山西省陽高縣不同月份草地蝗蟲的數量變化研究
2014-09-11 06:22:28張夏剛武曉梅蘇軍虎周恩芳姚繼廣趙美清閆科技劉興明
草原與草坪 2014年3期
關鍵詞:研究
張夏剛,武曉梅,蘇軍虎 ,周恩芳,劉 旗,姚繼廣,趙美清,閆科技,劉興明,師 俊,杭 軍
(1.山西省牧草工作站,山西 太原 030001; 2.甘肅農業大學 草業學院/草業生態系統教育部重點實驗室/甘肅省草業工程實驗室/中-美草地畜牧業可持續發展研究中心,甘肅 蘭州 730070;3.山西省陽高縣畜牧獸醫局,山西 陽高 038100)
種群動態是種群生態的核心問題。種群動態研究種群大小或數量在時間上和空間上的變動規律。簡單地分析,即①有多少(數量或密度);②哪里多,哪里少(分布和空間結構);③怎樣變動(數量變動);④為什么這樣變動(種群數量的調節機制)。許多生態學家還把動物的多度和分布的研究視為生態學的研究對象,甚至作為生態學的定義,這就足以證明種群動態問題在生態學中的重要性[1-3]。
蝗蟲是山西各類型草地的主要害蟲之一,由于受氣候條件、食物資源、天敵及自身遺傳特性的影響,其種群發生數量表現出明顯的時空變化[4-5]。為了調查清楚山西草地蝗蟲有多少和哪里多、哪里少的問題,給草地蝗蟲防控措施提供依據,于2013年4~9月份在山西省大同市陽高縣對草地蝗蟲混合種群數量進行了調查分析研究。
1 材料和方法
1.1 研究區域概況
研究區域位于山西省大同市陽高縣。陽高縣地處山西省東北部,晉冀蒙三省交界,位于E 113°28′36″~114°6′15″與E 39°49′50″~40°31′10″。東與天鎮縣、河北省陽原縣接壤,南與渾源、廣靈以六棱山為界,西與大同市新榮區、大同縣毗連,北與內蒙古豐鎮市、興和縣隔山為鄰。地形呈窄長形,南西北三面環山,中部丘陵起伏,將全縣分成南北2個小盆地。最高海拔為2 391 m,最低海拔899 m,平均海拔1 050 m。按照農業區劃,全縣分為黃土丘陵區、土石山區、邊川峪口區、平川區、灘地區5種地形,其中,前3種占土地總面積的68%。陽高縣屬典型的內蒙古大陸性氣候,四季分明,冬季寒冷干燥,夏季短,高溫多雨,春秋兩季涼爽。氣溫變化較大。年降水量420 mm,主要集中在7、8、9月,近年降水不足300 mm;年蒸發量1 924 mm,為平均降水量的5倍多,十年九旱,年年春旱。平均日照時數為2 747 h。年均氣溫7.1 ℃,極端最高氣溫38 ℃,極端最低氣溫-37 ℃,無霜期110~135 d。年平均風速3 m/s,最大風速達4.3~4.5 m/s,年平均≥8級大風日達31 d,大風日有增無減。
陽高縣境內土壤分5個土壤類,17個亞類,42個土屬,79個土種,其中,栗鈣土和栗鈣性土面積最大,占總面積的74.9%,大部分土壤養分含量較低,N、P比例失調,質地較差。
陽高縣植被處于暖溫地帶闊葉林帶向溫帶草原帶過渡的區域,即溫帶草原地帶[6-8],全縣有天然草地3.6×104hm2,占總土地面積20.1%。山地灌叢類草地1.95×104hm2,占草地總面積的58%,主要分布在大泉山、綠園山一帶,涉及王官屯鎮、下深井鄉、大白登鎮;山地草原類草地0.87×104hm2,占草地總面積的26%,主要分布在云門山一帶,涉及長城鄉、羅文皂鎮;低濕草甸類草地0.53×104hm,占草地總面積的16%,分布在北徐屯、王官屯、龍泉鎮和獅子屯、大白登及羅文皂的部分河灘區,以白登河沿岸較為集中。
1.2 調查樣地選擇與編號
在前期工作基礎上[10,11],通過實地踏察,根據地形和植被等特點,在草原和草甸區域共劃分出8種主要生境作為調查蝗蟲數量的樣地。樣地Ⅰ 茭蒿(Artemisiagiraldii)草原;Ⅱ 百里香(Thymusmongolicus)、針茅(Stipaklemenzii)草原;Ⅲ 針茅草原;Ⅳ 達烏里胡枝子(Lespedezadavurica)草原;Ⅴ 苔草(Carexdolichostachya)草甸;Ⅵ 五花草甸;Ⅶ 狗牙根(Cynodondactylon)草甸;Ⅷ 鵝絨委陵菜(Potentillaanserina)、金戴戴(Trolliusfarreri)、水葫蘆苗(Halerpestessarmentosa)雜類草草甸[9]。
1.3 各生境的蟲口密度調查
從4月到9月,每月下旬在已選定的各個生境內抽樣調查蝗蟲混合種群密度。在每個生境用1 m×1 m×0.2 m的無底樣方框隨機抽樣12個,逐樣方捕捉并計數蝗蟲。每月調查8個生境,共計抽取96個樣方。全年調查6次,累計抽樣576個樣方。以所采集的蝗蟲標本和樣方資料作為分析研究依據[10-14]。
1.4 數據處理
對調查數據用Excel軟件分別按不同月份和不同生境進行統計處理,并進行對比分析。
2 結果與分析
研究從4月下旬開始到9月下旬,在8個不同草地類型(生境)、576個樣方中共捕獲20 701頭蝗蟲,其中最大樣方捕獲量為314頭/m2,最小為0頭/m2,平均蟲口密度為35.9頭/m2。
2.1 不同月份之間的蟲口數量、密度和變化趨勢
比較各月份的總捕獲量、蟲口密度及其月份之間的變化趨勢可以看出,(1)數量 7月和9月是2012年各月蝗蟲數量的兩個極端,7月的96個樣方中共捕獲4 860頭蝗蟲,位居各月總捕獲量之首,9月在相同數量的樣方內捕獲了1 466頭蝗蟲,是各月份捕獲量最少的,兩者之間相差3.32倍;(2)平均密度 從4月到9月各月平均蟲口密度依次為:45.9、35.0、38.1、50.6、30.6和15.3頭/m2,這個序列表明7月平均密度最大,9月平均密度最小,說明平均密度和總數量變化趨勢相同,具有密切相關性;(3)數量變化趨勢 比較各月蟲口數量變化趨勢可以看出,4月蟲口數量明顯高于5月,說明4月是蟲卵孵化的高峰期,低齡若蟲數量較多,但同時由于氣溫波動較大,低齡若蟲死亡率也比較高,進入5月數量有所下降,但從5月開始氣溫穩定回升,蟲口數量呈逐月增加趨勢,7月底達到一年中的峰值,然后到氣溫開始下降的后半年,蟲口數量呈逐月減少的趨勢,到9月達到觀測期的最低值。
陽高縣草地蝗蟲的月數量、平均密度和數量變化趨勢是該地區當年的氣溫、濕度、地形、土壤、植被等自然因素的綜合反映,也與當地蝗蟲的個體發育有關[15]。4月底大量蝗卵開始孵化出若蟲,此時是蝗蟲數量最多的時候,但剛剛孵化出的低齡若蟲死亡率比較高,因而5月的蝗蟲數量和密度則低于4月;5月以后氣溫穩定回升,有一部分蟲卵繼續孵化出新蝗蝻,加之若蟲死亡率逐漸減少,因而蟲口數量逐漸增加;到8月份,隨著雨季到來、氣溫下降,部分蝗蟲逐漸死亡,呈現出蝗蟲數量下降趨勢。
2.2 同一月不同生境之間蟲口密度的差異性分析
對調查結果按月進行統計處理(表1)可見:(1) 經F測驗結果,4、5、6、7、8月各生境之間的蟲口密度差異均達到極顯著水平(P<0.01),9月各生境之間的蟲口密度差異顯著(P<0.05);(2)與樣本的平均密度相比較,各月的各個生境樣本標準差普遍較大(接近樣本平均數),反映出樣本內各個變數與平均數離散程度較高,差異較大,說明即使同一月,同一生境的蟲口分布不均,樣方間數量差異很大;(3)標準誤差普遍較小,由于標準誤差反映的是樣本均數之間的變異程度,表明本次試驗樣本抽樣誤差較小,統計量與總體參數的值比較接近,因而各生境對整個草地均具有一定的代表性,用各生境統計量推斷整個草地蝗蟲相關參數的可靠度也比較大。因此,統計推斷有一定的可靠性。
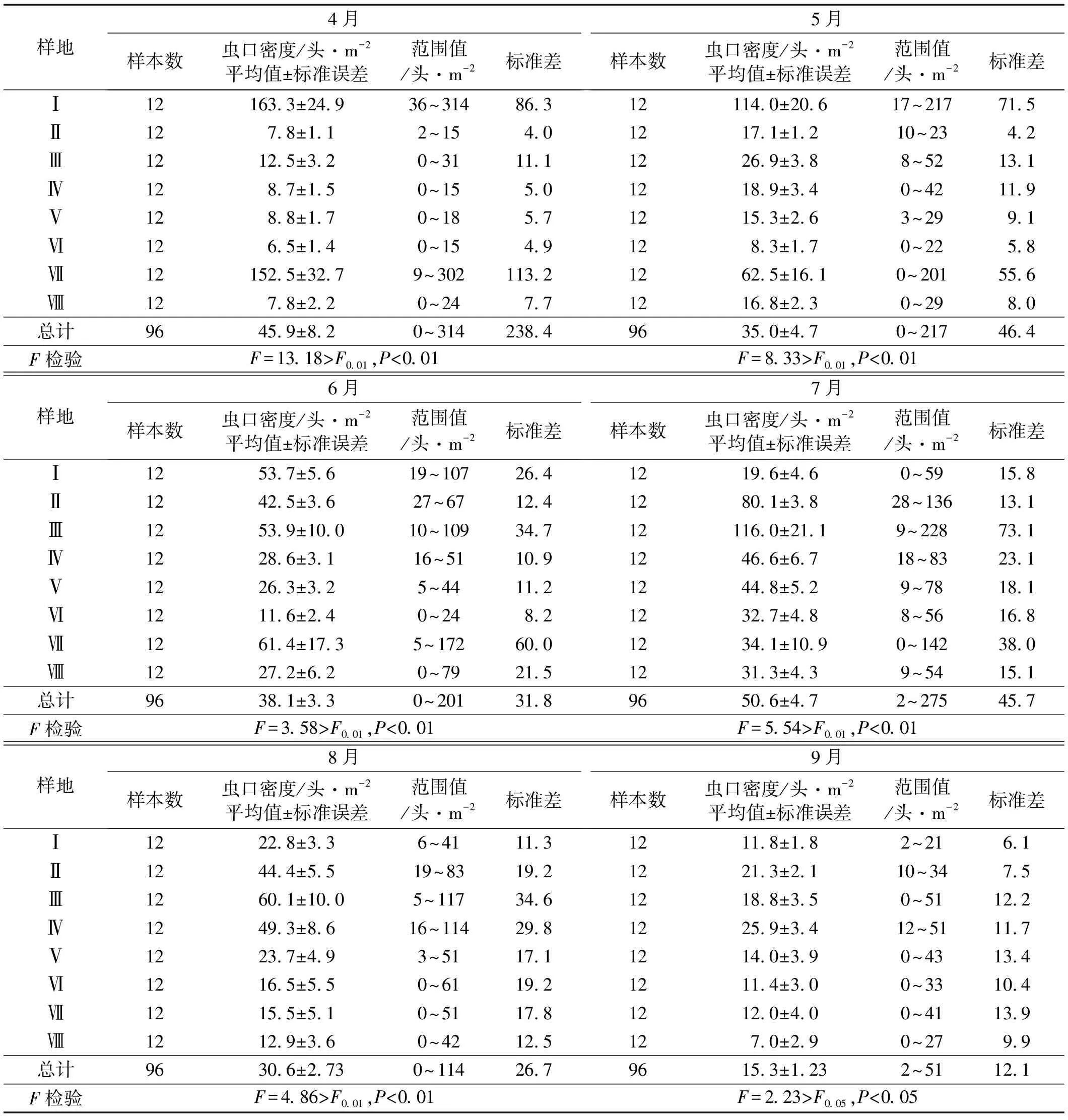
表1 同月不同生境之間的蟲口密度
2.3 蝗蟲在不同月的分布特點
把各月不同生境里的最高數量、最低數量和平均數做成分布圖(圖1~6),可以看出陽高縣草地蝗蟲在不同月里的分布生境以及生境里的分布數量。
從圖1、2分析,4、5月蝗蟲分布極為相似,其共同特點是蝗蟲幾乎只分布在茭蒿草原和狗牙根草甸中,其中,4月茭蒿草原蝗蟲數量占該月8個生境蝗蟲總數量的44.39%,狗牙根草甸蝗蟲數量占該月8個生境蝗蟲總數量的41.47%,這2個生境里的數量占8 個生境總數量的85.86%;5月茭蒿草原蝗蟲數量占該月8個生境蝗蟲總數量的40.74%,狗牙根草甸蝗蟲數量占該月8個生境蝗蟲總數量的22.33%,這兩個生境里的數量占8個生境總數量的63.07% 。4、5月蝗蟲分布的特點表明茭蒿草原和狗牙根草甸是當年蝗蝻的主要棲息地,也是前一年蝗蟲的主要產卵地和當年的繁殖地。
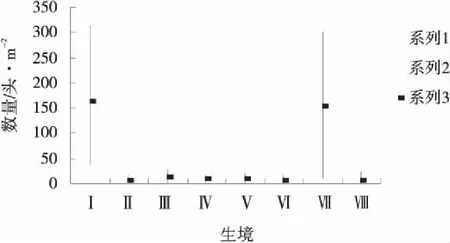
圖1 4月各生境蝗蟲分布Fig.1 Grasshopper distribution at each habitat in April
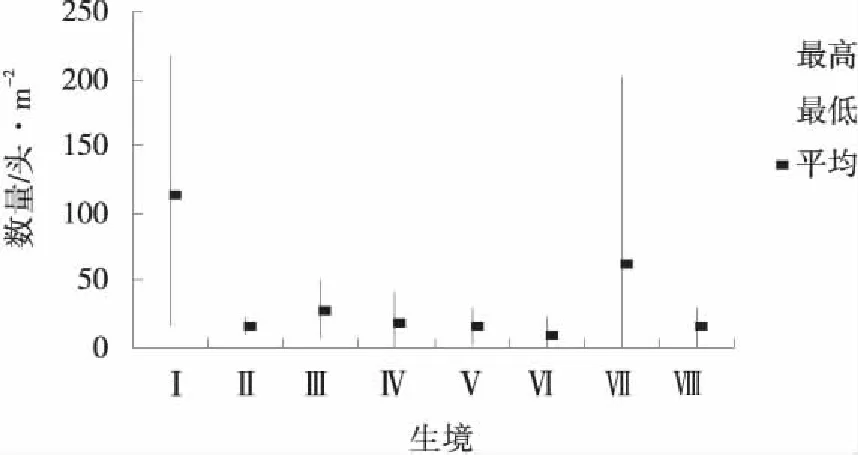
圖2 5月各生境蝗蟲分布Fig.2 Grasshopper distribution at each habitat in May
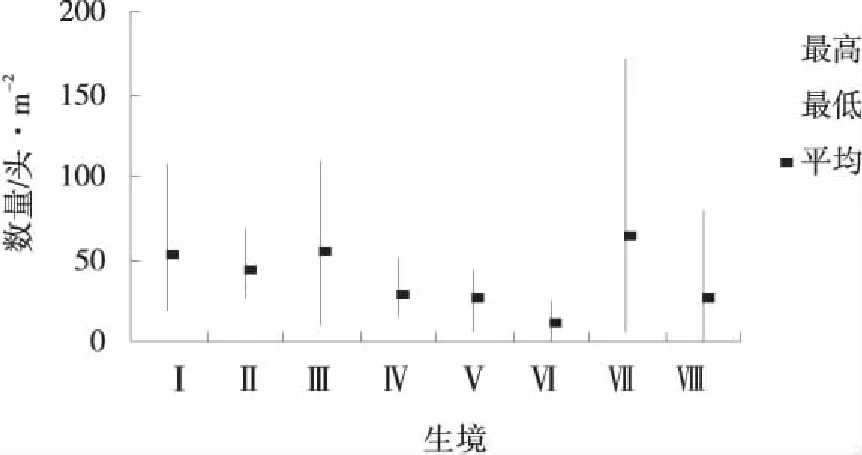
圖3 6月各生境蝗蟲分布Fig.3 Grasshopper distribution at each habitat in June
圖3~5反映出的特點是隨著蟲齡增長,蝗蟲開始從主要繁殖地逐漸向各種生境滲透,個別生境最大極值有所下降,最小極值有所提高,使得各生境間的差異縮小,但每個月都有1個數量最多或1~2個數量較多的生境,例如,6月數量最多的前3個生境依次是狗牙根草甸、針茅草原和茭蒿草原,3個生境蝗蟲數量占該月各生境總數量的55.37%;7月數量最多的前3個生境是針茅草原、百里香-針茅草原和達烏里胡枝子草原,3個生境蝗蟲數量占該月各生境總數量的59.92%;8月數量最多的前3個生境是Ⅲ、Ⅳ、Ⅱ,3個生境蝗蟲數量占該月各生境總數量的53.91%。
圖6表明9月各個生境的蝗蟲數量分布已經相當均勻,基本沒有蝗蟲數量特別多和特別少的生境。
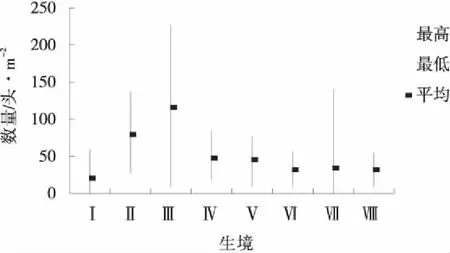
圖4 7月各生境蝗蟲分布Fig.4 Grasshopper distribution at each habitat in July
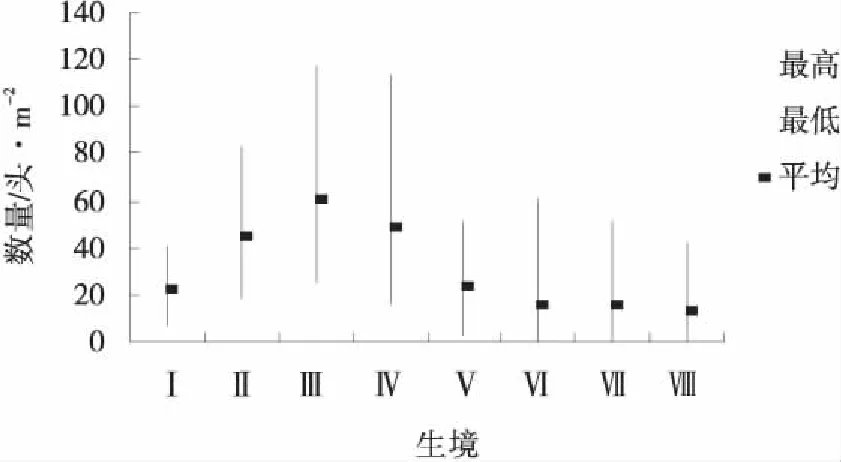
圖5 8月生境蝗蟲的種群數量統計Fig.5 Grasshopper population at each habitat in August
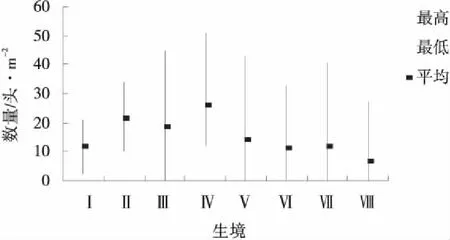
圖6 9月生境蝗蟲的種群數量統計Fig.6 Grasshopper population at each habitat in September
3 討論與結論
3.1 討論
群落組成中的每一個成份在決定整個群落的性質和功能上并不具有完全相同的地位和作用,可以依據種群在群落中的數量大小以及食物鏈中的地位等,將種群劃分為優勢種和從屬種[16],再分別探討其在生態系統中的地位和作用問題[17]。同時,在以往的許多研究工作中,用來衡量草原蝗蟲的數量指標大多都使用物種數和種群個體數量,這對正確反映各種群在生態系統中的功能時往往會增大小個體種類的作用[18]。本研究經過幾年觀察,發現陽高縣草地蝗蟲主要為混合種群,且主要由小個體種組成,為此本研究選擇混合種群個體數量為研究指標既不失其科學內涵,亦具有一定的實用性。
由于不同月份的氣溫、地溫、濕度、旱澇、地形、海拔、土壤和植被條件等各不相同,并表現出與該月蝗蟲數量變化相似的趨勢[19,20],因此,各個月的蝗蟲數量就是各月包括氣象等因子在內的綜合自然因子的函數。這是因為蝗蟲的產卵(產卵地和產卵時間)、孵化、個體發育、世代發生等生活史的各個環節都受當時當地綜合自然條件的影響所致。
在整個時序過程中,種群數量的波動受蝗蝻尤其是1-3齡若蟲影響較大,并產生數量高峰值超前,這是造成4月份的數量比5月份高的主要原因;然而,不同種類蝗蟲發育時序是有分化的,因此,5月份以后,雖然早期發生種類個體數量大量減少,但晚期發生種類個體數量逐漸增加,這就是5~7月混合種群數量還能繼續增長的原因,同時證明混合種群發生和蔓延的時序分化是非常明顯的[21-23]。
數量分析表明2012年陽高縣草地蝗蟲以茭蒿草原和狗牙根草甸為最主要的發生地,并由此逐月向外擴散和蔓延,形成了7月份的峰值和9月份的谷值,那么,在未來的年份里該區域草地蝗蟲還會不會以相同或相似模式發生和蔓延,還需要繼續跟蹤研究。
3.2 結論
(1)研究山西省陽高縣4~9月草地蝗蟲的總捕獲量和蟲口密度的結果證明:7月的數量最多、密度最大,9月的數量最少、密度最小。
(2)4月底蝗蟲數量較多,但基本都是低齡若蟲,因此,到5月底數量略有下降,從5月底到7月底蝗蟲數量呈增加趨勢,之后到9月底蝗蟲數量呈減少趨勢。
(3)逐月進行F測驗,結果表明4、5、6、7、8月各生境(樣本)之間的蟲口密度差異均達到極顯著水平(P<0.01),9月各生境之間的蟲口密度差異為顯著水平(P<0.05);各個月的樣本標準差普遍較大(接近樣本平均數),反映出樣本內各個蟲口密度與平均數之間離散程度較高,差異較大;標準誤普遍較小,表明本研究抽樣誤差較小,樣本統計量與總體參數的值比較接近,用各生境統計量推斷整個草地蝗蟲相關參數的可靠度也比較大。
(4)4、5兩個月蝗蟲群落以各齡級的若蟲為主,且幾乎只分布在茭蒿草原和狗牙根草甸,說明這兩類生境是上年蝗蟲的主要產卵地和當年的繁殖地。從6月開始蝗蟲逐漸由繁殖地向外擴散,起初高數量只出現在少數幾個生境,到7月份數量達到年峰值以后,9月份各生境里的數量逐漸接近相等,再無數量特別多或特別少的生境了。
參考文獻:
[1] 孫儒泳.動物生態學原理[M].北京:北京師范大學出版社,2001:125.
[2] 王宏平.偃師市1998 年東亞飛蝗大發生原因分析[J].昆蟲知識,1999,6(4):221.
[3] 陳永林.蝗蟲再猖獗的控制和生態學治理[J].中國科學院院刊,2000,15(5):341-345.
[4] 蘆榮勝,常巖林,秦邦才,等.山西蝗蟲名錄[J].太原師范學院學報(自然科學版),2002(1):51-55.
[5] 鄭哲民.中國蝗蟲的分類學研究[J].陜西師范大學學報(自然科學版),2003(S2):74-76.
[6] 上官鐵梁.山西主要植被類型及其分布的初步研究[J].山西大學學報(自然科學版),1985(1):36-48.
[7] 茹文明.中條山東段植被及其垂直分布的研究[J].晉東南師范專科學校學報,2000(3):11-19.
[8] 張金屯.山西蘆芽山植被垂直帶的劃分[J].地理科學,1989(4)57-63.
[9] 楊汝榮.山西省草原概況和畜牧業發展方向[J].中國草地學報,1986(1):53-58.
[10] 武曉梅,張夏剛,蘇軍虎,等.山西省晉西7~8月草地蝗蟲種的多樣性及豐富度研究[J].草原與草坪,2013(1):34-39.
[11] 李寶,武曉梅,張夏剛,等.4.5%氯氰菊酯對草原蝗蟲混合種群防效研究[J].草原與草坪,2013(2):34-39.
[12] 王俊梅,豆衛,楊自芳,等.苦參堿對草地蝗蟲種群密度的控制效果[J].草原與草坪,2008(6):23-36.
[13] 李笑碩,邢振彪,徐東,等.包頭市蝗蟲常見種類與綜合控制措施[J].華北農學報,2004(1):58-63.
[14] 李笑碩,邢振彪,徐東,等.包頭市蝗蟲常見種類與綜合控制措施[J].華北農學報,2004(1):58-63.
[15] 吳瑞芬,霍治國,盧志光,等.蝗蟲發生的氣象環境成因研究概述[J].自然災害學報,2005(6):66-73.
[16] 馮光翰.草原蝗蟲防治指標的研究[J].植物保護學報,1995(3):33-37.
[17] 孫濤,龍瑞軍,劉志云.祁連山北麓四種天然草地蝗蟲物種多樣性比較研究[J].草原與草坪,2010(6):58-62.
[18] 柳小妮,蔣文蘭,劉曉靜,等.夏河甘加草原草地蝗蟲優勢種的確定及混合種群密度高峰值模型研究[J].草原與草坪,2007(4):33-38.
[19] 胡靖,張廷偉,劉長仲,等.基于模糊聚類法的亞洲小車蝗種群空間格局分析[J].甘肅農業大學學報,2012,47(3):62-66.
[20] 鄭鳳英,張金屯,張峰,等.山西省草地資源及合理利用[J].山地研究,1996(1):27-31.
[21] 賀達漢,田真,金桂蘭,等.荒漠草原蝗蟲種群地位及時空變化的數量分析[J].1996,17(3):18-26.
[22] 王華弟,徐志宏,馮志全,等.中華稻蝗發生規律與防治技術研究[J].中國農學通報,2007,23(8):387-390.
[23] 任炳忠,楊鳳清.吉林省蝗蟲的區域分布特征[J].東北師大學報(自然科學版),1993(4):54-58.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
