乳制品乳酸菌遺傳學概述*
2014-12-16 08:01:54焦晶凱劉振民莫蓓紅
食品與發酵工業 2014年1期
焦晶凱,劉振民,莫蓓紅
(乳業生物技術國家重點實驗室,光明乳業股份有限公司乳業研究院,上海,200436)
乳酸菌作為人和動物腸道的正常菌群,具有阻止病原菌對腸道的感染和侵害、抑制內毒素的產生、維持腸道的微生態平衡、提高機體免疫力、預防和抑制腫瘤的發生、降低膽固醇、延緩衰老和抗輻射等功效。乳酸菌還有抑制過敏發炎反應,對氣喘、異位性皮膚炎有很好的效果。特別是口服乳酸菌具有免疫耐受性作用,微量釋放過敏原疫苗蛋白,降低過敏炎性反應,改善氣喘和減少過敏性鼻炎發作的機會等[1]。
新的分子生物學技術極大地促進了在宿主微生物區系水平上對乳酸菌益生作用的研究,乳酸菌遺傳學的主要突破是基因組學研究的進步,目前已成功獲得了幾個種類乳酸菌的全基因組序列[2]。全基因組序列無疑會提高我們對乳酸菌代謝、乳酸菌間水平基因轉移及其生態作用的認識和了解。特別是在乳糖和檸檬酸代謝、蛋白質水解[3]、噬菌體抗性機制及細菌素產生等方面,通過全基因組序列可以了解這些過程是如何被控制或調節的。詳細的序列信息可以在轉錄和翻譯水平上開辟基因表達分析的途徑,從而進一步推斷這些微生物在益生方面的代謝模式。隨著人們對分子生物學的進一步了解以及生物信息學研究的深入,在宿主基因表達水平上研究益生菌與宿主相互作用亦成為可能。
1 基因組成
細菌中的基因組成分為兩個部分,大部分基因作為主要DNA分子存在于細菌染色體中,另一小部分儲存于自主復制的質粒中。乳酸菌遺傳學研究的主要是它們當中的質粒和噬菌體,這些質粒和噬菌體在乳酸菌中起著比較重要的作用,對比染色體,采用分子遺傳分析方法來分析的質粒和噬菌體更加容易獲取和研究。眾所周知,好的發酵劑菌株對于乳制品發酵來說至關重要,但是這些菌株的某些特性并不穩定,而大部分原因歸咎于質粒的缺失,質粒的編碼消除等對菌株的特性至關重要。
1.1 質粒DNA的檢測及其豐度
1972年,Mckay等人[4]推斷乳酸鏈球菌(Str.lactis)喪失利用乳糖的能力可能是由于質粒消除引起,此后乳酸鏈球菌染色體外DNA研究開始映入人們的視線。對質粒研究之初,是通過顯微鏡對放射性標記的質粒DNA進行檢測的。對這種質粒DNA進行電子顯微鏡檢查發現,這些不同大小的環狀質粒分子可以由其拉伸長度進行評定。隨后研究者發現,瓊脂糖電泳可以更加快速而有效的進行乳酸菌質粒DNA的檢測。隨著技術研究的進步,對質粒DNA檢測技術更加完善,分子質量超過30兆道爾頓的質粒DNA可以很容易的被檢測出來。目前已發展了更多快速篩選菌株中質粒含量的方法[5]。
質粒譜提供了一個有效的“指紋圖譜”,可以更簡便地去觀察菌株特性及其相互之間的關系。通過質粒譜觀察到乳酸鏈球菌(Str.lactis)菌株和乳脂鏈球菌(Str.cremoris)菌株中包含大而復雜的質粒,相反,菌株嗜熱鏈球菌(Str.thermophilus)不包含質粒或包含質粒數目較少。不同乳桿菌中所包含的質粒數目差異較大,通常乳桿菌中不包含或僅僅包含一個質粒,但如嗜酸乳桿菌(Lb.acidophilus)等菌株包含多個質粒。片球菌(Pediococci)包含0~2個質粒,Leuconostoc包含2 個質粒[6]。
1.2 質粒消除法和表型歸屬性
研究發現,質粒的突然消失能夠對表型帶來影響,吖叮黃、溴化乙錠濃度和溫度的增加對消除頻率會產生影響,由此通過觀察菌株間基因轉移的特征得出表型是由質粒基因控制的結論。找到決定菌株表型的質粒中特定分子是目前的一大研究熱點。質粒消除法對研究特定質粒的特性尤為重要,例如乳酸鏈球菌712(Str.lactis712)和雙醋酸乳鏈球菌176(Str.Lactis diacetylactis 176)中的乳糖質粒,研究者們觀察了在原生質體恢復之后乳酸鏈球菌712(Str.lactis 712)質粒的消除現象,并通過連續的對質粒進行消除獲得了僅攜帶特定質粒分子的菌株及非攜帶質粒的菌株。通過觀察這些“衍生”的菌株的表型,可以清晰地辨別出乳糖質粒分子,這是通過傳統的表型消除法和質粒圖譜分析難以實現的。乳酸鏈球菌復雜的質粒組成為分析質粒編碼的基因轉移引起的表型的改變帶來很大困難,并且使對純化質粒進行分子分析更加復雜化,基于以上原因,分離出帶單一質粒的菌株及非攜帶質粒的菌株更具有重要意義,目前,已經成功使用N-甲基-N’-硝基-N-亞硝基胍試劑分離出非攜帶質粒的乳酸鏈球菌712(Str.lactis 712)和乳酸鏈球菌C2(Str.lactis C2)、通過使用”原生質體消除”法獲得了非攜帶質粒的乳酸鏈球菌TL1987(Str.lactis TL1987)、通過吖叮黃獲得了非攜帶質粒的雙醋酸乳鏈球菌176(Str.lactis subsp.diacetylactis 176)及通過提高溫度的方法獲得了非攜帶質粒的雙醋酸乳鏈球菌BU2(Str.lactis subsp.diacetylactis BU2)和乳脂鏈球菌1200(Str.cremoris 1200)。
1.3 乳糖和蛋白酶質粒
表型、遺傳、物理等多種方法已被用來證明在乳鏈球菌中乳糖的發酵能力是由質粒編碼調控的。在20世紀30年代有學者就研究了乳糖發酵能力的突然消失的原因,1972年McKay等人研究得出增加吖叮黃的濃度或增加溫度可引起乳糖質粒消除的增加[4]。研究已證明在乳酸鏈球菌(Str.lactis)和雙醋酸乳酸鏈球菌(Str.Lactis subsp.diacetylactis)中乳糖基因是由特殊的分子控制的。由消除實驗鑒定了在菌株乳脂鏈球菌B1(Str.cremoris B1)中含有36Mdal乳糖質粒,但在該屬的其他菌株中,并未得到相關的乳糖質粒[7]。隨后,有學者研究當一些乳脂鏈球菌(Str.cremoris)菌株作為供體與乳酸鏈球菌(Str.lactis)進行基因轉移時,乳糖基因轉移和質粒分子的轉移表現出了一定的相關性,Snook和McKay[8]推斷乳脂鏈球菌(Str.cremoris)菌株可能攜帶了罕見穩定的乳糖質粒。而有研究者發現一個非攜帶質粒的乳脂鏈球菌菌株1200(Str.cremoris 1200)依然保留了乳糖發酵能力,因此,在不同乳脂鏈球菌(Str.cremoris)菌株中乳糖表型差異很大,一些攜帶了乳糖質粒,一些僅包含染色體乳糖基因,此外在一些乳桿菌中也鑒定出了乳糖質粒。
已有學者致力于研究菌株中的乳糖分解代謝途徑中的酶是由哪一種質粒基因調控的。有研究得出在乳酸鏈球菌C2(Str.lactis C2)和乳脂鏈球菌B1(Str.cremoris B1)菌株里磷酸稀醇式丙酮酸依賴性磷酸轉移酶(PTS)系統中用于糖的攝取的酶Ⅱ、因子Ⅲ和乳糖分解酶-磷酸-β-半乳糖苷酶是由質粒編碼基因調控的[9]。有學者采用乳糖陰性突變的方法證實乳酸鏈球菌11454(Str.lactis 11454)菌株乳糖質粒攜帶有關編碼乳糖磷酸轉移酶和磷酸-β-半乳糖苷酶的基因。另有學者發現D-塔格糖-6-磷酸途徑中的3種酶(塔格糖-6-磷酸異構酶、D-塔格糖-6-磷酸激酶、塔格糖-1,6-二磷酸醛縮酶)的基因與乳酸鏈球菌Str.lactis C10、H1和133菌株的乳糖質粒相關。干酪乳桿菌64H(Lb.casei 64H)也被研究發現其中的乳糖質粒編碼乳糖磷酸轉移酶系統中的酶和磷酸-β-半乳糖苷酶[10]。半乳糖的利用也受乳糖質粒的消失的影響。目前已獲知乳酸鏈球菌(Str.lactis)的乳糖陰性突變體減緩了乳糖的增長率。
由于乳酸鏈球菌是營養依賴型微生物,而牛奶中缺乏足夠的游離氨基酸或小的肽類,需要將牛奶中酪蛋白分解,因此發酵劑的生長和產酸量依賴于發酵劑細胞壁上蛋白酶分解酪蛋白的能力。失去這種分解蛋白的能力會導致產酸量的減少。McKay等人[11]和Klaenhammer等人[12]證實質粒消除法可以得到這種產酸量少的突變體。蛋白酶質粒已在一些乳脂鏈球菌(Str.cremoris)中鑒定出來。
乳酸鏈球菌(Str.lactis)的蛋白酶質粒鑒定有很多不確定性,主要是因為該表型特點與乳糖基因調控之間存在一定的相關性。在乳酸鏈球菌(Str.lactis C2、ML3、712、C10、M18)和雙醋酸乳酸鏈球菌(diacetylactis DRC1)中,均表現乳糖和蛋白酶表型特征的同時消失與一個大小為30~45 Mdal的質粒相關,同時也發現了一些與之不同的現象,如保留了乳糖基因的乳酸鏈球菌C2(Str.Lactis C2)蛋白酶陰性突變體丟失了大小為12.5Mdal和18Mdal的質粒而保留了30Mdal的質粒分子[12]。近期有學者進行了高頻率的刪除組織構架和對一種未知的質粒的高頻率消除實驗[13],對解釋這些復雜現象起到了一定的作用。首先,很容易從乳酸鏈球菌712(Str.lactis 712)消除一個9Mdal的質粒,而大多數這些消除質粒的菌株通常表現為乳糖或蛋白酶陰性突變體。這個9Mdal的質粒并不調控乳糖基因或蛋白酶基因,但卻對這2個基因的表型有極大的影響,由此可以看出質粒與表型表現相關性是有條件的。第2,乳酸鏈球菌712(Str.lactis 712)的乳糖和蛋白酶質粒受支配于刪除組織構架的頻率。根據分子出現的位置的不同,通過消除實驗可以得到3種表型的菌株,分別是Lac-Prt-、Lac-Prt+和Lac+Prt-。再者,消除小于等于10%的質粒DNA并不能通過瓊脂糖凝膠檢測出來。pLP712的轉導實驗中,為了接受來自噬菌體頭部的DNA需要破壞受體的組織結構,根據破壞位點的不同得到了2種轉導產物,Lac+Prt+和Lac+Prt-。另外將乳酸鏈球菌(Str.Lactis 712、ML3、C2)中的乳糖質粒接合轉移到非攜帶質粒菌株乳酸鏈球菌712(Str.Lactis 712)中,3種菌株轉移后均得到了Lac+Prt+和Lac+Prt-兩種表象的菌株,并由限制性內切酶進行酶解,對這些質粒進行了對比,Lac+Prt-型菌株攜帶了1個了與蛋白酶質粒類似的質粒但是該質粒缺少了1個編碼蛋白酶的區域。乳酸鏈球菌(Str.Lactis)菌株似乎通常攜帶了1個同時編碼乳糖基因和蛋白酶基因的質粒,但是消除實驗可能破壞了該質粒與2種表現型之間的遺傳關系。
1.4 編碼表型的其他質粒
隨著乳糖和蛋白酶基因的研究,雙醋酸乳鏈球菌(Str.Lactis sub.diacetylactis)菌株發酵檸檬酸(Cit)的能力也被發現,Kempler和McKay[14]采用吖啶橙消除法分離了Cit-突變體和菌株18-16、DRC1。另外,拮抗控制質粒也得到了廣泛的研究,Neve等人使用消除法和接合轉移實驗鑒定了乳脂鏈球菌(Str.cremoris)菌株4G6和9B4及雙醋酸乳鏈球菌6F7(Str.Lactis sub.diacetylactis 6F7)中的細菌素質粒[15]。目前,已成功分離了乳酸鏈球菌 354-07(Str.lactis 354-07)編碼葡萄糖和甘露糖基因大小為23Mdal的質粒和乳酸鏈球菌DR1253(Str.Lactis DR1253)中的木糖質粒。并成功從1株乳酸鏈球菌71(Str.lactis 71)中將卡鈉徽素抗性質粒轉進了枯草芽孢桿菌(Bacillus sbtilis)中,這是第1個關于在乳鏈球菌中存在藥物抗性質粒的報告。無機離子抗性和無機離子敏感型特性被鑒定是由質粒編碼調控的。乳酸鏈球菌C2(Str.lactis C2)的乳糖質粒控制砷酸鹽、亞砷酸鹽和鉻酸鹽抗性和對銅的敏感特性,乳脂鏈球菌Wg2(Str.cremoris Wg2)中的1個6.1Mdal大小的質粒可能調控對銅的抗性特征[16]。菌株的其他特征也可能是質粒調控的,如對乳過氧化物酶-硫氰酸鹽-過氧化氫系統抗性、乳脂鏈球菌(Str.cremoris)菌株菌毛的生長和雙醋酸乳鏈球菌(Str.lactis sub.diacetylactis)菌株精氨酸水解的調控,仍需進一步研究。
2 基因轉移過程
2.1 轉導
第1個比較詳細的有關乳鏈球菌基因轉移的敘述是乳酸鏈球菌C2(Str.lactis C2)中的噬菌體介導的轉導[17]。更早的轉導相關報道還有乳酸鏈球菌C2(Str.lactis C2)中的鏈霉素抗性基因的轉導和雙醋酸乳鏈球菌18-16(Str.lactis sub.diacetylactis 18-16)中的色氨酸獨立性轉導,采用的是烈性噬菌體,而McKay關于乳酸鏈球菌C2(Str.Lactis C2)的研究和隨后的所有通過轉導進行基因轉移的實驗都是采用溫和的噬菌體。
McKay等人對乳酸鏈球菌C2(Str.Lactis C2)中染色體標記的麥芽糖和甘露糖基因和質粒調控的乳糖基因轉導進行了檢測[18]。在后續實驗中發現選取的乳糖轉導子供體中大概有50%的轉導子伴隨著非選擇性的蛋白酶基因的轉導。使用噬菌體對一個乳酸鏈球菌C2(Str.Lactis C2)乳糖轉導子基因進行了轉導重復實驗,發現乳糖基因的轉移頻率顯著增加了,該現象同樣出現在了其它轉導系統中,這樣的轉導稱為高頻轉導(HFT)。除乳糖質粒外,有學者對一個來自于糞鏈球菌(Str.faecalis)大小為17Mdal的紅霉素抗性質粒進行了轉導,但并未發現高頻轉導(HFT)現象[19]。
近來,有學者成功將由溫和噬菌體介導的乳脂鏈球菌C3(Str.cremoris C3)的乳糖基因分別轉移到了乳酸鏈球菌(Str.lactis ML3)和一個非攜帶質粒的乳酸鏈球菌 C2(Str.lactis C2)突變體中[20]。
轉導這種基因轉移方式是有限制的,只有具備相應噬菌體受體的宿主菌株才可以用來進行基因轉移,同時噬菌體的頭部容量限制了轉移的DNA的數量,而頭部容量的限制會影響遺傳物質的結構。那些相對于噬菌體頭部來說分子太大難以被全部容納的基因,乳糖質粒的轉導過程中會引起基因缺失的產生[13]。這種基因缺失的產生對于質粒編碼基因在限制酶切割圖譜中的分析是非常有用的。
此外,轉導有時會引起乳糖和蛋白酶質粒基因整合到細菌染色體上,產生了2個非常重要的現象,即基因通過轉移到染色體上而使遺傳變得非常穩定,另外,降低了基因表達水平[20]。有學者證實染色體上整合蛋白酶基因的菌株乳酸鏈球菌C2(Str.lactis C2)減少了蛋白水解能力,由此降低了苦味物質的產生,從而得到了由此菌株發酵的風味更佳的奶酪產品[21]。
2.2 接合轉移
了解乳鏈球菌中通過交配過程進行的基因轉移要求對細胞與細胞間的接觸有深入的研究。最早的研究是關于乳酸鏈球菌712(Str.lactis 712)之間的質粒編碼乳糖基因的接合轉移和由雙醋酸乳鏈球菌18-16(Str.lactis sub.diacetylactis 18-16)菌株的質粒轉移到非攜帶質粒乳酸鏈球菌C2(Str.lactis C2)突變體的研究[22]。隨后檢測出更多不同菌株間的乳糖質粒的轉移,如從乳酸鏈球菌ML3(Str.lactis ML3)、乳酸鏈球菌C2(Str.lactis C2)、乳脂鏈球菌C3(Str.cremoris C3)和雙醋酸乳鏈球菌(Str.Lactis sub.diacetylactis DRC3,11007,WM4)轉移到同樣突變的乳酸鏈球菌C2(Str.lactis C2)中,從乳脂鏈球菌(Str.cremoris R1,EB7,28,C3)中轉移到突變的乳酸鏈球菌ML3(Str.lactis ML3)中,從乳脂鏈球菌C3(Str.cremoris C3)中轉移到乳脂鏈球菌B1(Str.cremoris B1)突變體中。一個來自干酪乳桿菌64H(Lb.casei 64H)菌株的乳糖質粒已經被證明具有接合轉移的能力[10]。目前已在一些交配實驗獲得的雜合體菌株中分離出了高頻率的轉移供體菌株。這樣的高頻突變菌株最早在乳酸鏈球菌712(Str.lactis 712)菌株中發現,這些菌株表現了新的細胞聚集現象,該現象使得菌株的菌落形態和在培養基上的生長特征發生了顯著的變化。在乳糖質粒從乳酸鏈球菌ML3(Str.lactis ML3)轉移到一個非攜帶質粒的乳酸鏈球菌C2(Str.lactis C2)突變體過程中,也發現了高頻的子菌株。有學者將從高頻子菌株乳脂鏈球菌(Str.cremoris C3,Z8)中得到乳糖質粒及一代乳酸鏈球菌ML3(Str.lactis ML3)中的質粒轉移到了另一株乳酸鏈球菌ML3(Str.lactis ML3)宿主菌株中,進行對比,發現由于插入了新的DNA片段,這些由高頻接合轉移得到的子菌株包含了擴大的乳糖質粒[23-24]。從進化的觀點來看,細胞聚集也是復雜的誘導共軛系統中的一個現象并在一些糞鏈球菌(Str.faecalis)菌株中發現此共軛系統的作用。
開始時僅作為糞鏈球菌DS-5(Str.faecalis DS-5)[25]中的特殊質粒的紅霉素抗性質粒pAMβ1自發遺傳到了不同種類的革蘭氏陽性種和屬的細菌中,成為一種在體內發展和載體構建方面具有重要作用的質粒。該質粒首次從一個溶菌素缺陷菌株糞鏈球菌DS-5(Str.faecalis DS-5)中轉移到了乳酸鏈球菌712(Str.lactis 712)中,隨后該質粒從乳酸鏈球菌712(Str.lactis 712)中被轉移到了其他乳酸鏈球菌(Str.lactis),乳脂鏈球菌(Str.cremoris),雙醋酸乳鏈球菌(Str.Lactis sub.diacetylactis)和嗜熱鏈球菌(Str.thermophilus)中,此外,該質粒還被轉移到唾液乳桿菌(Lb.salivarius),嗜酸乳桿菌(Lb.acidophilus),干酪乳桿菌(Lb.casei),羅伊乳桿菌(Lb.reuteri),枯草芽孢桿菌(Bacillus subtilis)和金黃色葡萄球菌(Staphylococcus aureus)中。轉移過程中也會遇到一些問題,如一些轉移頻率是非常低的,還有些轉移是無法被檢測到的,解決這些問題的一種方法就是分離受體能力更強的突變體,該方法已成功應用于提高pAMβ1轉移到嗜熱鏈球菌1526(Str.Thermophilus 1526)中的轉移頻率,每一個受體轉移頻率從4.2×10-7提高到2.9×10-2。此外將一個分布廣泛的可遺傳的藥物抗性質粒作為對比進行了試驗,Gonzalez& Kunka[6]成功將氯霉素和紅霉素抗性質粒pIP501轉移到了戊糖片球菌(Pediococcus pentosaceus)和乳酸片球菌(Pediococcus acidilaciti)菌株中,質粒pAMβ1在這些菌株中卻不能成功轉移。很多細菌素質粒、控制蔗糖利用質粒、nisin和雙球菌素產生質粒類似基因模塊可以通過接合轉移來影響他們自身的轉移。
接合轉移在構建新的乳制品發酵劑菌株方面具有廣泛應用潛力。該方法沒有轉移DNA容量的理論極限。pAMβ1接合轉移的混雜性質使得在差異較大的菌株間的轉移成為可能。乳糖質粒可以轉移非自身的可遺傳的乳酸鏈球菌(Str.lactis)菌株中的隱秘的質粒。另外還發現了一種新型的具有潛力的接合轉移,即轉座子,并在糞鏈球菌(Str.faecalis)菌株中得到驗證。可遺傳的四環素抗性轉座子Tn916被成功轉移到了乳酸鏈球菌(Str.lactis M1中),這種在乳鏈球菌中結合了轉位技術的新型基因轉移系統具有廣闊的開發應用前景。
2.3 轉化和轉染
將純化的DNA導入可繁殖細菌這一過程對于基因轉移有重要的意義,因為它是基因克隆技術出現的前提條件。目前還不能將整個細胞倒入繁殖細菌,主要是將DNA導入細菌原生質體的方法進行轉化。原生質體溶解和再生主要依賴細胞壁的溶解,溶解使用的酶有溶菌酶、變溶菌素或結合的淀粉酶和溶菌酶。目前為止,僅有Kondo和 McKay[26]使用的聚乙二醇轉化實驗成功,他們對乳酸鏈球菌ML3(Str.lactis ML3)原生質體進行處理,引入了一個縮短的乳糖質粒,目前已經成功獲得了大約105/μg DNA轉化子的頻率。
2.4 原生質體融合
通過聚乙二醇來融合原生質體已成功實現了基因轉移。有學者使用帶有遺傳標記的乳酸鏈球菌712(Str.lactis 712)間的質粒和染色體基因進行轉移。原核生物的原生質體融合是一種特殊基因轉移過程,因為它是非特異性且包含整個基因組的轉移而不是部分DNA的單向轉移,這對于開發新型發酵劑菌株來說是一種非常有效的技術。針對發酵劑菌株用途的偏好性進行遺傳是非常困難的,而且該性質特也可能是被多基因控制的,有可能分布在染色體周圍。但通過原生質體融合技術可以得到混合的基因遺傳產物,此項技術將會使這種復雜的性狀轉移在一定程度上成為可能。
2.5 基因克隆技術
基因克隆技術是解決遺傳復雜性的有效手段之一。它為分離一個單獨組織的基因提供了可能,也為引入這些基因來構建一個新的發酵劑菌株提供了可能。有學者將氯霉素和紅霉素抗性基因克隆到一個乳脂鏈球菌Wg2(Str.Cremoris Wg2)小的隱蔽性質粒中,并成功獲得了一個抗性載體。這種雜交分子具有特殊的限制酶BclⅠ和BglⅡ的酶切位點,從而可以利用插入失活這一特性。該質粒不僅可以在枯草芽孢桿菌(Bacillus subtilis)中復制,也可以在大腸桿菌(Escherichia coli)復制,它可以在多宿主細胞中作為穿梭載體,這一性質是非常有價值的。基因工程菌株還未用于食品和乳制品工業相關應用中,有人認為將含藥物抗性基因的傳統克隆載體直接在食品工業上使用是不安全的,所以有必要使用乳鏈球菌和乳桿菌等原始乳制品相關菌株的代謝質粒進行載體的構建。目前有許多實驗室正使用現有的基因工程構建系統克隆,從而進行合適的代謝質粒的分子分析及相關發酵劑基因的分離。
3 質粒和基因轉移的分子學研究
限制酶圖譜是研究質粒分子之間的關系及發生變化的有效手段。結合缺失定位法和克隆可以將特殊的基因功能植入DNA中,同時可以推進對基因轉移過程的分子生物學分析。
3.1 限制酶圖譜
目前已經繪出了已知代謝功能的多種質粒和可以進行轉染的噬菌體的限制酶圖譜。圖1顯示的是雙醋酸乳鏈球菌176(Str.lactis subsp.diacetylactis 176)的檸檬酸質粒pCT176限制酶圖譜,該質粒與其它菌株的檸檬酸質粒大致是一樣的,并且可以用作代謝載體的基礎。這是惟一一個了解其功能及了解其在簡單鑒別培養基中生長中表型特征的多拷貝質粒,該質粒的限制酶圖譜顯示了特殊的臨近的配對位點,這些位點對應的限制酶有 Bam HI、BclⅠ、BglⅡ、ClaⅠ、Eco RⅠ和XhoⅠ,所有這些限制酶都可用于基因克隆。
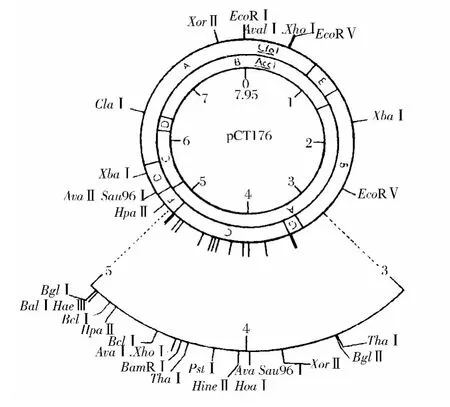
圖1 檸檬酸質粒pCT176限制酶切圖譜Fig.1
Loof等人發現噬菌體 P008基因組包含一個19.6Mdal具有粘性末端的雙鏈DNA,并繪制了它的限制酶圖譜,發現了限制酶BclⅠ、BglⅡ和HindⅢ的特殊的位點[27]。因為該噬菌體的DNA可以被轉移進入原生質體,所以它可作為一個有潛力的噬菌體載體。
有學者采用38種不同的限制酶對來自乳酸鏈球菌712(Str.lactis 712)的55kb的pLP712質粒進行了分析,并且繪制出了綜合圖譜[13]。圖譜中顯示了pLP712中的消除位點,圖譜中還將控制乳糖利用、蛋白酶生產和質粒復制的區域劃分出來。Kok等人使用7種不同的限制酶繪制了來自乳脂鏈球菌Wg2(Str.Cremoris Wg2)的17Mdal的蛋白酶質粒[28]。
3.2 乳酸菌的克隆基因
目前已有學者成功進行了發酵劑菌株在宿主細胞中的的基因表達和克隆。使用鏈狀球菌的載體pDB101將來自乳酸鏈球菌C2(Str.lactis C2)菌株的乳糖基因克隆進了血鏈球菌Challis(Str.sanguis Challis)菌株的乳糖陰性突變體中。被克隆的乳酸鏈球菌(Str.lactis)基因補充了新宿主細菌的磷酸-β-半乳糖苷酶突變和磷酸轉移酶系統突變。Lee等人[10]構建了干酪乳桿菌64H(Lb.casei 64H)乳糖質粒大腸桿菌(E.coli)基因庫并且成功分離了一株表達磷酸-β-半乳糖苷酶基因的克隆菌株,檢測了酶活性,同時用大腸桿菌(E.coli)微小細胞驗證了放射性同位素標記蛋白的存在。有學者試圖克隆pLP712質粒中控制蛋白酶活性的基因,使用多拷貝大腸桿菌(E.coli)載體pAT153,通過分離缺失圖中片段,但并未成功。另也曾試圖克隆乳脂鏈球菌(Str.cremoris)質粒pWV05的蛋白酶基因到大腸桿菌(E.coli)中,亦未能成功,原因可能為在大腸桿菌(E.coli)中蛋白酶的表達是致死的,目前研究者已開始研究替代大腸桿菌(E.coli)的方法來克隆蛋白酶基因。
3.3 轉導的分子生物學分析
有學者對乳酸鏈球菌(Str.lactis C2、712、ML3)中的乳糖基因進行轉導,創造了新的乳糖質粒,該質粒與噬菌體基因組大小匹配,這些質粒在轉移實驗中增加了轉導的頻率,此外,這些質粒可能攜帶了蛋白酶基因[17]。有學者使用乳酸鏈球菌(Str.lactis 712)的限制酶分析對噬菌體DNA、pLP712質粒DNA和一些Lac+Prt+、Lac+Prt-轉導質粒進行了對比。轉導質粒含有部分缺失的pLP712質粒DNA,而未攜帶噬菌體DNA。Lac+Prt+轉導產物具有單一的基因缺失,并表現一定的一致性。相反,Lac+Prt-轉導產物是不一致的,并且含有兩個或多個基因缺失。天然乳糖質粒pLP712是不穩定的,而且很容易發生缺失現象[13]。對溶菌原進行誘導后,一個大小合適的自發缺失的乳糖質粒也許會被錯誤的轉入噬菌體頭部,缺失的乳糖質粒可能就會進行轉導,采用pLP712的限制酶圖譜和缺失圖譜可以進一步解釋Lac+Prt-轉導產物多個基因缺失發生的原因。
在對乳糖質粒進行轉導之后,有學者對包含隱蔽性質粒補體的乳酸鏈球菌(Str.lactis 712)菌株的乳糖質粒進行了轉導實驗[19],同時也進行了插入染色體的實驗,實驗后發現了大的新的乳糖質粒,大小分別是44Mdal和55Mdal。在這些大的乳糖質粒轉移進入非攜帶質粒的菌株后,對其進行了分子生物學分析,發現它們是通過與一個受體隱蔽性質粒重組轉導后形成的。
3.4 乳糖質粒接合轉移的分子生物學分析
有學者將來自乳酸鏈球菌(Str.lactis ML3)的乳糖質粒接合轉移進入一個非攜帶質粒的乳酸鏈球菌(Str.lactis C2)突變體和非攜帶質粒的乳酸鏈球菌(Str.lactis 712)后,產生了變異的菌株,變異的菌株具有高供體能力,并且表現為細胞聚集型組成型表達。對這些菌株進行堿變性裂解實驗后發現這些菌株具有擴大的乳糖質粒[23],同樣有些表型不變的菌株也具有擴大的質粒。有學者對這2種擴大的質粒分別與Str.lactis ML3的天然乳糖質粒進行了對比,擴大的分子包含了完整的乳糖質粒,但插入了新的DNA 分子[24]。
Anderson和McKay[5]使用重組缺陷的乳酸鏈球菌(Str.lactis ML3)菌株驗證了插入過程與宿主細胞生成的重組系統是相互獨立的,并繪制了大量的擴大的乳糖質粒的限制酶圖譜,這些乳糖質粒有些具有表型變化,有些不具有表型變化,并發現這些新的DNA的原始結構都是環形的,可以在乳糖質粒的固定位置以不同的指向自發的進行融合。插入的環形分子的融合位置決定了雜交的質粒是否表現聚集型或(和)高頻轉導。該項研究表明可能存在乳糖編碼的轉移因子以特定的方式編碼生成不同表型的擴大質粒。
有學者通過在pLP712中引入兩個缺失位點而形成一個23.7kb微小乳糖質粒,該質粒小分子結構更適合于分離和對插入DNA的分析。對大量的擴大的質粒進行限制酶分析發現,插入位點發生在pLP712限制酶圖譜中一個3kb大小的固定位點,正如Anderson和McKay研究的一樣,插入的DNA不同但表現了一定的相關性。已成功繪制乳酸鏈球菌(Str.lactis 712)中環狀分子插入DNA的圖譜,觀察到插入的DNA具有一個相同的末端和一個不同的末端,從而決定了插入DNA分子大小的不同和插入系譜成員的不同。目前亟待解決的問題之一是新插入DNA片段來自哪里,有學者分析該片段可能是一個難以分離的質粒,也可能是一個位于細菌染色體上的轉座轉移因子或者兩者都有[24]。
3.5 乳鏈球菌體內DNA重排
當一個給定的乳鏈球菌的DNA包含一個細菌染色體和一個補體質粒時,DNA在體內的排列過程會發生改變。乳酸鏈球菌(Str.lactis 712)的乳糖質粒缺失是非常容易發生的,在通過吖啶黃或升高溫度處理、原生質體再生、重復培養傳代、接合轉移之后,通過轉導可以被篩選出來。乳酸鏈球菌(Str.lactis 712、C2)和乳脂鏈球菌(Str.cremoris C3)乳糖質粒的乳糖和蛋白酶基因可以在轉導后插入到到染色體中。在接合轉移過程中有時可以通過插入新的DNA來構建擴大的乳糖質粒。有學者已成功將一個擴大的乳糖質粒的乳糖基因轉移到了一個8.5kb的缺乏重組系統的Str.lactis ML3突變體隱蔽性質粒中[29]。在這些實驗中發現可以將DNA插入到一個乳糖質粒的位點上來構建通過接合轉移生成的擴大的乳糖質粒,所有從非攜帶質粒的乳酸鏈球菌(Str.lactis 712)菌株中篩選出來的Lac+Prt+質粒都具有相同的消除位點,95%通過原生質體融合生成的Lac+Prt-質粒消除位點也是相同的。由這些有序的分子學相互作用可以看出在乳鏈球菌的基因組中可能存在插入序列和轉座子。Neve等人[15]發現細菌素質粒與多種隱蔽性質粒之間有較弱的雜交現象。
4 結論
乳酸菌遺傳學將會應用到強化和創造新的發酵劑菌株當中,目前已經發現了兩條質粒調控的噬菌體抗性、限制修飾和吸附阻塞機制。控制這些特性的基因被引用到不同的發酵劑菌株上,插入到已存在的發酵劑菌株質粒上或通過基因克隆分離后的質粒上,并可以由此獲取一套對噬菌體侵染防御更強的發酵劑菌株防御系統。通過插入染色體的方法,已成功獲取了乳酸鏈球菌(Str.lactis C2和712)中穩定的乳糖和蛋白水解酶,這有利于控制蛋白水解酶及它在奶酪成熟過程中發揮的作用,改善它在干酪制作過程中產生異味物質,并且還有可能控制基因的過表達。可以通過分離藥物抗性菌株的方法進行改善奶酪中存留抗生素而導致發酵失敗的現象。通過原生質體融合技術可以將抗冷凍基因轉移到新的菌株中,從而增加直投式發酵的應用范圍。基因克隆技術的發展使乳酸菌在工業領域的應用取得了更大的突破,如小牛皺胃酶,該基因已成功在大腸桿菌(E.coli)和酵母中克隆。
過去的10年間,乳制品乳酸菌已經在遺傳工程和微生物群研究方面取得了重大的突破。在接下來的幾十年中將會看到分子生物學技術在實踐中的應用及遺傳工程在乳酸菌中的應用,對近年來難以攻克的遺傳現象將會有更加深入的理解。這些已經成熟的傳統遺傳學分析手段將會在不久的將來用于生產乳業工業中新型的發酵劑菌株,例如質粒轉移會應用到生產噬菌體抗性更強或具有新的蛋白水解酶活性的菌株當中。遺傳學研究的未來是被肯定的,我們已經進入到了乳業生物技術的時代中。
[1] 吳妍妍,張文舉,胡猛,等.乳酸菌在動物生產中應用的研究進展[J].中國畜牧獸醫,2013(02):221-224.
[2] 陳琦,馬燕芬,王利.乳酸菌基因組學與干酪風味的關系[J].中國乳品工業,2011(10):40-43.
[3] 王俊滬,霍貴成.乳球菌的功能性質粒[J].中國乳品工業,2006(05):43-51.
[4] McKay L L,Baldwin KA,Zottola EA.Loss of lactose metabolism in lactic streptococci[J].Appl Microbiol,1972,23(6):1 090-1 096.
[5] Anderson D G,McKay L L.Simple and rapid method for isolating large plasmid DNA from lactic streptococci[J].Appl Environ Microbiol,1983,46(3):549-552.
[6] Gonzalez C F,Kunka BS.Plasmid transfer in Pediococcus spp.:intergeneric and intrageneric transfer of pIP501[J].Appl Environ Microbiol,1983,46(1):81-89.
[7] Larsen L D,McKay L L.Isolation and characterization of plasmid DNA in Streptococcus cremoris[J].Appl Environ Microbiol,1978,36(6):944-952.
[8] Snook R J,McKay L L.Conjugal Transfer of Lactose-Fermenting Ability Among Streptococcus cremoris and Streptococcus lactis Strains[J].Appl Environ Microbiol,1981,42(5):904,911.
[9] McKay L L.Functional properties of plasmids in lactic streptococci[J].Antonie Van Leeuwenhoek,1983,49(3):259-274.
[10] Lee L J,Hansen J B,Jagusztyn-Krynicka E K,et al.Cloning and expression of the beta-D-phosphogalactosidegalactohydrolase gene of Lactobacillus casei in Escherichia coli K-12[J].Journal of Bacteriology,1982,152(3):1 138-1 146.
[11] McKay L L,Baldwin K A.Plasmid distribution and evidence for a proteinase plasmid in Streptococcus lactis C2-1[J].Appl Microbiol,1975,29(4):546-548.
[12] Klaenhammer T R,McKay L L,Baldwin K A.Improved lysis of group N streptococci for isolation and rapid characterization of plasmid deoxyribonucleic acid[J].Appl Environ Microbiol,1978,35(3):592-600.
[13] Gasson M J.Plasmid complements of Streptococcus lactis NCDO 712 and other lactic streptococci after protoplast-induced curing[J].Journal of Bacteriology,1983,154(1):1-9.
[14] Kempler G M,McKay L L.Characterization of Plasmid Deoxyribonucleic Acid in Streptococcus lactis subsp.diacetylactis:Evidence for Plasmid-Linked Citrate Utilization[J].Appl Environ Microbiol,1979,37(2):316-323.
[15] Neve H,Geis A and Teuber M.Conjugal transfer and characterization of bacteriocin plasmids in group N(lactic acid)streptococci[J].Journal of Bacteriology,1984,157(3):833-838.
[16] Otto R,de Vos WM and Gavrieli J.Plasmid DNA in Streptococcus cremoris Wg2:Influence of pH on Selection in Chemostats of a Variant Lacking a Protease Plasmid[J].Appl Environ Microbiol,1982,43(6):1 272-1 727.
[17] McKay L L,Baldwin K A.Simultaneous loss of proteinase-and lactose-utilizing enzyme activities in Streptococcus lactis and reversal of loss by transduction[J].Appl Microbiol,1974,28(3):342-346.
[18] McKay L L,Cords B R,Baldwin K A.Transduction of lactose metabolism in Streptococcus lactis C2[J].Journal of Bacteriology,1973,115(3):810-815.
[19] Davies F L,Gasson M J.Reviews of the progress of dairy science:genetics of lactic acid bacteria[J].Journal of Dairy Research,1981,48(2):363-376.
[20] Snook R J,McKay L L,Ahlstrand G G.Transduction of Lactose Metabolism by Streptococcus cremoris C3 Temperate Phage[J].Appl Environ Microbiol,1981,42(5):897-903.
[21] McKay L L,Baldwin K A.Stabilization of Lactose Metabolism in Streptococcus lactis C2[J].Appl Environ Microbiol,1978,36(2):360-367.
[22] Kempler G M,McKay L L.Genetic Evidence for Plasmid-Linked Lactose Metabolism in Streptococcus lactis subsp.diacetylactis[J].Appl Environ Microbiol,1979,37(5):1041-1 043.
[23] Walsh P M,McKay L L.Recombinant plasmid associated cell aggregation and high-frequency conjugation of Streptococcus lactis ML3[J].Journal of Bacteriology,1981,146(3):937-944.
[24] Walsh P M ,McKay L L.Restriction endonuclease analysis of the lactose plasmid in Streptococcus lactis ML3 and two recombinant lactose plasmids[J].Appl Environ Microbiol,1982,43(5):1 006-1 010.
[25] Clewell DB,Yagi Y,Dunny GM,et al.Characterization of three plasmid deoxyribonucleic acid molecules in a strain of Streptococcus faecalis:identification of a plasmid determining erythromycin resistance[J].Journal of Bacteriology,1974,117(1):283-289.
[26] Kondo J K and McKay L L.Transformation of Streptococcus lactis Protoplasts by Plasmid DNA[J].Appl Environ Microbiol,1982,43(5):1 213-1 215.
[27] Loof M,Lembke J,Teuber M.Characterization of the Genome of the Streptococcus lactis"subsp.diacetylactis"Bacteriophage P008 Wide-spread in German Cheese Factories[J].Systematic and Applied Microbiology,1983,4(3):413-423.
[28] Kok J,De Vos W,Vosman B,et al.Genetic marking of cryptic Streptococcus cremoris plasmids for the development of a transformation system[J].Lactic Acid Bacteria in Foods:Genetics,Metabolism and Applications,1983:64.
[29] Anderson D G,McKay L L.In Vivo Cloning of lac Genes in Streptococcus lactis ML3[J].Appl Environ Microbiol,1984,47(2):245-249.
