糖鏈長度對α-乳白蛋白-糖復合物抗原性的影響
2014-11-20 12:10:58范金波蔡茜彤侯宇周素珍馮敘橋
食品與發酵工業 2014年10期
范金波,蔡茜彤,侯宇,周素珍,馮敘橋
(渤海大學食品科學研究院遼寧省食品安全重點實驗室,遼寧錦州,121013)
近年來,食物過敏現象逐年增多,已成為人類營養尤其是兒童營養中的一個重要的社會健康問題。調查表明,公眾自訴有食物過敏史的大約有15%,約8%小于3歲的兒童有過食物過敏經歷。在美國、歐洲等發達地區,已經十分重視食物過敏的研究與防治,而在我國,食物過敏問題還沒有引起足夠的重視,對食物過敏方面的研究報道較少[1-2]。
牛乳是優質的營養食品,但也是較易引起過敏反應的食物之一,約有2%~6%的嬰幼兒對牛乳蛋白有不同程度的過敏。研究表明,α-乳白蛋白(α-LA)、β-乳球蛋白和酪蛋白是牛乳中引起過敏反應的主要乳蛋白。目前,α-LA由于富含必需氨基酸、營養豐富,經常添加到嬰幼兒配方奶粉、乳飲料及運動營養補充劑中[3],但是α-LA的抗原性嚴重影響了α-LA的應用。因此,探索合適的加工方式來降低牛乳α-LA的致敏性,對于提高牛乳制品的安全性具有重要的現實意義。在蛋白質改性技術中,糖基化改性被認為是最安全、最有前景的降低牛乳蛋白致敏性的改性方法。
目前,國內對應用糖基化降低乳蛋白致敏性的研究較少,尤其是對糖的選擇方面,缺乏理論基礎。有研究表明,在蛋白質的糖基化反應中糖鏈長度對于糖基化蛋白性能的提高起著十分重要的作用。與單糖、雙糖相比,添加多糖可增加蛋白質的功能特性,包括熱穩定性、乳化性等[4]。然而,對于糖基化反應中糖鏈長度對于蛋白質抗原性的影響未有報道。因此,本研究利用α-乳白蛋白(α-LA)-糖復合物,研究糖鏈長度對α-LA-糖復合物抗原性的影響,為低敏性乳制品的開發提供一定的理論依據。
1 材料與方法
1.1 材料與儀器
胰蛋白酶,購自美國Sigma公司;甲醇(色譜級),購自美 國 Fisher 公 司;NaCl、KCl、KH2PO4、K2HPO4·12H2O、NaOH均為分析純,購自北京化學試劑公司。
電子天平(ALC-210),美國Satorious公司;酶標儀(M680),美國伯樂公司;高效液相色譜(1100),美國Agilent公司;微量振蕩器(ZW-A),江蘇省壇金市熒華儀器制造有限公司。
1.2 實驗方法
1.2.1 α-乳白蛋白-糖復合物的制備
將α-LA和不同碳鏈長度的還原寡糖(葡萄糖、麥芽糖、麥芽三糖、麥芽四糖、麥芽五糖)按摩爾比1∶13溶解于去離子水中,制成最終蛋白質質量分數為5%的混合液,用1 mol/L NaOH調節溶液pH至8.0,冷凍干燥。將凍干的混合物粉末放入培養皿中,保持相對濕度在65%(飽和KBr溶液)左右,放入培養箱中,55 ℃ 下進行糖基化反應,分別于 0、12、24、36、48 h取出混合物,所得α-LA-糖復合物溶解于去離子水中,溶液最終質量分數為5%,透析除去未反應的糖及鹽分,再次凍干,得到的凍干粉于-20℃保存。同時,將不加入糖的α-LA在相同條件下處理作為對照(ct)。
1.2.2 糖基化反應程度的測定
采用鄰苯二甲醛法(OPA法)[5]。準確稱取40 mg的OPA溶解于1 mL的甲醇中,分別加入20%(w/v)的 SDS 2.5 mL、0.1 mol/L 的硼砂 25 mL、100 μL β-巰基乙醇,最后用去離子水定容至50 mL,此為OPA試劑。測定時取50 μL的樣品(蛋白濃度2 mg/mL)與1 mL OPA溶液混合,室溫放置2 min。反應完畢后,于340 nm下測其吸光值A1,以在OPA試劑中加入50 μL去離子水為空白樣,兩者之差ΔA為自由氨基的凈吸光值。用濃度為(0.25~2.00)×10-3mol/L賴氨酸作標準曲線,通過標準曲線轉換成賴氨酸的濃度。游離氨基酸殘留量即為糖基化反應后的α-LA-糖中游離氨基酸的濃度與未反應的α-LA中游離氨基酸的濃度的比值。
1.2.3 間接競爭ELISA方法測定α-乳白蛋白及復合物的抗原性
前期通過α-LA免疫新西蘭大白兔,制備了兔抗α-乳白蛋白的多克隆抗體。經測定最高效價達到了1∶256 000;同時,所制備的抗體與乳中其他主要蛋白質不存在免疫交叉反應,特異性良好,利用此抗體測定α-乳白蛋白的抗原性,過程如下:
(1)抗原包被:用包被液(濃度為50 mmol/L,pH 9.6的碳酸鹽緩沖液)稀釋α-LA至質量濃度為0.5 μg/mL,100 μL/孔加入96 孔酶標板,于4 ℃放置過夜。
(2)樣品和抗血清預混合:在反應管中加入蛋白質量濃度為0.1 g/L的樣品和相同體積一定稀釋度(α-LA,1∶64 000)的抗血清,不加樣品的反應管作為無競爭體系,4℃放置過夜,次日傾去孔內液體,PBST(PBS+0.05%Tween-20)洗滌3次,拍干。
(3)封閉:加封閉液(含0.1 g/L明膠的包被液)進行封閉,100 μL/孔,37 ℃ 放置 1 h,PBST 洗滌 3次,拍干。
(4)抗原抗體反應:加入(2)中樣品和抗血清的預混合液,100 μL/孔,37 ℃ 溫育 1 h,PBST 洗滌 3次,拍干。
(5)加酶標二抗:用封閉液將HRP-羊抗兔IgG稀釋 5 000 倍,100 μl/孔,37 ℃溫育1 h,PBST 洗滌3次,拍干。
(6)顯色:加新鮮配制的 TMB底物溶液,100 μL/孔,37℃暗處反應15 min,顯示藍色。
(7)終止反應:加濃度為2 mol/L的H2SO4,50 μL/孔終止反應,顏色變黃。
(8)吸光值測定:利用酶標儀雙波長測定各孔的吸光值。
(9)數據處理:被測物殘留的抗原性大小用其抗原抑制率來評價,按下式計算:
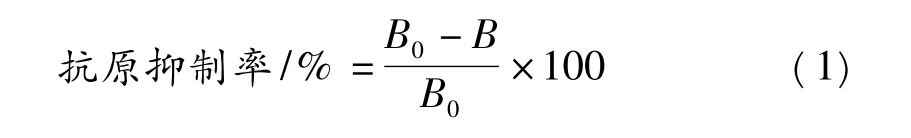
式中:B為被測樣的A值;B0為無競爭體系的A值。
1.2.4 糖基化位點分析
采用胰蛋白酶酶解糖基化后的樣品,酶與底物摩爾比1∶100,在37℃下進行并調節樣品至pH 8.5,酶解2 h后滅酶,利用高效液相色譜-質譜聯用分析糖基化蛋白酶解產物,根據蛋白一級序列分析其糖基化位點。
色譜條件:Zobalx SB-C18色譜柱(2.1 mm ×150 mm)。洗脫速度為0.2 mL/min,流動相分別為5%~45%的緩沖液A(三氟乙酸∶水=體積比0.1∶99.9),洗脫 50 min;45%~60%緩沖液 A,洗脫5 min;60%~90%的洗脫液A,洗脫65 min。質譜條件:噴霧電壓為3.5 kV,干燥氣溫度為300℃,質量掃描范圍300~2 000 m/z。
2 結果與分析
2.1 糖基化反應程度的測定
糖基化反應的基礎是蛋白質中氨基酸側鏈(主要為賴氨酸的ε-氨基)與還原糖的還原性羰基末端反應,得到共價復合物。所以,通過測定糖基化前后的自由氨基殘留量的變化可以定量分析α-LA糖基化反應程度。如圖1所示,所有糖基化的α-LA-糖復合物中的游離氨基酸殘留量均隨反應時間的增加而降低,表明α-LA與還原糖之間發生了交聯。其中,單糖與α-LA混合物游離氨基酸降低最快,在處理48 h后,葡萄糖-α-LA的游離氨基酸殘留量為40.59%。在與其他糖的反應中,游離氨基酸降低較慢,麥芽糖-α-LA、麥芽三糖-α-LA、麥芽四糖-α-LA 和麥芽五糖-α-LA的游離氨基酸殘留量分別為50.49%、57.98%、70.38%和72.94%。結果表明,與α-LA的糖基化反應中,糖的反應性由高到低分別為:葡萄糖>麥芽糖>麥芽三糖>麥芽四糖>麥芽五糖。糖基化程度以及反應速率隨糖鏈長度的增加而降低,可能是由不同分子大小的糖所引起的結合位阻不同而引起的[6]。糖鏈越短,所含的開鏈形式越多,糖與蛋白質中氨基的反應活性就越強[7-8]。
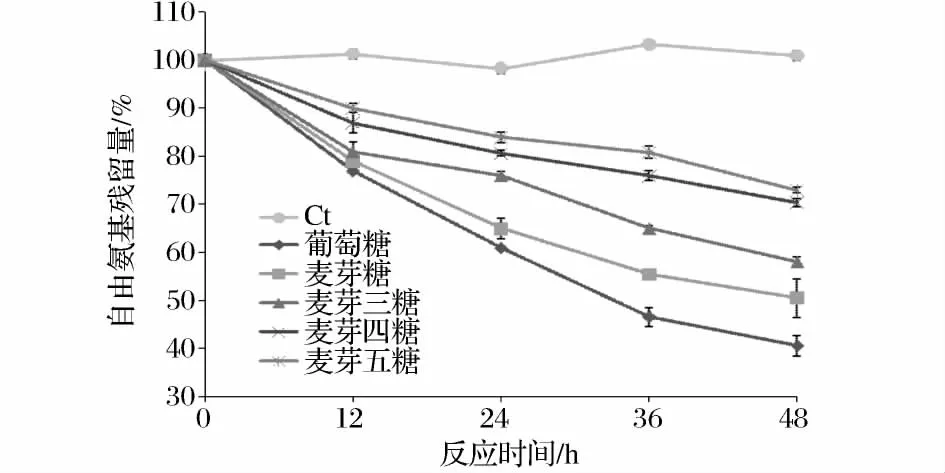
圖1 糖鏈長度對自由氨基殘留量的影響Fig.1 Effect of saccharide size on free amino groups of glycated α-LA
2.2 α-乳白蛋白-糖復合物抗原性分析
圖2顯示的是不同鏈長寡糖糖基化產物抗原性的變化。在55℃加熱條件下,未經過糖基化的α-LA的抗原性伴隨加熱時間的延長稍有提高,這可能是由于長時間的熱處理,使α-LA的構象改變,蛋白質空間結構的展開導致分子內部的部分抗原決定簇暴露[9],這與 Kleber[10]等人研究 β-LG 加熱條件下抗原性變化的趨勢一致。
5種糖基化樣品的抗原性與原α-LA蛋白相比,抗原性均有明顯下降。α-LA的抗原性隨糖基化反應時間的增長而降低,說明糖基化程度對于降低蛋白質抗原性起著十分重要的作用。被引入的糖分子通過空間位阻和電荷作用,掩蔽蛋白上的抗原決定簇,從而降低α-LA的抗原性[11]。然而,抗原性降低效果最好的是麥芽三糖-α-LA復合物,48h糖基化處理后,α-LA的抗原抑制率達到51.67%,但是由糖基化反應程度結果看,麥芽三糖-α-LA的糖基化程度并不是最高的。這說明,糖與蛋白的結合部位可能對其抗原性起到更為關鍵的作用。
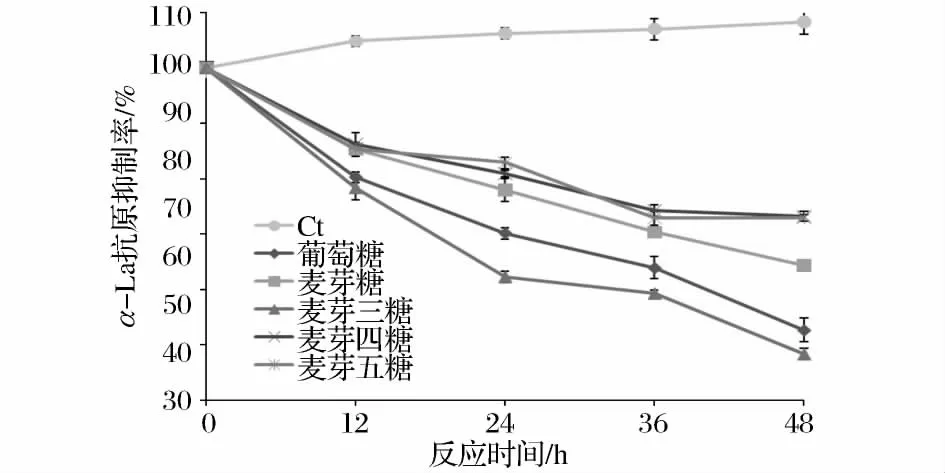
圖2 糖鏈長度對α-乳白蛋白抗原性的影響Fig.2 Effect of saccharide size on the antigenicity of α-LA
2.3 糖基化位點分析
圖3為葡萄糖-α-LA糖基化樣品的酶切肽段的ESI質譜圖,其中,經過分子質量對比分析(有1分子葡萄糖加入到肽段上,肽段分子質量增加162)得出糖基化肽段有6條,分別為肽段(17~62)、(59~79)、(80~94)、(95~108)、(99~114)和(109~122)。結果表明,葡萄糖糖基化樣品中,Lys58、Lys62、Lys93、Lys98、Lys108以及 Lys1146個賴氨酸殘基被糖基化。利用同樣的方法,分析麥芽糖、麥芽三糖、麥芽四糖及麥芽五糖糖基化產物的糖基化位點,如表1所示。對比5種還原糖的糖基化位點及其糖基化產物的抗原性,我們發現Lys58糖基化位點可能對于α-LA的抗原性起重要作用。Lys58位點僅在抗原性降低最多的葡萄糖-α-LA和麥芽三糖-α-LA中被糖基化。由于糖基化作用是通過被引入的糖分子的空間位阻和電荷作用,掩蔽蛋白上的抗原決定簇,從而降低蛋白的過敏原性。因此,我們判斷Lys58位點位于α-LA的抗原決定簇序列上。Maynard[12]等人的研究也指出,肽段(17~58)以及包含此段氨基酸序列的更大肽段與IgE有強烈的反應,表明肽段(17~58)為α-LA的主要抗原決定簇。綜合以上結果說明糖基化反應產物的抗原性受糖基化程度及糖基化位置的共同影響。
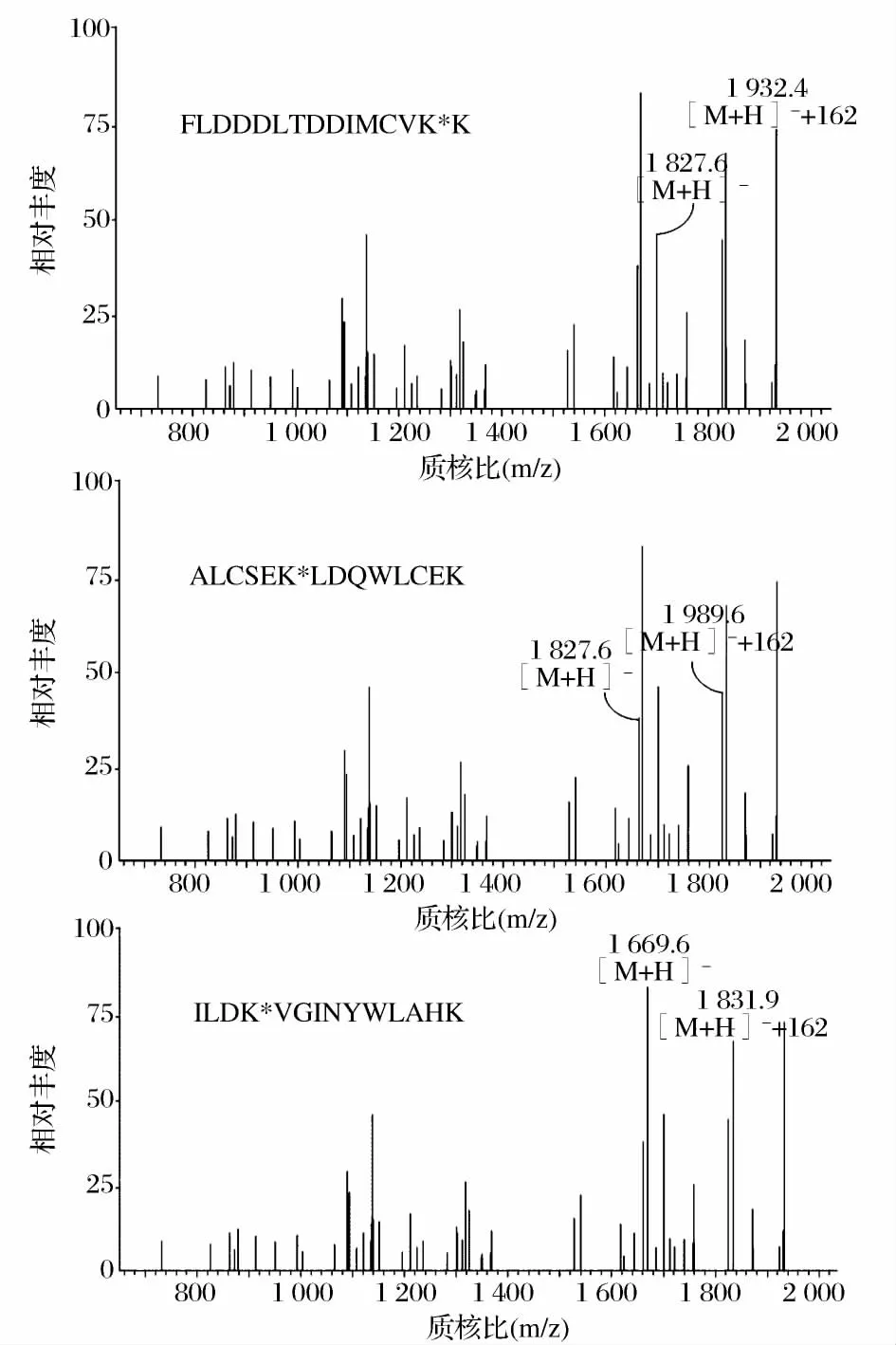
圖3 葡萄糖-α-LA的糖基化肽段(80~94,95~108,109~122)Figure 3 Sequences correspond to glycated peptides of
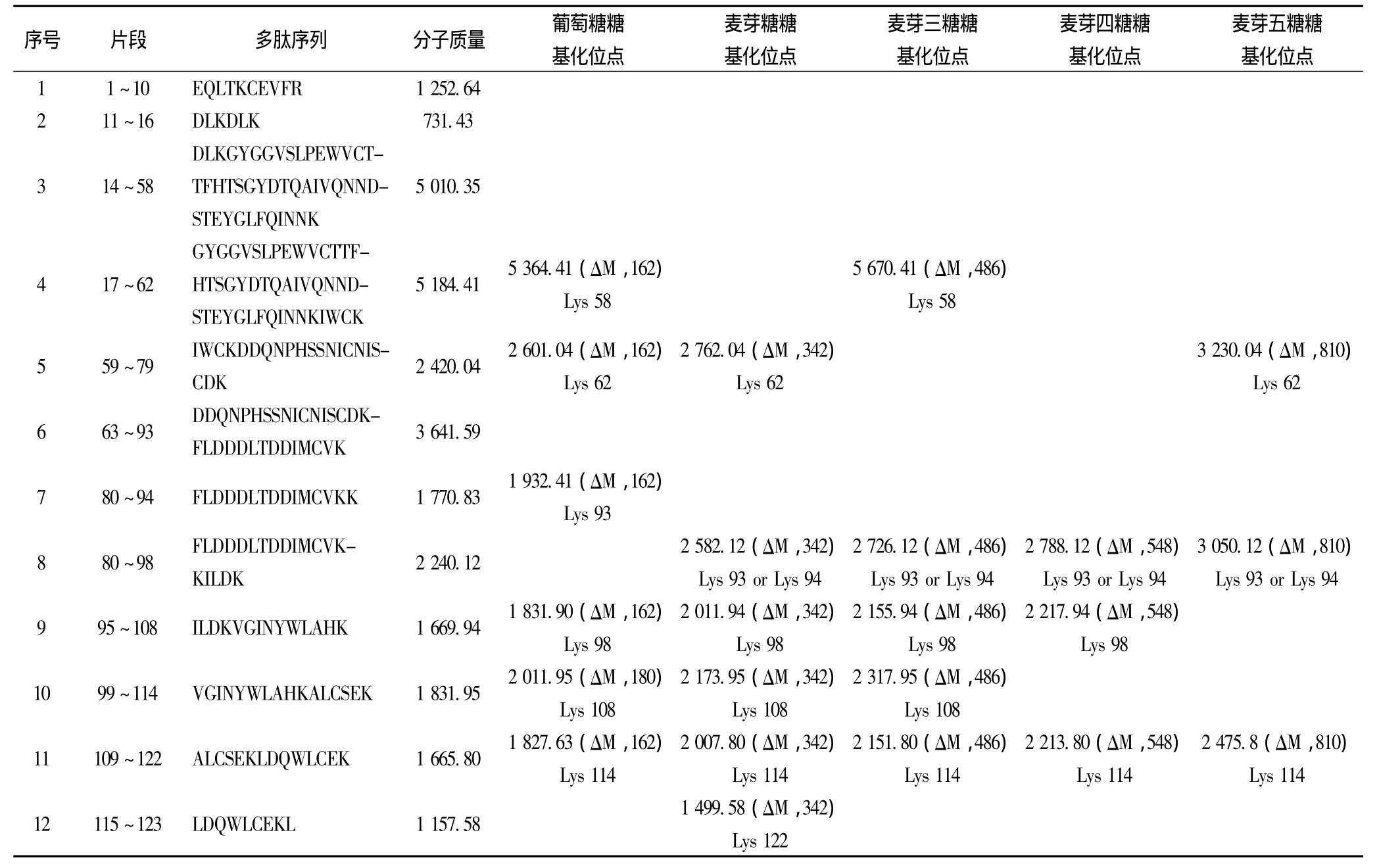
表1 α-乳白蛋白糖基化樣品胰蛋白酶酶解肽段質譜分析Table 1 HPLC-MS/MS analysis of peptides obtained from the trypsin digestion of glycated α-LA after glycation
3 結論
本文重點研究了5種不同糖鏈長度的糖與α-乳白蛋白糖基化反應對后者抗原性的影響,結果表明糖基化程度隨糖鏈長度的增長而降低。此外,經過糖基化反應α-LA抗原性顯著下降,并且α-LA的抗原性隨糖基化反應時間的增長而降低,說明糖基化程度對于降低蛋白質抗原性起著十分重要的作用。但是抗原性降低幅度最大的是麥芽三糖-α-LA,進一步通過測定糖基化位點,發現Lys58位點僅存在于抗原性降低最多的葡萄糖-α-LA和麥芽三糖-α-LA中,說明α-LA糖基化產物的抗原性受糖基化程度及糖基化位點的共同影響。
[1]包怡紅.乳清多肽及其發酵制品的研究[D].哈爾濱:東北農業大學,2001:11-34.
[2]Maldonado J,Gil A,Narbona E,et al.Special formulas in infant nutrition:a review[J].Early Human Development,1998,53(1):S23-S32.
[3]L?nnerdal B,Lien E L.Nutritional and physiologic significance of α-lactalbumin in infants[J].Nutrition Reviews,2003,61(9):295-305.
[4]Shu Y W,Sahara S,Nakamura S,et al.Effects of the length of polysaccharide chains on the functional properties of the Maillard-type lysozyme-polysaccharide conjugate[J].Journal of Agricultural and Food Chemistry,1996,44(9):2 544-2 548.
[5]Fayle S E,Healy J P,Brown P A,et al.Novel approaches to the analysis of the Maillard reaction of proteins[J].E-lectrophoresis,2001,22(8):1 518-1 525.
[6]Nacka F,Chobert J M,Burova T,et al.Induction of new physicochemical and functional properties by the glycosylation of whey proteins[J].Journal of Protein Chemistry,1998,17(5):495-503.
[7]Chevalier F,Chobert J M,Popineau Y,et al.Improvement of functional properties of β-lactoglobulin glycated through the Maillard reaction is related to the nature of the sugar[J].International Dairy Journal,2001,11(3):145-152.
[8]Pokharna H K,Pottenger L A.Nonenzymatic glycation of cartilage proteoglycans:an in vivo and in vitro study[J].Glycoconjugate Journal,1997,14(8):917-923.
[9]Kleber N,Hinrichs J.Antigenic response of β-lactoglobulin in thermally treated bovine skim milk and sweet whey[J].Milchwissenschaft,2007,62(2):121-124.
[10]Kleber N,Krause I,Illgner S,et al.The antigenic response of β-lactoglobulin is modulated by thermally induced aggregation[J].European Food Research and Technology,2004,219(2):105-110.
[11]Arita K,Babiker E E,Azakami H,et al.Effect of chemical and genetic attachment of polysaccharides to proteins on the production of IgG and IgE[J].Journal of Agricultural and Food Chemistry,2001,49(4):2 030-2 036.
[12]Maynard F,Jost R,Wal J M.Human IgE binding capacity of tryptic peptides from bovine α-lactalbumin[J].International Archives of Allergy and Immunology,1997,113(4):478-488.
