質粒介導的氟喹諾酮類抗生素耐藥機制研究進展
2014-11-23 03:55:44孔令聰劉樹明馬紅霞
中國獸藥雜志 2014年7期
王 梓,孔令聰,高 鐸,劉樹明,馬紅霞,2*
(1.吉林農業大學動物科技學院,長春130118;2.吉林農業大學動物生產及產品質量安全教育部重點實驗室,長春130118)
氟喹諾酮類藥物作為廣譜抗菌藥自1962年投入臨床使用至今已有50余年歷史,已成為臨床中最為常用的抗生素之一。氟喹諾酮類藥物屬于濃度依賴型抗生素,致病菌如果長期處于其亞抑菌濃度時極易產生突變成為耐藥菌[1-3]。作為氟喹諾酮類藥物的主要耐藥機制之一,質粒介導的氟喹諾酮類藥物耐藥(Plasmid-mediated quinolone resistance,PMQR)具有傳播速度快、基因亞型較多等特點,且其攜帶的耐藥基因可對細菌染色體上氟喹諾酮類藥物耐藥基因進行靶位修飾,使靶位突變頻率增加[4-5],嚴重影響氟喹諾酮類藥物的使用壽命。故探索質粒介導的氟喹諾酮類耐藥機制對于控制耐藥基因的快速傳播,減少耐藥菌的危害具有重要意義。
1 質粒介導的氟喹諾酮類藥物耐藥基因的發現
現有研究表明,質粒攜帶的氟喹諾酮類藥物耐藥基因主要有3種,分別為包含有多種亞型的qnr基因、介導主動外排的基因、發生點突變的氨基糖苷乙酰轉移酶基因。其中,Martínez-Martínez等在研究肺炎克雷伯菌天然攜帶的pMG252質粒時,首次發現了五肽重復序列qnr基因[6],隨后陸續報道了多種qnr基因亞型[7-9]。此后,在對 qnr基因的作用機制進行研究時發現,qnr基因既能介導氟喹諾酮類藥物耐藥,還可以通過增加gyrA、parC等靶位基因的突變頻率,使細菌耐藥能力增強[10]。各qnr基因亞型概況如表1所示。隨后,Haru Kato等[11]在日本確證了由質粒介導的外排基因QepA,在該試驗中發現,QepA基因能夠介導細菌對諾氟沙星的外排作用,且其外排可以被外排泵抑制劑CCCP(carbony cyanide m-chlorophenyl-hydrazone)抑制,為質子依賴型外排泵。由此推測,QepA基因所介導的外排作用可能是質粒介導的氟喹諾酮類藥物耐藥的重要途徑之一。G.A.Jacoby等在研究中又發現了另一種PMQR基因aac(6')-Ib-cr,該基因由氨基糖苷乙酰轉移酶基因發生Trp102Arg和Asp179Tyr點突變獲得,其可介導大腸桿菌對環丙沙星的抗性[12],該基因自發現以來在氟喹諾酮耐藥菌株中傳播迅速,在三類PMQR基因中檢出率較高[13-15]。
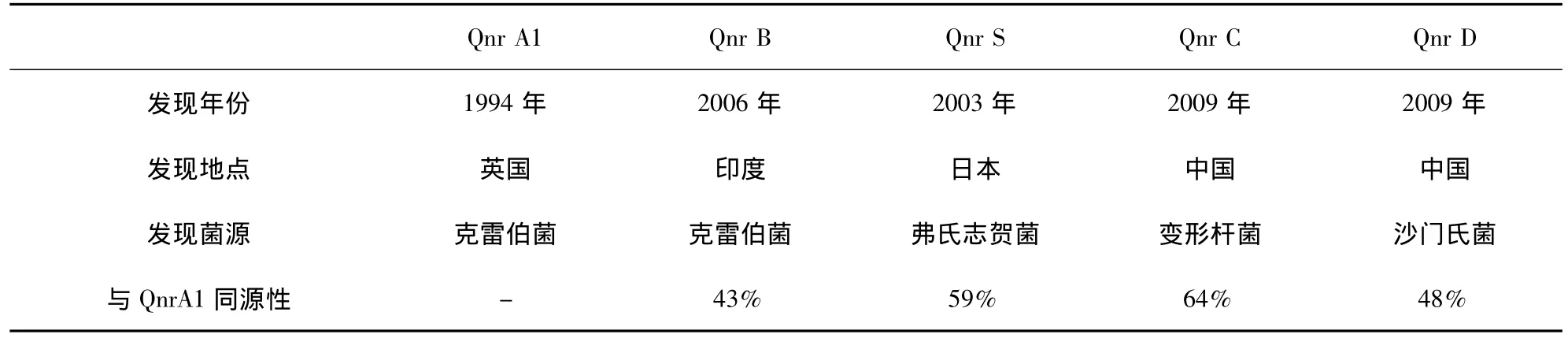
表1 qnr基因各亞型概況
2 質粒介導的氟喹諾酮類藥物耐藥基因的分布
現有流行病學調查結果發現,多種PMQR基因常共存于同一耐藥質粒中。Kees Veldman等在歐洲13個國家分離出的沙門氏菌和大腸桿菌中分別檢測出 qnrA1、qnrB、qnrD、aac(6’)-Ib-cr基因,其中 qnrD基因為在歐洲出現的首次報道[16];Ya-Hong Liu等在廣東省分離的大腸桿菌中分別檢測出 qnrB、qnrS、aac(6’)- Ib-cr、QepA 等基因[17];Patricia Andres等在南美洲分離的百余株腸桿菌中也擴增出 qnrB、qnrS、aac(6’)-Ib-cr等氟喹諾酮耐藥基因[18]。由于單獨的PMQR基因通常只介導低水平耐藥,故耐藥菌常通過多種耐藥基因的共同作用以形成高水平耐藥性。近年來,由于動物運輸、耐藥質粒的轉移等因素,PMQR基因已經在世界范圍內廣泛分布,伴隨著PMQR基因突變的不斷發生,未來將會有更多的PMQR基因耐藥亞型不斷出現。同時,攜帶氟喹諾酮類藥物耐藥基因的質粒也可能攜帶其他多種耐藥基因,報道最多的為與廣譜β-內酰胺酶基因共存。通過多種類耐藥基因在同一質粒上的整合即可形成多重耐藥質粒,而且多重耐藥質粒的分布范圍也在逐步擴大,在沙門氏菌、克雷伯菌、變形桿菌中均發現多重耐藥質粒的存在[19-20],但質粒攜帶的多種耐藥基因的表達水平是否可以相互影響,還需進一步研究驗證。
3 質粒介導的氟喹諾酮類藥物耐藥機制
3.1 PMQR基因的傳播機制 攜帶PMQR基因的耐藥質粒快速傳播是目前氟喹諾酮類藥物耐藥性增強的重要因素之一。質粒上攜帶的PMQR基因可以直接通過質粒轉移實現基因的相互傳遞,也可利用質粒中整合子、轉座子的傳遞實現PMQR基因的快速傳播。Humberto Barrios等將攜帶β-內酰胺耐藥基因(CTX-M-15)和多個氟喹諾酮耐藥基因(qnrA1、qnrB2、aac(6’)-Ib-cr)的質粒在不同溫度下進行接合轉移實驗:結果在受體菌中發現了兩個不同的質粒,其中大小為300 kb的質粒pSZ300中攜帶了 CTX -M -15、qnrA1、aac(6’)-Ib-cr基因,并產生氟喹諾酮類藥物抗性;而大小為50 kb的質粒pSZ50中僅發現有qnrA1基因,測序結果顯示 pSZ50包含于質粒 pSZ300中[21]。此試驗證明,質粒在水平傳播過程中能發生重組,從而導致耐藥質粒多樣化。Be'la Kocsis等首次發現在肺炎克雷伯菌的質粒ST147中qnrS1可與窄譜β-內酰胺酶LAP-2基因共存,并進一步通過質粒接合轉移實驗獲得了一個97 kb質粒,其同時攜帶有供體菌上的上述兩種耐藥基因,進而使受體菌對頭孢他啶、環丙沙星等藥物的MIC值均提高了2倍以上[22]。如果細菌的染色體上存在耐藥基因,則質粒介導的PMQR機制可促使細菌產生更高水平的耐藥,給臨床治療帶來困難。通過類似多藥耐藥轉移質粒的橫向傳播,將會使不同菌屬細菌間形成復雜的高水平多重耐藥機制。而設法限制耐藥質粒的快速傳播,將成為減緩細菌對氟喹諾酮類抗菌藥的主要途徑之一。
3.2 PMQR基因表達的調控機制 目前有關QepA、aac(6’)-Ib-cr基因的研究多集中于流行病學調查中耐藥基因的種類檢測,關于影響PMQR基因調節和表達的報道還較少。最近的研究證實,細菌的SOS反應是調節qnr基因表達量的重要因素之一。SOS反應是細菌在DNA受到持續損傷壓力下產生的一種自救反應,可以由LexA、RecA等基因調控,能夠增加細菌耐藥基因的突變頻率。近年來已有學者相繼報道了多種qnr基因可以在細菌發生SOS反應時發生表達量的改變。其中,David C.Hooper等通過改變溫度增加細菌的生存壓力使其產生SOS反應的實驗結果發現,受試菌的qnrB基因表達量增加,進而對環丙沙星的耐受能力明顯增強,對qnrB基因進一步基因環境研究表明,qnrB基因的上游序列中存在的LexA基因可對細菌DNA的持續損傷壓力產生應答,從而引發細菌的SOS反應提高qnrB的表達量[23]。隨后,A.Briales等發現qnrD基因以及一個新型染色體攜帶的Smaqnr基因都擁有相似的LexA結構,可以在不同濃度的環丙沙星、莫西沙星等藥物作用下將上述qnr基因表達量提高 1.5 ~16 倍[23-24]。上述實驗結果證明,通過溫度改變、藥物作用等方式增加細菌生存壓力時,可以選擇性的誘發細菌的SOS反應,繼而使含有LexA結構的qnr基因發生表達量的改變,使細菌產生更強的耐藥性。目前,有關三種PMQR基因表達機制的深入探索還有待進一步的研究。
3.3 PMQR基因的作用機制 在三種PMQR基因中,QepA的作用機制主要通過主動外排系統減少藥物在細菌菌體內的聚集,其外排泵結構與MFS外排泵超家族極為相似,為質子依賴型外排泵,細菌可依靠其外排作用提高對諾氟沙星、環丙沙星等藥物的MIC值2倍以上,并能在其他耐藥基因共同作用下使細菌產生更強耐藥性[11]。aac(6’)-Ib-cr的作用機制為,當 aac(6’)-Ib基因的102位色氨酸突變為組氨酸及179位天冬氨酸突變為酪氨酸共同發生時,即可將環丙沙星與細菌結合位點乙酰化,從而使其失去對細菌的抑制作用。細菌存在aac(6’)-Ib-cr基因的同時,還可增加染色體上相關耐藥基因的突變頻率,從而增強細菌的耐藥性。而當QepA基因與aac(6’)-Ib-cr基因共存與耐藥質粒上時,可使細菌對環丙沙星的耐藥性較qnrA基因單獨作用時提高4倍以上,達到藥物敏感性折點[25]。而目前有關qnr基因作用機制尚未完全闡明,其機制的探索主要參考其他五肽重復蛋白(pentapeptide repeat-containing proteins),如染色體上同樣可以介導氟喹諾酮類藥物耐藥的McbG和MfpA蛋白。編碼McbG和MfpA的基因與qnr基因具有19.6%和18.9%的同源性,其主要耐藥機制是通過保護DNA促旋酶和抑制藥物對DNA的超螺旋作用[26-27]。近年來新發現了兩種新型染色體攜帶的 qnr-like基因,即 EfsQnr和 Smaqnr。其作用機制與MfpA較相似,可以抑制ATP依賴型DNA超螺旋,從而產生對氟喹諾酮類藥物的耐藥性;而Smaqnr基因與qnrB1基因具有高達80%的同源性,是迄今為止發現的同源性最高的qnr-like基因[28-29]。通過對這些染色體上攜帶的qnr-like基因作用機制的深入研究,可以逐步探索質粒攜帶qnr基因的作用機制及遺傳環境,為質粒介導的氟喹諾酮類藥物耐藥機制的進一步闡明及在根本上限制細菌的耐藥性奠定基礎。
4 展望
隨著質粒介導的氟喹諾酮類藥物耐藥機制研究的不斷深入,人們對細菌氟喹諾酮類耐藥性的產生及傳播有了更全面的認識。但同時,細菌的耐藥性也日趨復雜,近期研究顯示,質粒介導的 aac(6’)-Ib-cr基因在染色體中也被發現[25],這說明細菌正通過改變自身的耐藥途徑來增加人們控制其耐藥性的難度。在未來的研究中,可能還會有更多耐藥基因在染色體和質粒中相互整合,形成復雜的耐藥環境。因此,在規范臨床合理用藥的同時,利用現有細菌耐藥機制研究的深層成果,通過基因敲除、反義RNA等生物學方法控制耐藥質粒的快速傳播、抑制耐藥基因的表達,將是未來控制細菌耐藥性的有效途徑之一。同時,對于細菌耐藥機制的研究也應繼續進行,闡明耐藥基因在染色體與質粒中的整合途徑及影響方式等未知耐藥機制。相信經過廣大科研工作者的不斷努力,一定能夠有效控制細菌耐藥性,為人類的生命安全提供保障。
[1]Veldman K,Cavaco L M,Mevius D,et al.International collaborative study on the occurrence of plasmid-mediated quinolone resistance in Salmonella enterica and Escherichia coli isolated from animals,humans,food and the environment in 13 European countries[J].Journal of antimicrobial chemotherapy,2011,66(6):1278-1286.
[2]王小亮,徐立蒲,曹 歡,等.魚源病原菌對氟氟喹諾酮類藥物的耐藥性分析[J].中國畜牧獸醫,2013,40(3):195-198.
[3]Nozomi Aoike,Tomoo Saga,Sakata R,et al.Molecular Characterization of Extraintestinal Escherichia coli Isolates in Japan:Relationship between Sequence Types and Mutation Patterns of Quinolone Resistance-Determining Regions Analyzed by Pyrosequencing[J].Journal of Clinical Microbiology,2013,51(6):1692.
[4]Xiang Chen,Weiqiu Zhang,Pan W,et al.Prevalence of qnr,aac(6’)-Ib-cr,qepA,and oqxAB in Escherichia coli Isolates from Humans,Animals,and the Environment[J].Antimicrob Agents Chemother,2012,56(6):3423 -3427.
[5]Monika Dolejska,Laura Villa,Henrik Hasman et al.Characterization of IncN plasmids carrying blaCTX-M-1 and qnr genes in Escherichia coli and Salmonella from animals,the environment and humans[J].J Antimicrob Chemother,2013,68:333 -339.
[6]Martínez- Martínez L,Pascual A,Jacoby G A.Quinolone resistance from a transferable plasmid[J].The Lancet,1998,351(9105):797-799.
[7]Hata M,Suzuki M,Matsumoto M,et al.Cloning of a novel gene for quinolone resistance from a transferable plasmid in Shigella flexneri 2b[J].Antimicrobial agents and chemotherapy,2005,49(2):801-803.
[8]Jacoby G A,Walsh K E,Mills D M,et al.qnrB,another plasmid-mediated gene for quinolone resistance[J].Antimicrob Agents Chemother,2006,50:1178 -1182.
[9]Wang M,Q Guo,Xu X,et al.New plasmid-mediated quinolone resistance gene,qnrC1,found in a clinical isolate of Proteus mirabilis[J].Antimicrob.Agents Chemother,2009,53:1892 -1897.
[10]Cattoir V,P Nordmann.Plasmid-mediated quinolone resistance in gram-negative bacterial species:an update[J].Curr Med Chem,2009,16:1028 -1046.
[11]Yamane K,Wachino J,Suzuki S,et al.New plasmid-mediated fluoroquinolone efflux pump,QepA,found in an Escherichia coli clinical isolate[J].Antimicrobial agents and chemotherapy,2007,51(9):3354-3360.
[12]Robicsek A,Strahilevitz J,Jacoby G A,et al.Fluoroquinolone-modifying enzyme:a new adaptation of a common aminoglycoside acetyltransferase[J].Nat Med,2006,12:83 -88.
[13]Virginia Garcia- Fulgueiras,Luciana Robino,Gabriela Algorta,et al.Extended-spectrum β -lactamases and plasmid-mediated quinolone resistance in enterobacterial clinical isolates in the paediatric hospital of Uruguay[J].Antimicrob Chemother,2011,66:1725-1729.
[14]Fortini D,Fashae K,García-Fernández A,et al.Plasmid -mediated quinolone resistance and β-lactamases in Escherichia coli from healthy animals from Nigeria[J].Journal of antimicrobial chemotherapy,2011,66(6):1269-1272.
[15]Walsh F,Rogers T R.Comparison of plasmid-mediated quinolone resistance and extended-spectrum β-lactamases in third-generation cephalosporin-resistant Enterobacteriaceae from four Irish hospitals[J].Journal of medical microbiology,2012,61(1):142-147.
[16]Veldman K,Cavaco L M,Mevius D,et al.International collaborative study on the occurrence of plasmid-mediated quinolone resistance in Salmonella enterica and Escherichia coli isolated from animals,humans,food and the environment in 13 European countries[J].Journal of antimicrobial chemotherapy,2011,66(6):1278-1286.
[17]Liu B T,Liao X P,Yang S S,et al.Detection of mutations in the gyrA and parC genes in Escherichia coli isolates carrying plasmid-mediated quinolone resistance genes from diseased food-producing animals[J].Journal of medical microbiology,2012,61(Pt 11):1591-1599.
[18]Andres P,Lucero C,Soler-Bistué A,et al.Differential distribution of plasmid-mediated quinolone resistance genes in clinical enterobacteria with unusual phenotypes of quinolone susceptibility from Argentina[J].Antimicrobial agents and chemotherapy,2013,57(6):2467 -2475.
[19]Hu Y,Cai J,Zhang R,et al.Emergence of Proteus mirabilis Harboring blaKPC-2 and qnrD in a Chinese Hospital[J].Antimicrobial agentsand chemotherapy, 2012, 56(5):2278-2282.
[20]Herrera-León S,González- Sanz R,Herrera-León L,et al.Characterization of multidrug-resistant Enterobacteriaceae carrying plasmid-mediated quinolone resistance mechanisms in Spain[J].Journal of Antimicrobial Chemotherapy,2011,66(2):287-290.
[21]Ulises Garza- Ramos,Humberto Barrios,Maria Jose Hermandez- Vargas,et al.Transfer of quinolone resistance gene qnrA1 to Escherichia coli through a 50 kb conjugative plasmid resulting from the splitting of a 300 kb plasmid[J].Antimicrob Chemother,2012;67:1627-1634.
[22]Kocsis B,Kocsis E,Fontana R,et al.Identification of blaLAP-2 and qnrS1 genes in the internationally successful Klebsiella pneumoniae ST147 clone[J].Journal of medical microbiology,2013,62(Pt 2):269-273.
[23]Wang M,Jacoby G A,Mills D M,et al.SOS regulation of qnrB expression[J].Antimicrobial agents and chemotherapy,2009,53(2):821-823.
[24]Briales A,Rodriguez-Martinez J M,Velasco C,et al.Exposure to diverse antimicrobials induces the expression of qnrB1,qnrD and smaqnr genes by SOS-dependent regulation[J].Journal of antimicrobial chemotherapy,2012,67(12):2854-2859.
[25]Elena Ruiz,Yolanda Saenz,Myriam Zarazaga,et al.Qnr,aac(6’)-Ib- cr and qepA genes in Escherichia coli and Klebsiella spp.:genetic environments and plasmid and chromosomal location[J].Journal of antimicrobial chemotherapy,2012,67:886 -897.
[26]Rodríguez- Martínez J M,Velasco C,Briales A,et al.Qnrlike pentapeptide repeat proteins in gram-positive bacteria[J].Journal of antimicrobial chemotherapy,2008,61(6):1240-1243.
[27]Jacoby G A,Hooper D C.Phylogenetic analysis of chromosomally determined Qnr and related proteins[J].Antimicrobial agents and chemotherapy,2013,57(4):1930-1934.
[28]Hegde S S,Vetting M W,Mitchenall L A,et al.Structural and biochemical analysis of the pentapeptide repeat protein EfsQnr,a potent DNA gyrase inhibitor[J].Antimicrobial agents and chemotherapy,2011,55(1):110 -117.
[29]Velasco C,Rodriguez-Martinez J M,Briales A,et al.Smaqnr,a new chromosome-encoded quinolone resistance determinant in Serratia marcescens[J].Journal of antimicrobial chemotherapy,2010,65(2):239-242.
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
四川勞動保障(2021年9期)2022-01-18 05:11:08
科學大眾(2020年12期)2020-08-13 03:22:22
文苑(2018年21期)2018-11-09 01:23:06
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
中國當代醫藥(2015年17期)2015-03-01 02:03:58
中國衛生(2014年3期)2014-11-12 13:18:12
