋圖譜中卻能將同屬不同種類的浮游植物分離.
2.4 浮游植物群落以及DNA指紋聚類分析
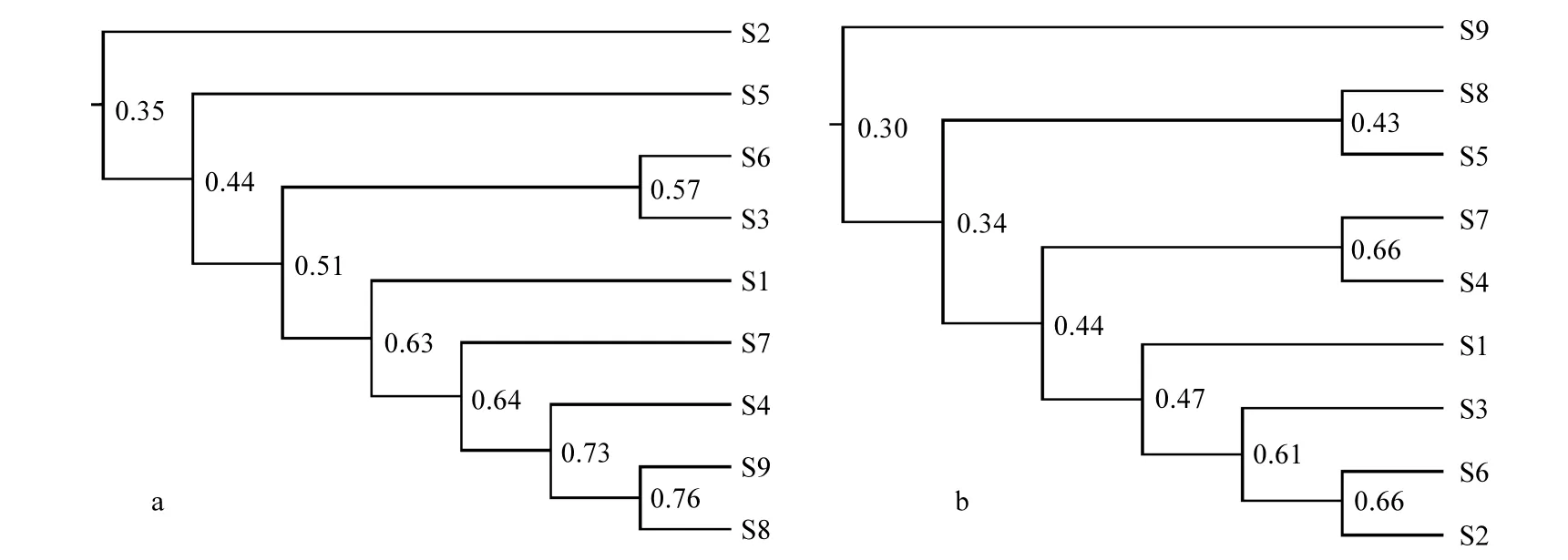
圖6 根據(jù)浮游植物DGGE指紋圖譜的聚類分析圖Fig.6 Clustering dendrogram of sampling station based on DGGE fingerprints of phytoplankton
從DGGE指紋圖譜得到的聚類分析結(jié)果與各站點的地理位置具有一定關系(圖1,圖6).近岸海域的站點如S2、S3、S6之間相似度高,聚為一類;而遠岸海域如 S1、S4、S5、S7、S8、S9聚在一起(圖 6a)或者單獨為一類(圖 6b).說明浮游植物 DNA植物具有一定的空間分布規(guī)律.而通過浮游植物群落進行聚類分析結(jié)果與DGGE指紋聚類結(jié)果相近(圖7),澳頭近岸海域的S2和S3總是聚在一起,其余較為遠岸站點聚成一大類.
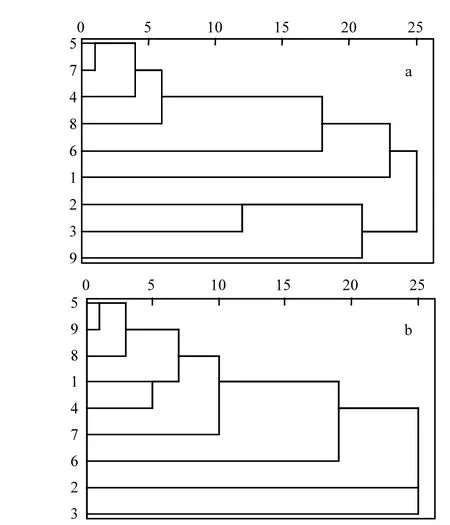
圖7 根據(jù)浮游植物群落豐度和種類數(shù)的聚類分析Fig.7 Clustering dendrogam of sampling stations based on cell abundance and species richness of phytoplankton community
3 討論
大亞灣夏季浮游植物種類豐富,共分析鑒定出浮游植物 72種;DNA指紋條帶也比較豐富,2個月份分別得到 26和 28條不同條帶.一般來說,DNA指紋條帶數(shù)要遠遠低于浮游植物種類數(shù),但是剔除非優(yōu)勢種后,DNA條帶數(shù)與種類數(shù)變化趨勢相近(圖5).結(jié)果說明DNA指紋圖譜能較大程度地反映浮游植物優(yōu)勢種群的組成,而對于相對含量較少的物種,可能會由于優(yōu)勢種的屏蔽作用而被掩蓋.作為一種分子手段, DGGE技術能夠快速、準確地分析群落結(jié)構(gòu)多樣性,彌補顯微觀察中一些難以區(qū)分的種類以及微小物種的分離鑒定[10].在本研究中,6月份的某些樣品中DNA指紋數(shù)要高于優(yōu)勢種類數(shù),說明對某些相對含量>0.1%的優(yōu)勢物種,DGGE技術對浮游植物物種的區(qū)分度要比顯微鏡分析更為精確.另一方面,由于DGGE技術本身所存在的缺陷,電泳過程中容易發(fā)生共遷移現(xiàn)象[15-16],造成堿基組成相近的DNA片段區(qū)分困難,一條DNA條帶中有時可能存在幾個相近物種的 DNA指紋;此外,對于較少的非優(yōu)勢種,由于DNA含量極少,在后續(xù)的PCR擴增中,會被數(shù)量較高種類的 DNA所掩蓋,導致其無法在DGGE圖譜中出現(xiàn).針對出現(xiàn)的這些技術缺陷,目前的解決方法是將條帶切膠回收測序,通過是否可獲得單一的序列來判斷條帶攜帶的遺傳信息.由此說明 PCR-DGGE技術在分析浮游植物群落結(jié)構(gòu)上具有一定的可行性.
通過聚類分析發(fā)現(xiàn),顯微鏡觀察的聚類結(jié)果與 DNA指紋的聚類分析結(jié)果較為一致,能較為清楚地將富營養(yǎng)化嚴重的近岸海域和較為清潔的遠岸海域區(qū)分開來(圖6).由于生活污水排放以及大規(guī)模的養(yǎng)殖,南澳以及大鵬澳近岸海域富營養(yǎng)化嚴重,營養(yǎng)鹽含量高[17].本研究結(jié)果顯示,靠近養(yǎng)殖區(qū)的站點S2、S3、S6浮游植物生物量豐富,甲藻數(shù)量明顯增多,說明富營養(yǎng)化已經(jīng)對浮游植物群落結(jié)構(gòu)產(chǎn)生嚴重影響.遠岸海域的幾個站位雖然所處的海域不同,離岸遠近也有些差異,但是沒有處于富營養(yǎng)化嚴重的養(yǎng)殖區(qū)域,這幾個站位的浮游植物群落結(jié)構(gòu)和 DNA指紋均相近,在聚類分析中也聚為一大類.本研究結(jié)果說明,營養(yǎng)化程度是影響大亞灣海域浮游植物分布的重要原因.此外,位于核電站附近的 S5站點浮游植物種類、數(shù)量以及DNA指紋條帶數(shù)均較低,核電站溫排水可能也是影響浮游植物群落結(jié)構(gòu)的重要因素,特別是在夏季高溫季節(jié)[18].
此外,6月份S8樣品中發(fā)現(xiàn)有大量的藍細菌,細胞密度達到 1.28×105cells/L,約占細胞總密度的60%.藍細菌是地球的先鋒生物,具有抗高溫、抗紫外以及抗攝食能力[19],隨著水溫、紫外輻射的增加以及富營養(yǎng)化的加劇,藍細菌水華已經(jīng)成為全球水環(huán)境的巨大挑戰(zhàn)[20].大亞灣海域也已出現(xiàn)藍細菌數(shù)量上升趨勢[18],而在本研究中出現(xiàn)的大量藍細菌更進一步說明了大亞灣海域浮游植物群落結(jié)構(gòu)有向藍細菌演變的趨勢.但是在DNA指紋圖譜研究中,由于PCR擴增中使用的引物為真核生物通用引物,未能將藍細菌 DNA擴增出來.因此,在以后的 DNA指紋圖譜研究中需要設計藍細菌的特異性引物進行PCR擴增,從而更全面了解浮游植物的DNA指紋和分子多樣性.
4 結(jié)論
4.1 DGGE技術能較好地反映浮游植物優(yōu)勢種群多樣性,在顯微觀察中,一些難以區(qū)分的種類以及微小物種的分離鑒定都可以通過DGGE技術得到解決,但是對于一些相對含量較少的種類,其遺傳信息可能會被優(yōu)勢種類所屏蔽.
4.2 近海岸富營養(yǎng)化對大亞灣海域浮游植物群落結(jié)構(gòu)影響較大,浮游植物種類和細胞密度相對其他海域較高,而在靠近核電站的附近海域中,核電站溫排水對浮游植物群落結(jié)構(gòu)具有一定影響.
4.3 大亞灣海域某些站點出現(xiàn)了大量的藍細菌和裸甲藻,存在引發(fā)赤潮的風險,在今后的DNA指紋分析中需要對原核微型浮游植物進行研究.
[1]Sun J, Yu Z G, Gao Y H.Phytoplankton diversity in the East China Sea and Yellow Sea measured by PCR-DGGE and its relationships with environmental factors [J].Chinese Journal of Oceanology and Limnology, 2010,28(2):315-322.
[2]Yuan J, Mi T Z, Zhen Y, et al.Development of a real-time PCR method (Taqman) for rapid identification and quantification of Prorocentrum donghaiense [J].Journal of Ocean University of China, 2012,11(3):366-374.
[3]Niu Y, Shen H, Chen J, et al.Phytoplankton community succession shaping bacterioplankton community composition in Lake Taihu,China [J].Water Res., 2011,45(14):4169-82.
[4]Fischer S G, Lerman L S.Length-independent separation of DNA restriction fragments in two- dimensional gel electrophoresis [J].Cell, 1979,(16):191-200.
[5]Lakaniemi A M, Hulatt C J, Wakeman K, et al.Eukaryotic and prokaryotic microbial communities during microalgal biomass production [J].Bioresour.Technol., 2012,124:387-93.
[6]Hoyles L, Clear J A, McCartney A L.Use of denaturing gradient gel electrophoresis to detect Actinobacteria associated with the human faecal microbiota [J].Anaerobe, 2013,22:90-96.
[7]Wang Q, Deeds J R, Place A R, et al.Dinoflagellate community analysis of a fish kill using denaturing gradient gel electrophoresis [J].Harmful Algae, 2005,4(1):151-162.
[8]邵 娟,王朝暉,林浪聰,等.常見赤潮藻類的變性梯度凝膠電泳分析 [J].暨南大學學報, 2011,32(5):504-508.
[9]吳 利,余育和,馮偉松,等.牛山湖浮游生物群落DNA指紋結(jié)構(gòu)與物種組成的關系 [J].中國海洋大學學報, 2009,39(1):84-90.
[10]Marie D, Zhu F, Balagué V, et al.Eukaryotic picoplankton communities of the Mediterranean Sea in summer assessed by molecular approaches (DGGE, TTGE, QPCR) [J].FEMS Microbiology Ecology, 2006,55(3):403-415.
[11]Wang Z H, Qi Y Z, Chen J F.Phytoplankton abundance, community structure and nutrients in cultural areas of Daya Bay, South China Sea[J].Journal of Marine Systems, 2006,62(1):85-94.
[12]邵 娟.大亞灣浮游植物群落的季節(jié)分布特征及 DNA 指紋分析 [D].廣州:暨南大學, 2012.
[13]van Hannen E J, van Agterveld M P, Gons H J, et al.Revealing genetic diversity of eukaryotic microorganisms in aquatic environments by denaturing gradient gel electrophoresis [J].Journal of Phycology, 1998,34(2):206-213.
[14]Ying Y, Lv Z, Min H,et al.Dynamic changes of microbial community diversity in a photohydrogen producing reactor monitored by PCR-DGGE [J].Journal of Environmental Sciences, 2008,20(9):1118-1125.
[15]柴麗紅,彭 謙,徐麗華,等.DGGE技術在微生物生態(tài)學研究中的應用.[J].發(fā)酵工業(yè), 2002,28(3):20-24.
[16]Neilson J W, Jordan F L, Maier R M.Analysis of artifacts suggests DGGE should not be used for quantitative diversity analysis [J].Journal of Microbiological Methods, 2013,92(3):256-263.
[17]王朝暉,李錦蓉,齊雨藻,等.大亞灣養(yǎng)殖區(qū)營養(yǎng)鹽狀況分析與評價 [J].海洋環(huán)境科學, 2004,23(2):25-28.
[18]劉 勝,黃暉,黃良民,等.大亞灣核電站對海灣浮游植物群落的生態(tài)效應 [J].海洋環(huán)境科學, 2006,25(2):9-13.
[19]Paerl H W, Paul W J.Climate change: Links to global expansion of harmful cyanobacteria [J].Water Res., 2012,46:1349-1363.
[20]O’Neil J M, Davis T W, Burford M A, et al.The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change [J].Harmful Algae, 2012,14:313–334.
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
