胃癌相關mir-148a靶向調控胃泌素受體CCKBR
2014-08-14 07:14:18郭水龍朱圣韜王擁軍郭慶東孫秀梅張澍田
中國比較醫學雜志 2014年9期
郭水龍,朱圣韜,李 鵬,3,王擁軍,3,王 民,邢 潔,郭慶東,孫秀梅,張澍田,3
(1.首都醫科大學附屬北京友誼醫院,北京 100050;2. 消化疾病癌前病變北京市重點實驗室,北京 100050;3.北京市消化疾病中心,北京 100050 )
胃癌是嚴重危害人類健康的常見惡性腫瘤,名列全球第二大癌癥死亡原因[1]。我國是胃癌高發國,每年新發胃癌患者40萬人,發病率在所有惡性腫瘤居首位,死亡人數達30萬人,約占所有惡性腫瘤死亡人數的25%~30%[2]。胃泌素受體(也稱膽囊收縮素B受體,CCKBR)是生長因子胃泌素(Gastrin)的重要受體, 主要分布于在胃體黏膜的壁細胞和ECL細胞。生理上,CCKBR主要通過與Gastrin結合,調節胃液、胃蛋白酶和膽汁的分泌[3]。近年來的研究發現,Gastrin及其受體CCKBR與胃腸道腫瘤尤其是胃癌的發生密切相關,可作為胃癌早期診斷的重要指標,并可能發展成為胃癌治療的靶位點[4-5]。miR-148a是最早發現,同時也是最重要的胃癌相關microRNA之一,但目前對其在胃癌中的作用機制還不十分明確[6]。本研究從多個方面證實miR-148a可以靶向CCKBR,揭示了二者參與胃癌發生發展的新的功能機制。
1 材料和方法
1.1 材料
真核表達載體pIRES2-EGFP購自BD公司,miR-148a Luciferase報告載體pGL3-CM 由pGL3-control改構。BGC-823和293T細胞由本實驗室保存。CCKBR抗體購自Abcom公司,
GAPDH抗體和二抗購自中杉金橋公司。各種限制性內切酶和T4多聚核苷酸激酶購自NEB公司。引物和探針合成自Invitrogen公司。質粒抽提及凝膠回收試劑盒購自QIAGEN公司,轉染試劑購自Invitrogen公司,雙熒光素酶檢測試劑盒購自Promega公司,其他試劑均為國產分析純試劑。
1.2 方法
1.2.1 miR-148a靶向CCKBR的生物信息學預測:使用microRNA在線靶點分析工具TargetScan(www.targetscan.org)分析人CCKBR基因3’UTR,發現3處miR-148a的潛在結合位點,分別位于第334、423和476堿基處(圖1)。可以看出,miR-148a與CCKBR之間存在很好的靶向關系。
1.2.2 miR-148a真核表達載體的構建:合成miR-148a上下游引物,序列分別為5′- aaaagatctgaacacac ctgcaggaagaa-3′和5′- aaaaaagcttctggcgtctggagcactg-3′,PCR擴增出164 bp的產物序列。PCR產物經切膠回收和連接,插入pIRES2-EGFP真核表達質粒。挑陽性克隆進行測序鑒定。
1.2.3 miR-148a過表達的Northern Blot檢測:miR-148a轉染細胞后24 h提取細胞RNA,根據文獻描述方法進行Northern Blot檢測,上樣量為20 μg[7]。 miR-148a探針序列為其完全互補序列,使用T4多聚核苷酸激酶將32P 標記5’末端。
1.2.4 熒光素酶活性檢測miR-148a對CCKBR 3’UTR的靶向作用: 使用上下游引物5′-ccgctcgagg gttgaggcagggcaaatgac-3′和5′-ccgacgcgtttgggtaaggaagg agagggc-3′ 擴增人CCKBR的3′UTR,并克隆到 pGL3-CM質粒構建熒光素酶報告載體(野生型)。采用引物5′-ccggaattcattgttttaga gactatggagc-3′和5′-ccggaattccattaatctatgtgtgtgag agg-3′;5′-ccggaattcaaaa taccatcaggcctaatc-3′和5′-ccggaatt ctgattgggaagggtcactgt -3′;5′-ccggaattcaaaaggttcttcatccct ttcc-3′和5′-ccggaattcagaacagcctgttggtcaga-3′分別將CCKBR 3′UTR的 334、423和476三處miR-148a潛在結合位點進行突變,構建熒光素酶報告載體(突變型)。將報告載體與miR-148a過表達載體共轉染細胞,48 h后裂解細胞,根據試劑說明書進行檢測。檢測儀器為LB 960 Centro XS3 luminometer。
1.2.5 Western Blot蛋白檢測:使用RIPA裂解細胞提取蛋白并檢測蛋白濃度。SDS-PAGE凝膠電泳,上樣量為25 ~ 50 μg。半干法轉膜,5%脫脂牛奶封閉,一抗(1∶1000)4℃孵育過夜,HRP偶聯二抗室溫孵育1 h,ECL發光顯影。
1.2.6 統計學分析 采用 SPSS 10進行統計分析,數值采用均數±標準差表示,組間數據比較采用t檢驗,P< 0.05表示差異具有統計學意義。
2 結果
2.1 miR-148a真核表達載體的構建

圖1 人CCKBR 3’UTR 上miR-148a結合位點預測
將構建的miR-148a表達載體(IE-148a)進行測序,結果顯示 164 bp的擴增產物(miR-148a前體)正確插入載體中,序列與pubmed數據庫完全一致。Northern Blot檢測miR-148a載體在BGC-823細胞中的表達(圖2),與空載體Vector相比,過表達組miR-148a的表達量明顯增加,證實構建的載體能夠在真核細胞中過表達miR-148a,適合下一步實驗使用。
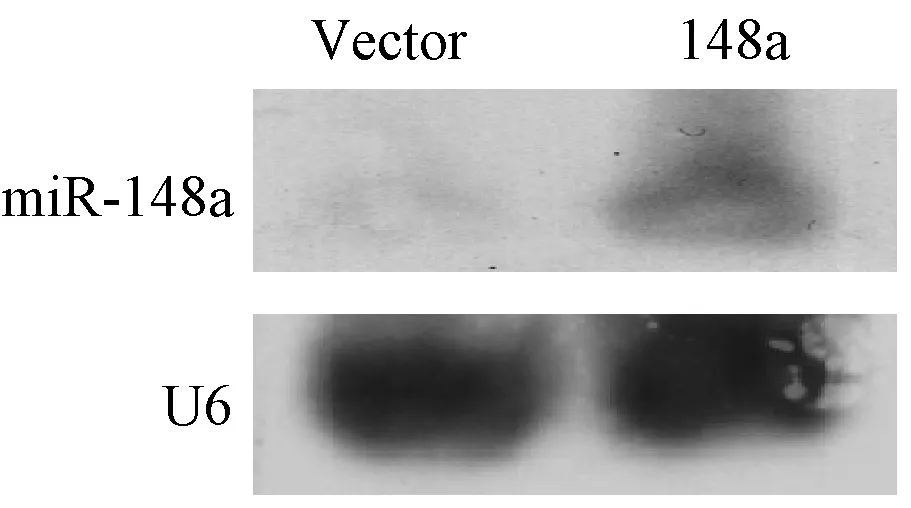
圖2 Northern Blot 檢測miR-148a真核載體表達
2.2 miR-148a通過與3’UTR 423 bp處的靶位點結合,抑制人CCKBR基因的表達
熒光素酶活性檢測結果顯示(圖3),miR-148a過表達可以明顯抑制含CCKBR 3’UTR報告載體的熒光素酶活性,大約為空載體對照組的64%(P< 0.05),表明miR-148a通過結合3’UTR區可抑制CCKBR基因的表達。當分別將三個預測的miR-148a結合位點突變后,3’UTR 423 bp處的突變可顯著消除miR-148a過表達導致的熒光素酶活性降低(P> 0.05)。而3’UTR 334 bp和476 bp處的結合位點突變沒有此效應(P< 0.05),與野生型報告載體的熒光素酶活性相同。結果表明,miR-148a通過與CCKBR 3’UTR 334 bp處結合,直接抑制該基因的表達。
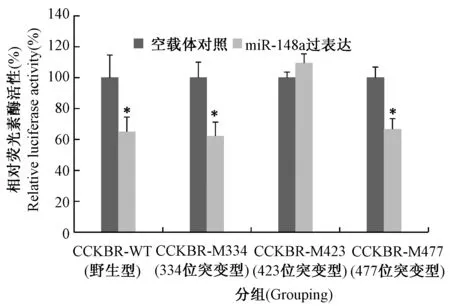
圖3 熒光素酶活性分析檢測各組報告載體表達(*P < 0.05)
2.3 miR-148a抑制CCKBR蛋白表達
Western Blot結果顯示,如圖4,與空白對照和空載體Vector相比,miR-148a過表達可顯著抑制胃癌細胞BGC-823中CCKBR的蛋白表達。
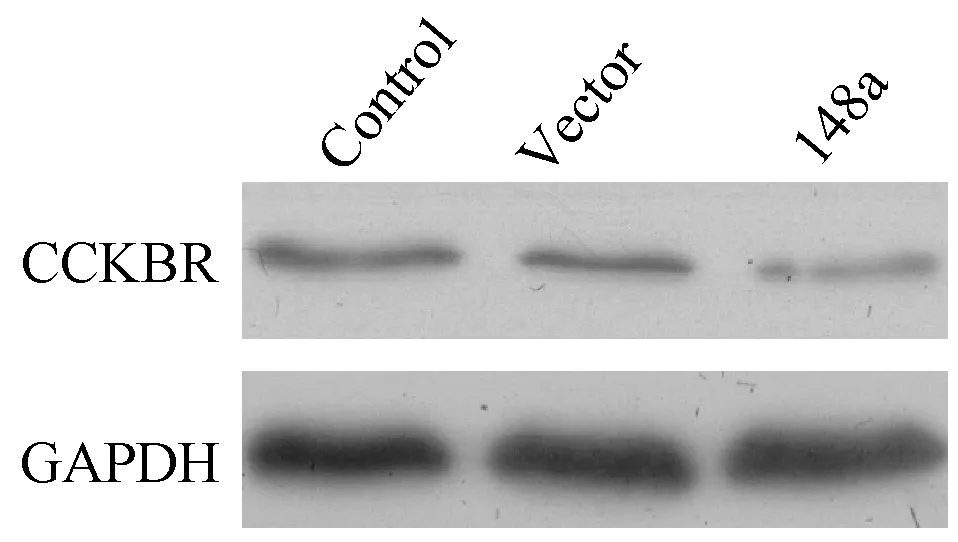
圖4 Western Blot檢測各組細胞CCKBR蛋白表達
3 討論
胃泌素主要由分布于胃竇和十二指腸近端的G細胞分泌,通過與受體結合調節胃酸和組胺的分泌來發揮作用。胃泌素在胃中作用的主要靶點是胃體黏膜的壁細胞和腸嗜鉻樣細胞(ECL)細胞表面分布的受體CCKBR。小鼠胃CCKBR基因敲除導致ECL細胞減少,胃壁變薄[8]。近年來胃泌素及其受體CCKBR在消化道腫瘤及其癌前病變中的作用越來越引起人們重視。研究證實,胃癌患者胃癌組織中CCKBR的含量明顯高于對照正常組織,胃泌素及CCKBR廣泛參與胃癌發生發展[4,9-10]。Cui等[11]發現高胃泌素血癥的日本綿鼠發生胃癌,該胃癌起源于ECL細胞,而CCKBR拮抗劑YF476可阻止高胃泌素引起的胃癌發生[12]。雖然CCKBR在胃癌發展過程中表達逐漸升高,但其異常表達的機制至今不是十分明確。
microRNA (miRNA)是一類22 bp左右的非編碼小RNA,通過序列互補與特異mRNA結合,在轉錄后水平調控靶基因的表達。我們前期的工作和眾多研究組研究結果表明,miR-148a在人胃癌組織中表達下調,并通過作用于不同的靶分子調控胃癌細胞的增殖、凋亡、侵襲和轉移等[13-17],是重要的胃癌抑制miRNAs之一。我們前期對miR-148a在胃黏膜中的表達定位研究結果顯示,miR-148a主要分布于小鼠胃黏膜底部,這一位置主要為壁細胞和ECL細胞等分泌細胞,提示miR-148a可能參與調控胃黏膜的分泌功能[14]。此外,我們前期研究還發現,miR-148a下調是胃癌發生的早期事件,胃癌小鼠模型胃組織中miR-148a在小鼠出生20 d后即觀察到50%的下調,60 d后下調約80%[18],這與胃癌發展過程中CCKBR的逐漸上升是同步的,提示二者之間可能存在調控關系。本研究中,我們首先通過信息學預測,在CCKBR中發現了3個miR-148a的結合位點。然后,通過構建的miR-148a真核表達載體并在胃癌細胞中過表達,我們證明miR-148a可直接與CCKBR的3’UTR結合,抑制其基因表達和蛋白翻譯。這一發現,為CCKBR表達調控機制和miR-148a在胃癌中的作用機制提供了新的解釋。
目前,國內外研究者都在開發新的胃癌早期診斷標志物和藥物作用靶點,而胃泌素及其受體和miRNA是其中的兩大熱點[9,19-21]。本研究的結果為胃癌早期篩查和治療提供了新的潛在靶分子,并為下一步的臨床研究奠定了基礎。
參考文獻:
[1] Kamangar F, Dores GM, Anderson WF. Patterns of cancer incidence, mortality, and prevalence across five continents: defining priorities to reduce cancer disparities in different geographic regions of the world[J]. J Clin Oncol, 2006, 24(14): 2137-2150.
[2] Yang L. Incidence and mortality of gastric cancer in China [J]. World J Gastroenterol, 2006, 12(1): 17-20.
[3] Han YM, Park JM, Park SH,etal. Gastrin promotes intestinal polyposis through cholecystokinin-B receptor-mediated proliferative signaling and fostering tumor microenvironment[J]. J Physiol Pharmacol, 2013, 64(4): 429-437.
[4] 黃亞娜,唐世孝. 胃泌素與胃癌關系的研究進展[J]. 西南軍醫, 2012, (05): 757-760.
[5] 趙蓓,吳巖. 胃泌素與腫瘤發生、發展的關系[J]. 內蒙古醫學院學報, 2008, (01): 65-68+72.
[6] Xia J, Guo X, Yan J,etal. The role of miR-148a in gastric cancer[J]. J Cancer Res Clin Oncol, 2014, 1649-1656.
[7] Sun Q, Zhang Y, Yang G,etal. Transforming growth factor-beta-regulated miR-24 promotes skeletal muscle differentiation[J]. Nucleic Acids Res, 2008, 36(8): 2690-2699.
[8] Nagata A, Ito M, Iwata N,etal. G protein-coupled cholecystokinin-B/gastrin receptors are responsible for physiological cell growth of the stomach mucosa in vivo[J]. Proc Natl Acad Sci U S A, 1996, 93(21): 11825-11830.
[9] Dimaline R,Varro A. Novel roles of gastrin[J]. J Physiol, 2014, 592(Pt 14): 2951-2958.
[10]. Rai R, Chandra V, Tewari M,etal. Cholecystokinin and gastrin receptors targeting in gastrointestinal cancer[J]. Surg Oncol, 2012, 21(4): 281-292.
[11] Cui G, Qvigstad G, Falkmer S,etal. Spontaneous ECLomas in cotton rats (Sigmodon hispidus): tumours occurring in hypoacidic/hypergastrinaemic animals with normal parietal cells[J]. Carcinogenesis, 2000, 21(1): 23-27.
[12] Martinsen TC, Kawase S, Hakanson R,etal. Spontaneous ECL cell carcinomas in cotton rats: natural course and prevention by a gastrin receptor antagonist[J]. Carcinogenesis, 2003, 24(12): 1887-1896.
[13] Yan J, Guo X, Xia J,etal. MiR-148a regulates MEG3 in gastric cancer by targeting DNA methyltransferase 1 [J]. Med Oncol, 2014, 31(3): 879-884.
[14] Guo SL, Peng Z, Yang X,etal. miR-148a promoted cell proliferation by targeting p27 in gastric cancer cells [J]. Int J Biol Sci, 2011, 7(5): 567-574.
[15] Zheng B, Liang L, Wang C,etal. MicroRNA-148a suppresses tumor cell invasion and metastasis by downregulating ROCK1 in gastric cancer[J]. Clin Cancer Res, 2011, 17(24): 7574-7583.
[16] Wang SH, Li X, Zhou LS,etal. microRNA-148a suppresses human gastric cancer cell metastasis by reversing epithelial-to-mesenchymal transition[J]. Tumour Biol, 2013, 34(6): 3705-3712.
[17] Sakamoto N, Naito Y, Oue N,etal. MicroRNA-148a is downregulated in gastric cancer, targets MMP7, and indicates tumor invasiveness and poor prognosis[J]. Cancer Sci, 2013, 105(2): 236-243.
[18] Guo SL, Ye H, Teng Y,etal. Akt-p53-miR-365-cyclin D1/cdc25A axis contributes to gastric tumorigenesis induced by PTEN deficiency[J]. Nat Commun, 2013, 4: 2544-2554.
[19] 張玲霞, 莊坤, 張瀝, 等. 血清胃泌素-17胃蛋白酶原及幽門螺桿菌Ig-G抗體與萎縮性胃炎及胃癌的關系研究[J]. 檢驗醫學與臨床, 2014(8): 1076-1078.
[20] 毛秋卉, 劉文佳, 張學儉, 等. 血清胃蛋白酶原和胃泌素-17在胃癌篩選中的作用[J]. 國際消化病雜志, 2012(2): 85-87.
[21] Tong F, Cao P, Yin Y,etal. MicroRNAs in gastric cancer: from benchtop to bedside[J]. Dig Dis Sci, 2014, 59(1): 24-30.
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
