藏北索縣—巴青地區中侏羅世生物礁古生態學研究①
2014-11-13 02:36:22肖傳桃柳成葉明陳肯艾軍范君
沉積學報 2014年1期
關鍵詞:生物
肖傳桃柳成葉明陳肯艾軍范君
(長江大學油氣資源與勘探技術教育部重點實驗室 武漢 430100)
0 引言
藏北索縣、巴青均位唐古拉山脈的南側,屬于藏北地層區[1],區內中侏羅世地層中發育有大量生物礁,這些生物礁系作者等發現[2,3]。研究區生物礁分別產于索縣中侏羅統桑卡拉傭組和巴青馬如布曲組中。桑卡拉傭組的下部主要發育潮坪相砂泥巖沉積,中部到上部為開闊臺地相灰巖沉積,布曲組也屬于一套正常淺海相灰巖沉積。索縣桑卡拉傭組中發現的中侏羅統生物礁與中揚子地區下奧陶統生物礁比較相似[4,5],其共同特點是數量多、分布廣,但厚度和規模較小(圖2),礁體厚大約2~7.5 m,在橫向上可見80~100 m,區域上出露范圍約10 km。生物礁多半呈面包狀、丘狀及似層狀等,與正常沉積巖層呈指狀交錯特征,且分帶比較明顯。
其中,本區雙殼類Liostrea生物礁在中侏羅統的發現,在我國雙殼類生物礁領域尚屬于首次。這一發現一方面填補了西藏北部地區生物礁領域的空缺,同時也豐富了中國侏羅系生物礁的類型,更有意義的是這個發現對于現今和以后在我國最后一塊尚未大規模勘探的大陸區域—西藏北部地區進行石油和天然氣勘探具有一定的經濟意義。
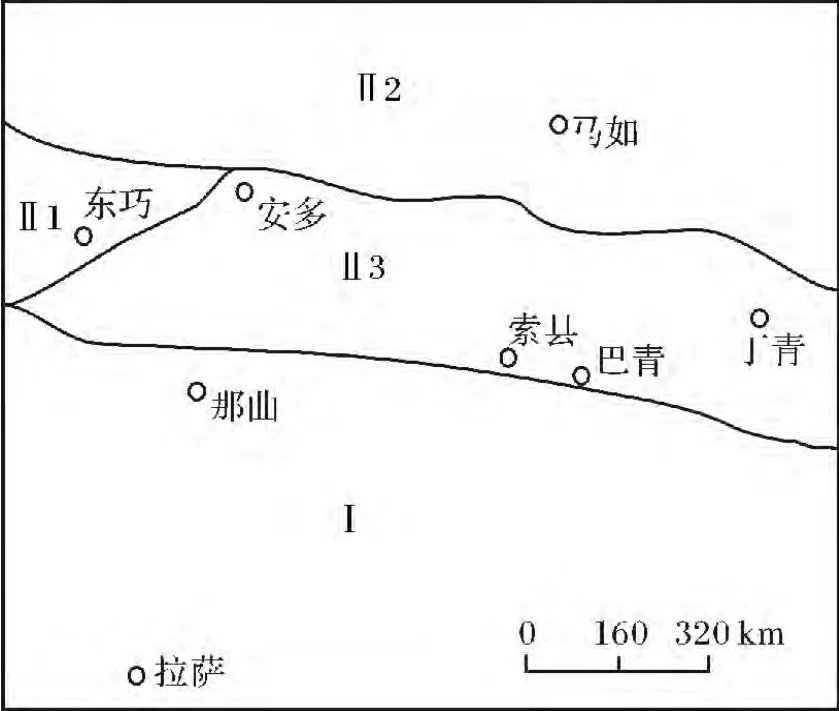
圖1 研究區地層區劃(引自文獻[1])Fig.1 Stratigraphic divisions of the study area
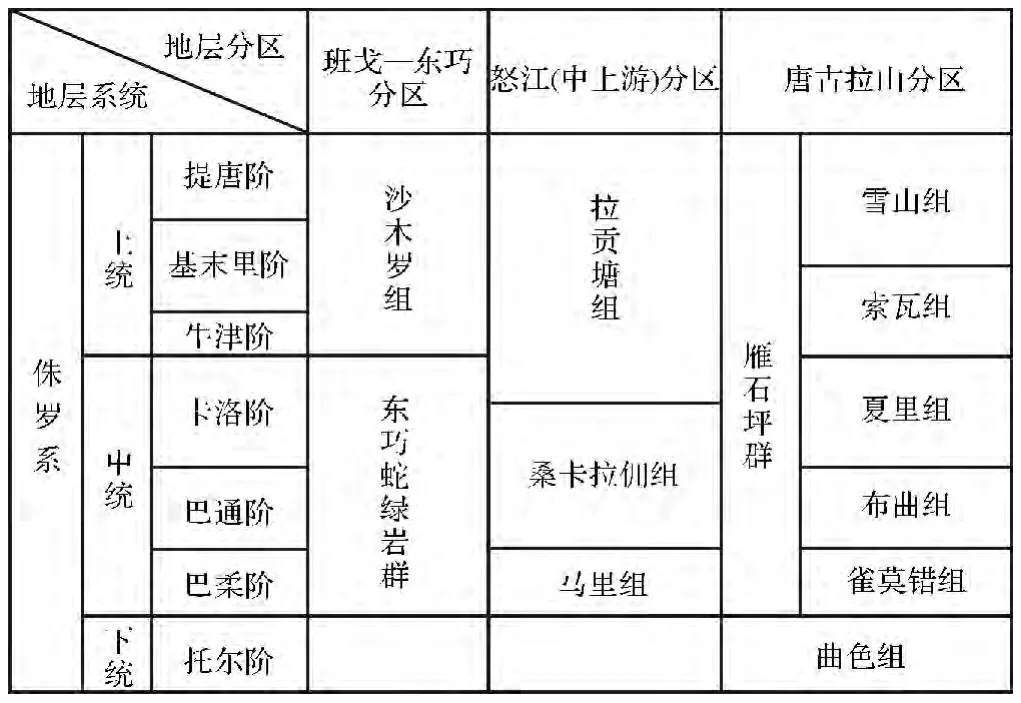
表1 藏北地區中—東部侏羅紀地層系統Table 1 Jurassic strata system of the middle-east part of North Tibet
1 區域地層發育特征
藏北索縣、巴青及安多地區均位唐古拉山脈的南側,屬于藏北地層區(表1),其中,索縣桑卡拉傭組中產有腕足類:Holcothyris elliprica,Futchiyris lingularis;層孔蟲 Parastromatopora memoria-naumarmi,P.sp.以及珊瑚Schizosmilia rollier等。其中Holcothyris elliprica,Holcothyris是中侏羅世標準化石,是著名的B-H動物群的命名分子,而且主要限于中侏羅世中期(巴通期)[6],Parastromatopora memoria-naumarmi分布較泛,見于歐洲、日本以及伊朗等地中上侏羅統[7~19]。布曲組中產有腕足類Burmirhynchia cuneata,B.trilobata,Holcothyris golmudensis以及 H.fleas等、雙殼類Liostrea birmanica,L.sublamellos和 Camptonectes(C.)lens等。這些化石均為中侏羅世標準化石,并構成了Burmirhynchia-Holcothyris組合帶,而且主要限于中侏羅世中期(巴通期),它們曾見于雙湖地區、雁石坪地區的布曲組、滇西、藏東的柳灣組、和平鄉組以及緬甸的南瑤組(Namyao Series)[5]。因此,本生物帶的地質時代應屬中侏羅世中期。
2 造礁生物的類別和生態特征
研究區的中侏羅世造礁生物類型以層孔蟲為主,其次為雙殼類及六射珊瑚等。在造礁生物中,層孔蟲宏觀形態為柱狀,而六射珊瑚則以叢狀群體為主,雙殼類單體屬于固著生活的生態類型,盡管上述三種生物均為造礁生物,但它們的生態特征均有差異。
2.1 層孔蟲的生態特征
層孔蟲主要見于索縣城東桑卡拉傭組,其形態呈柱狀或筒狀為,以 Parastromatopora memoria~naumanni為代表,其直徑1~1.5 cm,長5~8 cm不等。
層孔蟲屬于海洋底棲固著生活的動物(圖版IA,B),全部都是群體,沒有單體。層孔蟲的硬體構造比較復雜,具有固著功能的星根狀構,其主體部分包括有共骨、軸柱、蟲室、中柱以及泡沫組織和骨素等構造[2],層狀構造很發育,它是層孔蟲不同階段生長的產物。層孔蟲中群居的很多軟體個體都位于眾多的蟲室內,這些軟體的個體都能分泌鈣質的骨骼。層孔蟲這類動物一般固著于較硬的底質上生活,其攝食方式主要靠從流動的水體中濾食有機質或者微生物等,由其硬體構造和造礁功能推測其生態環境應為光線、氧氣充足、清潔、溫暖、且水體循環較好的正常淺海。由于筒狀層孔蟲的共骨結構較為結實,且能夠形成較堅實的抗浪格架,能夠起到障積灰泥或者碳酸鹽顆粒的作用,因此,其生態環境推測為水體較淺、能量相對較高的環境,筒狀層孔蟲共骨間充填有一定數量的粉屑和生物碎屑等現象說明了此點。
2.2 六射珊瑚的生態特征
六射珊瑚以 Schizosmilia rollieri為代表,主要出現在索縣桑卡拉傭組生物礁中,該類珊瑚呈叢狀復體,個體橫截面呈圓—橢圓形(圖版I-C,D,E),直徑25~30 mm,珊瑚體腔內具有兩級隔壁,其一級隔壁最長,可相交于體腔中心,二級隔壁較短,大約為為一級隔壁的2/3~4/5。在露頭中六射珊瑚多呈原始生長狀態保存,其生態功能主要是原地固著生長,并形成抵抗波浪的格架,障積灰泥及各類碳酸鹽的顆粒,最終形成了障積礁或骨架礁。
群體六射珊瑚是一類典型底棲固著型動物,和層孔蟲類似,其群居的許多軟體個體分別位于眾多的蟲室內,每個蟲室容納一個小的個體。其捕食方式和層孔蟲類似,靠其口周圍的一圈或多圈觸手的來回擺動,引起海水流動并進入其口中,之后濾食海水中所攜帶的有機質、微生物和養分等。六射珊瑚所要求的生態環境與層孔蟲較為相似,生活于溫暖、清潔、能量相對較高,光線和氧氣充足,水體循環較好的正常淺海中。
2.3 雙殼類的生態特征
研究區造礁的雙殼類只發育于巴青縣馬如鄉一帶的布曲組中[2],且是布曲組中主要的造礁生物,其特征主要表現為Liostrea(光蠣)一屬的大量繁盛,該類生物呈單體、且近似于牡蠣狀、具有左右兩瓣殼的動物,左、右兩殼就是其硬體,其兩瓣殼較厚,軟體位于左右兩殼之內,其左殼一般呈膨凸較強,右殼則比較扁平(圖版I-F,G,H),這種特殊的形態特征是由于Liostrea長時間以左殼固著于較硬的海底上生活,右殼已經特化為口蓋演化而成的。
Liostrea一般以左殼固著于較硬的海底上生活,利用其扁平右殼的頻繁地張、合作用而將帶有微生物及有機質的海水引入其殼內,然后從流動的水體中濾食有機質等食物為生,但是,當海水不清潔或渾濁時,其右殼便作為口蓋而蓋于左殼之上以保護其軟體組織[2]。由Liostrea生物特化的硬體構造等推測,該類生物能夠適應于不同鹽度和不同渾濁度的淺海環境,但當其生態環境為溫暖、清潔、光線和氧氣充足的正常淺海時,它們便能大量固著生長和繁殖,形成生物礁。
3 造礁生物群落劃分
造礁群落是群落最典型的體現,因為造礁群落中的大部分生物大都是固著生活的,其中的生物均有各自的生態位,每個群落具有較清晰的邊界,而死亡后的造礁生物群均屬于典型的原地埋葬生物群,所以,對造礁群落的研究具有重要的古生態學意義。本文通過對研究區造礁群落的研究,不僅可以用于古環境的分析,而且對于研究區的古氣候、古地理及古大地構造的研究提供重要的依據。根據生物礁內部造礁生物的生長階段性特征以及造礁生物的組合特征,本文將索縣桑卡拉傭組生物礁劃分為3個造礁群落,巴青布曲組生物礁也劃分為3個造礁群落(表1)。
3.1 Schizosmilia-Parastromatopora群落
該群落主要見于索縣中侏羅統柳彎組中,即圖2中的A群落,其下為生物礁的礁基,由生物碎屑灰巖組成,之上為Parastromatopora群落,這兩個群落間的邊界較清晰。
組成:本群落的組成分子主要有六射珊瑚Schizosmilia rollieri,S.sp 等和柱狀層孔蟲 Parastromatopora memoria-naumanni P.compacta,P.sp.。
結構與功能:在該群落中,Schizosmilia為優勢分子,其含量約為60% ~70%,在地層中多呈垂直于層面或者傾斜保存狀態,少數化石呈水平狀態。Schizosmilia的生態功能是在原生態狀態條件下固著生長,障積灰泥和碳酸鹽顆粒,該生物是本群落中的建設者,為群落的生長造就了穩定的生活環境。Parastromatopora memoria-naumann為本群落的特征分子,同時也構成了亞優勢分子,豐度為20% ~30%,其生態功能與Schizosmilia功能大體相同。塊狀的層孔蟲Milleporella pruvosti在本群落中含量很少。腕足類Tubithyris globata以及雙殼類Lopha solitaria,Chlamys baimaensis為本群落的居礁生物,他們基本屬于固著生活。
生態環境分析:在該群落中,營固著底棲方式生活的生物占100%,六射珊瑚Schizosmilia、層孔蟲Parastromatopora直接固著海底生活,腕足類Tubithyris以肉頸附著海底,Lopha以殼體固著,Chlamys可能以足絲附著生活。這些固著生活的生物靠流動的水體帶來營養和食物為生。由于六射珊瑚Schizosmilia屬于叢狀復體,其抵抗水動力強度較大,但由于Parastromatopora的柱體直徑1~1.5 cm,因此,很難抵抗較強的水動力條件,綜合以上因素,本文認為本群落的生態環境可能為光線、氧氣充足、清潔、溫暖、循環較好以及水體略深的正常淺海,水體深度約10~20 m。
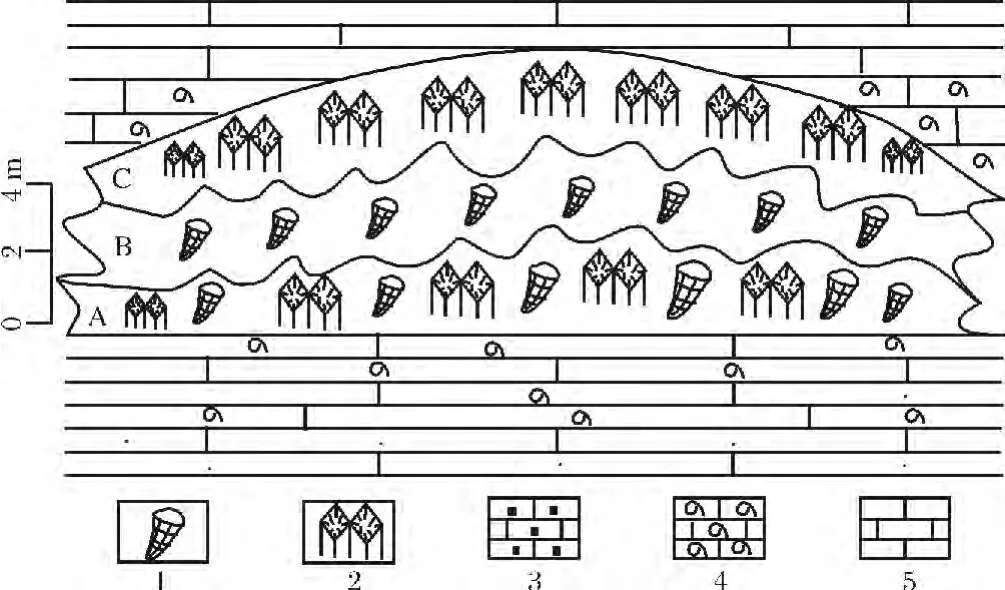
圖2 索縣城東中侏羅世桑卡拉傭組生物礁分布特征(據文獻[11]修改)Fig.2 Reef distribution of Liuwan Formation from Middle Jurassic in the eastern of Suoxian county
3.2 Parastromatopora群落
本群落主要見于索縣中侏羅統柳彎組中(圖版IA),即圖2中的B群落,群落的頂、底邊界清晰,之下為Schizosmilia-Parastromatopora群落或其形成的障積礁,之上為Schizosmilia群落或其形成的障積礁。

圖版Ⅰ說明:A、B.Parastromatopora memoria~naumanni,橫切面。A.索縣中侏羅統桑卡拉傭組層孔蟲障積巖巖。B.顯微照片,×3,橫切面。C、D.Schizosmilia rollieri,C.顯微照片,×3,橫切面,D.Schizosmilia rollieri障積巖,索縣中侏羅統桑卡拉傭組。E.Schizosmilia rollieri,索縣中侏羅統桑卡拉傭組。F、H.Liostrea障積巖,野外照片,巴青馬如布曲組。G.Cyanobacteria-Liostrea黏結—障積巖,野外照片,巴青馬如布曲組
組成:該群落主要由柱狀層孔蟲組成,如Parastromatopora memoria-naumarmi,P.compacta,P.sp.等,其次為腕足類 Tubithyris globata,T.whathyensis,Kutchithyris pingqua等。
結構與功能:在本群落中,Parastromatopora memoria-naumarmi占絕對優勢,其豐度為群落的80%,并構成群落的優勢分子,同時亦為特征分子,其次為P.compacta。它們多數呈垂直層面的直立狀態保存,平臥者很少。其生態功能是原地固著生活,障積和捕獲灰泥,給群落創造了穩定的生態環境,與前一群落相比,本群落的構架相對較小。
生態環境分析:本群落也屬于較典型的底棲固著生態型,營底棲固著型生物近于100%,即以柱狀層孔蟲Parastromatopora大量繁盛為特色,它們多數呈垂直層面的直立狀態保存,平臥者較少,由于本群落的空間構架相對較小,加之因Parastromatopora的柱體具有較小的直徑1~1.5 cm,因此,本群落難以抵抗很強的水動力,可能適應于能量中等—低能條件的生態環境,因此,其生態環境應為光線、氧氣充足、清潔、溫暖、循環較好以及水體稍深的正常淺海,水深約15~25 m。
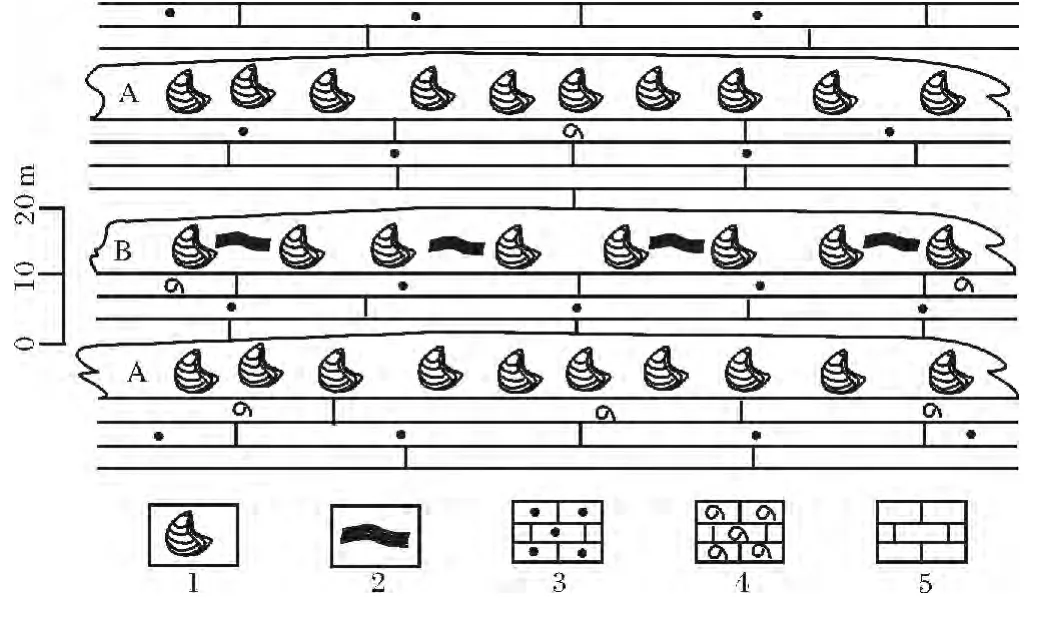
圖3 巴青馬如地區中侏羅世布曲組生物礁分布特征Fig.3 Reef distribution of Buqu Formation from Middle Jurassic in Maru area of Baqing county
3.3 Schizosmilia 群落
該群落主要見于索縣中侏羅統柳彎組中,即圖2中的C群落,群落的頂、底邊界清晰,之下為Parastromatopora群落或其形成的障積礁,其上為灰色薄至中層狀泥晶灰巖組成的礁蓋。
組成:本群落的組成分子主要為六射珊瑚Schizosmilia rollieri,S.sp 等和腕足類 Holcothyris elliptiyris,Kutchiyris lingularis。
結構與功能:在該群落中,Schizosmilia為優勢分子,其豐度約為50% ~60%,在地層中多呈垂直于層面或者傾斜保存狀態,少數化石呈水平狀態。其生態功能是在原生態狀態條件下固著生長,障積灰泥和碳酸鹽顆粒,該生物是本群落中的建設者,為群落的生長造就了穩定的生活環境。Holcothyris elliptiyris和Kutchiyris lingularis為居礁生物,豐度約為20% ~30%,但他們也是營固著生活。
生態環境分析:在該群落中,營底棲固著型生物為100%,六射珊瑚Schizosmilia直接固著海底生活,腕足類Holcothyris和Kutchiyris以肉頸附著海底,其個體較大,殼體較厚等特征說明其適應的水體較淺。這些固著生活的生物靠流動的水體帶來營養和食物為生。由于六射珊瑚Schizosmilia屬于叢狀復體,其復體呈圓形放射狀,直徑可達25~30 cm,能夠抵抗強度較大的水動力,因此,能夠適應較淺的水體,綜合上述因素認為,本群落代表的生態環境應為溫暖、清潔、氧和光線較為充足、循環較好水體稍深的正常淺海,水深約10~15 m。
3.4 Liostrea 群落
本群落主要分布于巴青縣馬如鄉雁石坪群布曲組下部和上部,即圖3中的A群落,與周圍沉積巖層邊界清晰,其下為亮晶生物碎屑組成的礁基,其上泥晶灰巖組成的礁蓋。
組成:組成本群落的生物主要有Liostrea birmanica,L.sp., Protocardia stricklandia,Camptonectes(Camptonectes)riches,P.cf.stricklandia 等。
結構與功能:Liostrea既是本群落的特征分子,同時構成了該群落的優勢分子,其含量占80% ~90%,該生物以左殼固著于海底生長,在地層中多數為雙殼一起保存,在群落中起著建設作用,并為該群落造就了穩定的生活環境。Camptonectes(Camptonectes)riches由于含量少,屬于偶見分子,其豐度為4%,該生物的足絲凹口較明顯,說明其生活方式是以足絲附著于較硬的海底或附著于Liostrea殼上生活,由其生活方式可推測為它屬于造礁生物。Protocardia則屬于移動底棲生活方式,其豐度為6% ~10%,屬于居礁生物。
生態環境分析:在本群落中,營固著底棲生活的生物約占90%以上,且以Liostrea占絕對優勢,其次為少量的Camptonectes(Camptonectes)riches,形成了雙殼類的障積礁,Liostrea具有殼體較厚、中等大小等特征,上述特征說明了該類生物能夠適應水動力較強的海洋環境,因此,推測該群落生態環境應為循環較好、中等能量、溫暖和清潔的正常淺海,海水深約約15~25 m。
3.5 Cyanobacteria-Liostrea 群落
該群落主要分布在巴青縣馬如鄉雁石坪群布曲組中部,即圖3中的B群落,與周圍沉積巖層邊界清晰,其下為亮晶生物碎屑與砂屑灰巖灰巖組成的礁基,其上為泥晶灰巖組成的礁蓋。
組成:該群落的組成分子主要有Liostrea eduliformis,L.sublamellose,Protocardia hepingxiangensis,Camptonectes(Camptonectes)lens,Pholadomya socialis qinghaiensis和Pseudetrapezium cordiforme以及藍細菌Cyanobacteria等。
結構與功能:在本群落中,以Liostrea一屬大量繁盛為特征,是本群落的優勢分子,其含量占生物的70%~80%,該生物以左殼固著于海底生長,在地層中多數為雙殼一起保存,在群落中起著建設作用,并為該群落造就了穩定的生活環境。Cyanobacteria為本群落的特征分子,其含量約為15% ~20%,其作用是黏結Liostrea以及灰泥等作用,在群落中也起到建設作用。Pholadomya含量較少,為該群落的偶見分子,含量約為2%,主要營移動底棲方式生活。
生態環境分析:在本群落中,以固著底棲方式生活的生物為主,約占70% ~85%,其中以Liostrea生物占絕對優勢,它多半適應于動力較強的海水環境,而藍細菌Cyanobacteria的出現說明海水需要一定的光照條件。所以,該群落可能代表光線和氧氣充足、溫暖和清潔、水體循環較好的潮間帶環境,水深大約5~15 m。
4 造礁群落的演化
群落的演化形式與生物礁的發育有著密切的關系,群落演化存在兩種演化形式,即演替與取代現象,群落演替現象多發生于環境漸變條件下,與之對應的生物礁更替多表現為生物礁的連續生長與發育,而群落取代現象則多發生于環境突變條件下,與之對應的生物礁發育多表現為生物礁演化的階段性變化。
4.1 索縣地區造礁群落的演化
索縣地區生物礁主要見于桑卡拉傭組中,桑卡拉傭組大致可以分為兩段,下段為碎屑巖與碳酸鹽巖混合沉積,上段為顆粒灰巖、泥晶灰巖及生物障積巖沉積,這兩段分別經歷了一個海平面變化旋回。研究區生物礁發育于桑卡拉傭組的上段,且生物礁的演化一般分為三個階段,即奠基階段、發育階段和衰亡階段,在奠基階段中形成礁基,沉積物多表現為亮晶砂屑灰巖及生屑灰巖,在生物礁的發育階段中,造礁群落演化具有連續性(表2),即存在演替系列,反應了生物礁的發育與海平面的變化之間基本保持同步。
在桑卡拉傭組一段形成之后,本區進入了第二個海平面變化旋回,同時,生物礁的發展進入了生長初期階段,由于海平面的廣泛上升,抑制了碎屑物質向盆地的帶入,在研究區形成了一套以泥晶灰巖、生物礁灰巖沉積;又因本區處于熱帶或亞熱帶地理位置,海平面上升導致了造礁生物如六射珊瑚和柱狀層孔蟲等得以大量繁盛,并形成了本區生物礁的第一個造礁群落,即Schizosmilia-Parastromatopora群落,由于造礁生物Schizosmilia和Schizosmilia的大量原地固著生長,從而形成了障積灰泥的抗浪格架,同時完成了本區生物礁發育初期階段,并形成了Schizosmilia-Parastromatopora障積骨架礁。
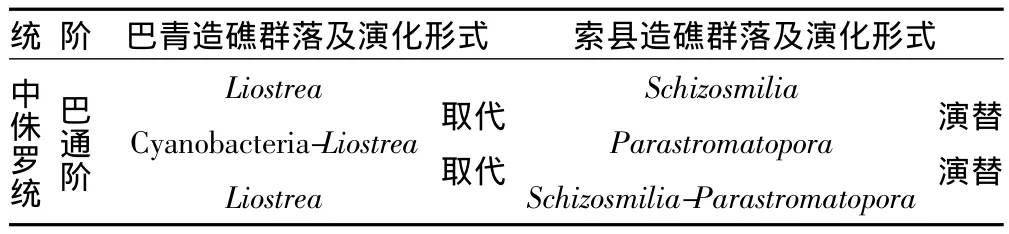
表2 索縣—巴青中侏羅世巴通期造礁群落及演化序列Table 2 Bathonian Reefs-bearing communities of Middle Jurassic and their evolution series of Suoxian-Baqing area
隨著時間的推移,本區海平面逐漸緩慢上升,隨著水體能量的不斷降低,六射珊瑚Schizosmilia逐漸減少,并以柱狀層孔蟲Parastromatopora大量發育為特征,最終形成以Parastromatopora為主體的造礁群落,并完成了 Schizosmilia-Parastromatopora群落向Parastromatopora群落的演替過程,同時也完成了本區生物礁發育中期階段,并形成了Parastromatopora障積骨架礁。之后,由于本區海平面有逐漸下降,水體逐漸變淺,能量逐漸增高,從而導致抗浪能力較弱的柱狀層孔蟲Parastromatopora逐漸減少,而抗浪能力較強的六射珊瑚 Schizosmilia大量發育,形成了Schizosmilia造礁群落,同時也完成了本區生物礁發育晚期階段,并形成了Schizosmilia障積骨架礁。在此之后,由于本區海平面的較快速的上升,從而導致生物礁生長速跟不上海平面的上升速率而衰亡,使得生物礁的發育進入了衰亡階段。
4.2 巴青地區造礁群落的演化
巴青地區生物礁主要見于布曲組中,布曲組主要為顆粒灰巖、泥晶灰巖及雙殼類障積巖沉積,其中的生物礁的演化大致為三個階段,即奠基階段、主要發育階段和衰退階段。與索縣地區相似,在奠基階段過程中以形成礁基為特點,但本區生物礁的發育階段中造礁群落的演化不具有連續性(表2),即不存在演替系列,而表現為群落的取代現象,反應了生物礁的發育與海平面的變化的非同步性。
在雀莫錯組沉積之后,本區進入了布曲組的發育時期,該組大體上包括了3個海平面變化旋回,每一個海平面變化旋回發育了一次生物礁事件沉積,每一個造礁事件表現為一次造礁群落的發育和繁殖。第一次海平面上升晚期,導致了造礁生物Liostrea的廣泛發育,且固著生長,形成Liostrea造礁群落,它們障積灰泥形成抗浪格架,之后由于海平面的進一步上升而結束了生物礁的生長,第一次生物礁發育進入了衰亡階段。在此之后,本區布曲組進入第二個海平面變化旋回,同時,發生了第二次生物礁繁殖事件,由于海平面的廣泛上升,在研究區形成了一套以泥晶灰巖、雙殼類障積灰巖沉積,又因本區處于熱帶或亞熱帶地理位置,海平面上升導致了造礁生物如Liostrea和藍細菌(Cyanobacteria)的大量繁盛,并形成了本區的第二個造礁群落,即Cyanobacteria-Liostrea群落,由于造礁生物Liostrea的大量原地固著生長以及Cyanobacteria的發育,從而形成了黏結—障積灰泥的抗浪格架,并形成了Cyanobacteria-Liostrea黏結—障積骨架礁,之后由于海平面的進一步上升而結束了生物礁的生長,使得第二次生物礁的發育進入了衰亡階段。
同樣的方式導致本區第三次造礁事件的發生,形成了Liostrea障積骨架礁。值得指出的是,本區由于三次造礁事件都是獨立發育的,期間存在著環境的顯著變化,因此,布曲組造礁群落之間關系與索縣桑卡拉傭組造礁群落之間關系不同,即布曲組中不存在連續的群落演替關系,而表現為取代關系。
5 生物礁的發育特征與成礁方式
不同時期生物礁的發育特征不同,不同類型生物礁的發育特征也不盡相同,不僅如此,生物礁的成礁方式也大不相同,這主要體現于生物礁的內部構筑方式上。
奠基階段是生物礁賴以發展的基礎,具體表現為各類生物礁的礁基由亮晶顆粒碳酸鹽巖組成,說明了生物礁都是在能量較高的生物灘或砂屑灘硬底基礎上發育起來的。
生物礁的發育階段是礁核的主要發育時期,其特征是造礁生物的快速和大量繁盛,由于生物礁類型的差異,研究區生物礁的發育階段表現為不同的成礁作用方式,分別表現為造礁生物的鈣質分泌作用、灰泥捕獲作用、生物原地堆積作用以及生物的障積作用等。在索縣地區,生物礁在早期發育階段和晚期發育階段以造架作用為其主要成礁作用方式(分別形成六射珊瑚—柱狀層孔蟲骨架礁與六射珊瑚骨架礁),并伴以造架生物的原地堆積、自身分泌和捕獲等成礁方式;在中期階段則以障積作用為主要成礁方式,其次為捕獲灰泥的作用(形成柱狀層孔蟲障積礁)。在巴青地區,生物礁發育階段的早期與晚期以固著族生的雙殼類障積作用為主成礁方式,中期階段中出現了藍細菌等低等藻類,以黏結—障積作用為其成礁方式。
衰退階段是生物礁發育的消亡期,表現為造礁生物迅速減少,或完全消失。生物礁衰亡的因素多表現為兩種情況。第一種可能是由于生物礁生態環境水體變淺,海水能量明顯加強,致使造礁生物難以適應高能環境而大量減少甚至消失,并被淺灘環境的礁蓋所取代;第二種可能是由于生物礁的生存環境快速變深,水體的光線和氧的含量、溫度、光照條件等環境因子均不適合造礁生物的生存,從而使得造礁生物迅速減少或完全消失。本區的生物礁的衰亡多屬于第二種情況。
6 結論
通過對西藏索縣—巴青地區中侏羅世生物礁的古生態學研究,本文得出如下認識:
(1)本區中侏羅世地層中,以層孔蟲為主要造礁生物,其次為六射珊瑚和雙殼類。其中,層孔蟲外形較為單調,以柱狀為主要類型,這些造礁生物多以原生態條件的狀態保存,它們均均要求光和氧氣充足、溫暖、清潔、循環良好得正常淺海環境。
(2)造礁生物組合分析表明,研究區晚侏羅世生物礁可以劃分為5個造礁群落,它們是Schizosmilia-Parastromatopora,Parastromatopora,Schizosmilia,Liostrea和Cyanobacteria-Liostrea造礁群落,各群落的組成、結構與功能以及生態環境特征分析表明,盡管造礁群落的所要求生態環境大體相似,但它們要求的水體具體深度可能存在一定的差異性。
(3)造礁群落演化分析表明,群落演化表現為取代兩種形式,第一種形式為群落演替,這種形式主要發育于索縣桑卡拉傭組生物礁中,主要由于生態環境處在漸變條件下,從而導致生物礁內部造礁生物之間的相互競爭而完成群落的更替。第二種形式為群落取代,這種形式主要發育于巴青布曲組生物礁中,主要是由于相對海平面和生態環境的明顯變化所造成,由于海平面的相對較快速的上升,導致生物礁的生長速率跟不上海平面的變化,故而導致生物礁的衰亡,從而導致群落的更替。
(4)對生物礁的發育特征與成礁方式研究認為,研究區主要成礁方式有造架作用、障積作用和黏結—障積作用,其中前兩者主要體現于桑卡拉傭組生物礁中,后兩種主要見于巴青布曲組生物礁中。
致謝 參加野外工作的還有龔文平、李藝斌、胡明毅、肖安成等,在此表示衷心的感謝。
References)
1 肖傳桃,夷曉偉,李夢,等.藏北安多東巧地區晚侏羅世生物礁古生態學研究[J].沉積學報,2011,29(4):752-760[Xiao Chuantao,Yi Xiaowei,Li Meng,et al.A study on palaeoecology of Late Jurassic reefs in Dongqiao area in Anduo county of northern Tibet[J].Acta Sedimentologica Sinica,2011,29(4):752-760]
2 肖傳桃,李藝斌,胡明毅,等.藏北地區侏羅紀生物礁的發現及其意義[J].中國科學基金,2000,14(3):178-182[Xiao Chuantao,Li Yibin,Hu Mingyi,et al.Discovery and geologic significance of Jurassic organic reef in northern Tibet[J].Chinese Science Foundation,2000,14(3):178-182]
3 肖傳桃,李藝斌,胡明毅,等.西藏安多縣東巧晚侏羅世生物礁的發現[J].地質科學,2000,35(4):501-506[Xiao Chuantao,Li Yibin,Hu Mingyi,et al.Late Jurassic reefs in Dongqiao area of Anduo,Xizang(Tibet)[J].Scientia Geologica Sinica,2000,35(4):501-506]
4 肖傳桃,姜衍文,劉秉理,等.中揚子地區早奧陶世早中期Batostoma屬的發現及其地質功能和生態學研究[J].科學通報,1993,38(14):1314-3116[Xiao Chuantao,Jiang Yanwen,Liu Bingli,et al.A discovery of Batostoma genus of Early-Middle Age of Early Ordovician and a study on its geologic function and ecology in Middle Yangtze area[J].Chinese Science Bulletin,1993,38(14):1314-1316]
5 Xiao C,Li M,Yang W,et al.Palaeoecology of Early Ordovician reefs in the Yichang area,Hubei—A correlation of organic reefs between Early Ordovician and Jurassic[J].Acta Geologica Sinica,2011,85(5):1003-1015
6 肖傳桃,龔文平,胡明毅,等.藏北地區中部侏羅紀生物地層層序[J].江漢石油學院學報,2004,26(4):1-5[Xiao Chuantao,Gong Wenping,Hu Mingyi,et al.Jurassic biostratigraphic sequence of central northern Tibet[J].Journal of Jianghan Petroleum Institute,2004,26(4):1-5]
7 董得源,汪明洲.藏北安多一帶晚侏羅世層孔蟲的新材料[J].古生物學報,1983,22(4):413-427[Dong Deyuan,Wang Mingzhou.New materials of the Upper Jurassic stromatoporoids in the Anduo county of North Xizang[J].Acta Palaeontologica Sinica,1983,22(4):413-427]
8 汪明洲,董得源.藏北東巧層孔蟲[J].古生物學報,1983,23(3):343-348[Wang Mingzhou,Dong Deyuan.Stromatoporoids from the Dongqiao Formation in Northern Tibet[J].Acta Palaeontologica Sinica,1983,23(3):343-348]
9 Hudson R G S.Jurassic Stromatoporoids from the Lebanon.J.Paleont.1954a,28,657-661
10 H.Yabe,T.Sugiyama.Jurassic Stromatoporoids fromJapan.Sci.Rept.Tohoku Imp.Univ.Japan Ser.1935,2(14):4-56
11 Hudson R G S.Tethyan Jurassic Stromatoporoids Stromatoporina,dehornella and Astroporina[J].Palaeontology,1960,(2),180-199
12 Kopaska-Merkel D C,Schmid,D U.New bioherm-building tubular organism in Jurassic Smackover Formation,Alabama:Transactions of the Gulf Coast Association of Geological Societies,1999b,49:300-309
13 Fursich F T,Werner W.Palaeoecology of calcisponge-coral meadows from the Upper Jurassic of Portugal.-Pal&aumlont.Z.,1991,65,35-69,Stuttgart
14 Kano A,Kakizaki Y,Takashima C,et al.Facies and depositional environment of the uppermost Jurassic stromatoporoid biostromes in the Zagros Mountains of Iran.GFF,2007,129:107-112
15 Reinhold R.Leinfelder,Felix Schlagintweit,Winfried Werner,et al.Significance of stromatoporoids in Jurassic reefs and carbonate platforms-concepts and implications[J].Facies,2005,51:287-325
16 Akihiro Kano,Yoshihiro Kakizaki,Chiduru Takashima,et al.Facies and depositional environment of the uppermost Jurassic stromatoporoid biostromes in the Zagros Mountains of Iran.GFF 2007,129:107-112
17 Kershaw S,Keeling M.Factors controlling the growth of stromatoporoid biostromes in the Ludlow of Gotland,Sweden[J].Sediment Geology,1994,89:325-335
18 Eric Fookes,Geneva.Development and eustatic control of an Upper Jurassic reef complex(Saint Germain-de-Joux,Eastern France)[J].Facies,1995,33:129-150
19 Markus Jank ,Andreas Wetzel,Christian A.Meyer.Late Jurassic sealevel fluctuations in NW Switzerland(Late Oxfordian to Late Kimmeridgian):closing the gap between the Boreal and Tethyan realm in Western Europe,Original Article Facies,2006,52:487-519
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
