食鹽質量濃度對傳統自然發酵圓白菜的菌系結構和代謝的影響
2015-01-04 08:55:04李軍波關倩倩
食品科學 2015年11期
熊 濤,李軍波,彭 飛,關倩倩
食鹽質量濃度對傳統自然發酵圓白菜的菌系結構和代謝的影響
熊 濤,李軍波,彭 飛,關倩倩
(南昌大學食品學院,食品科學與技術國家重點實驗室,江西 南昌 330047)
以2、5、8 g/100 mL食鹽添加量的傳統自然發酵泡菜為研究對象,比較了3種不同食鹽質量濃度的傳統自然發酵泡菜中優勢菌系結構、鹵水pH值和總酸、底物和代謝產物、亞硝酸鹽含量的變化。結果表明:乳酸菌主導整個泡菜發酵過程,蔗糖一直被代謝利用,葡萄糖和果糖含量逐步增加,發酵結束時乳酸大量積累。食鹽質量濃度對泡菜發酵前期有顯著影響,2 g/100 mL食鹽質量濃度的泡菜中乳酸菌的繁殖代謝最快,發酵結束時pH值最低,總酸含量最高;5 g/100 mL食鹽質量濃度能較好地抑制有害微生物的繁殖,最快通過亞硝峰,產生的乙醇量最高;8 g/100 mL食鹽質量濃度抑制了泡菜中乳酸菌的繁殖代謝,使泡菜的成熟期延遲,對蔗糖的利用率最低。
泡菜;食鹽質量濃度;乳酸菌;發酵;代謝
我國傳統四川泡菜以自然發酵為主,是利用原料表面自帶的微生物在中低質量濃度鹽水(2~10 g/100 mL)里厭氧或兼性厭氧泡漬發酵,具有益生功能的乳酸菌主導著泡菜發酵過程[1]。現有研究發現鹵水鹽質量濃度不僅影響泡菜的咸味,還影響泡菜發酵過程中微生物的結構,從而直接和間接地影響泡菜的風味[2]。盡管有研究表明鹵水鹽濃度的改變能引起泡菜發酵過程中微生物的結構變化[3-4],泡菜發酵過程中鹵水pH值和總酸、還原糖以及亞硝酸鹽的變化規律已有研究[5-6],然而尚缺少鹵水鹽質量濃度與泡菜發酵過程中微生物的消長、底物消耗、代謝產物合成的關聯性研究。本實驗結合四川泡菜的食鹽質量濃度特點,研究3種不同食鹽質量濃度(2、5、8 g/100 mL)泡菜中鹵水的pH值及總酸、乳酸菌、酵母菌、多種糖類、有機酸、亞硝酸鹽的變化。通過比較傳統發酵泡菜在不同食鹽質量濃度下優勢微生物的消長規律、底物及代謝產物的變化,旨在揭示食鹽質量濃度對泡菜發酵特性的影響,期望對傳統自然發酵泡菜的工業化生產提供科學依據和理論指導。
1 材料與方法
1.1材料
圓白菜、食鹽、冰糖、生姜、大蒜、干辣椒、花椒等購于當地市場。
1.2試劑與培養基
氫氧化鈉、濃硫酸、無水對氨基苯磺酸、鹽酸萘乙二胺、亞鐵氰化鉀、乙酸鋅、硼砂、亞硝酸鈉(均為分析純)國藥集團化學試劑有限公司;蔗糖、葡萄糖、果糖、乙醇、檸檬酸、乳酸、乙酸標準品(均為色譜純)美國Sigma公司。
乳酸菌分離計數培養基:MRS培養基,參見GB 4789.35—2010《食品微生物學檢驗乳酸菌檢驗》;酵母菌分離計數培養基:虎紅瓊脂北京奧博星公司。
1.3儀器與設備
Agilent 1260型高效液相色譜美國安捷倫公司;Aminex-87H色譜柱美國Bio-Rad公司;PHS-25型pH計上海精密科學儀器有限公司;SP-725分光光度計上海普光儀器有限公司。
1.4 傳統泡菜制作方法

配方:圓白菜1 000 g、冷卻的開水2 000 mL、大蒜60 g、花椒30 g、辣椒80 g、生姜40 g、冰糖80 g、分別向冷卻的開水中加入40、100、160 g食鹽配制成2、5、8 g/100 mL的鹽水溶液。
1.5 分析方法
1.5.1取樣
制作好泡菜后每隔12 h在無菌條件下取樣泡菜液(10 mL),取樣前將泡菜液搖勻,取樣時間為7 d。
1.5.2優勢微生物的分離計數
取1 mL泡菜液加入到9 mL無菌生理鹽水中,適當稀釋后選擇3個合適的梯度稀釋液,分別取100μL涂布于MRS固體培養基和虎紅固體培養基,每個梯度做2次平行涂布。將MRS固體培養基置于37 ℃培養箱培養48 h,虎紅固體培養基置于30 ℃培養箱培養3~5 d后計數。
1.5.3 pH值和總酸含量的測定
用pH計測定鹵水pH值。總酸含量測定按GB/T 12456—2008《食品中總酸的測定》的方法測定總酸含量(g/L,以乳酸計)。
1.5.4亞硝酸鹽含量的測定
泡菜中亞硝酸鹽含量按GB/T 5009.33—2010《食品中亞硝酸鹽與硝酸鹽的測定》中鹽酸萘乙二胺法測定。
1.5.5發酵液中糖和有機酸含量的測定
將泡菜液在12 000 r/min條件下離心10 min,取上清液經0.22μm水系濾膜過濾,得到的濾液用于高效液相色譜分析。色譜條件:進樣量20μL,流動相為6 mmol/L硫酸溶液,流速為0.5 mL/min,柱溫45 ℃,紫外檢測器檢測波長205 nm[7]。選用示差折光檢測器檢測糖醇(蔗糖、葡萄糖、果糖、乙醇)含量,紫外檢測器檢測有機酸(檸檬酸、乳酸、乙酸)含量。
2 結果與分析
2.1 3 種食鹽質量濃度泡菜發酵過程中pH值和總酸含量的變化
pH值和酸度會影響到泡菜的風味,當鹵水pH值為3.5~3.8,即認為泡菜已成熟[8]。成品泡菜的pH值控制在3.0左右,乳酸控制在1%左右效果最佳[9]。有研究表明泡菜發酵過程的啟動菌以代謝周期短、耐酸性差的腸膜明串珠菌為主,代謝中后期是由耐酸性較強的同型乳酸菌(如:植物乳桿菌)主導[10-11]。在發酵的前3 d,乳酸菌快速繁殖并產生有機酸使鹵水pH值快速降低。
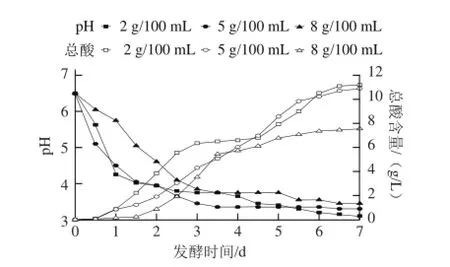
圖1 3 種食鹽質量濃度泡菜發酵過程中pH值和總酸含量的變化Fig.1 Changes in pH and total acid during the fermentation process ofpickled cabbage with three addition amounts of salt
如圖1所示,3種食鹽質量濃度的泡菜pH值在前1.5 d均急劇下降,然而5 g/100 mL食鹽質量濃度泡菜pH值先于另外2種食鹽質量濃度泡菜達到穩定(pH 3.3)。這可能是因鹵水鹽質量濃度不同產生的滲透壓不同所導致,1 g/100 mL食鹽溶液可以產生61.17 kN/m2的滲透壓,而大多數微生物細胞的滲透壓為30.17~61.15 kN/m2[12]。而2 g/100 mL食鹽質量濃度泡菜中可能存在一些不耐鹽的雜菌,與乳酸菌之間存在著一定的競爭關系,減緩了鹵水pH值的下降;8 g/100 mL的食鹽質量濃度產生的滲透壓遠高于乳酸菌的細胞液滲透壓,可能抑制了乳酸菌的繁殖和代謝,從而減緩了鹵水pH值的下降。發酵中后期鹵水中的乳酸和H+大量積累,影響了菌體產酸[13];3種不同食鹽質量濃度泡菜的pH值變化在發酵中后期下降緩慢。不同食鹽質量濃度的pH值變化并不相同,2 g/100 mL和8 g/100 mL食鹽質量濃度泡菜的pH值先出現短暫的穩定(pH 3.75)然后緩慢下降,而5 g/100 mL食鹽質量濃度泡菜的pH值一直維持在3.3左右。發酵結束時,2 g/100 mL食鹽質量濃度泡菜pH值最低(3.15),8 g/100 mL食鹽質量濃度泡菜pH值最高(3.45)。發酵過程中,5 g/100 mL食鹽質量濃度泡菜的酸度一直保持穩定的上升,最終總酸達到10.9 g/L;2 g/100 mL和8 g/100 mL食鹽質量濃度泡菜的酸度前期會快速升高然后出現一個相對穩定的過程,之后又緩慢上升,最終總酸含量分別達到11.2 g/L和7.6 g/L。
2.2 3 種食鹽質量濃度泡菜發酵過程中優勢微生物的變化
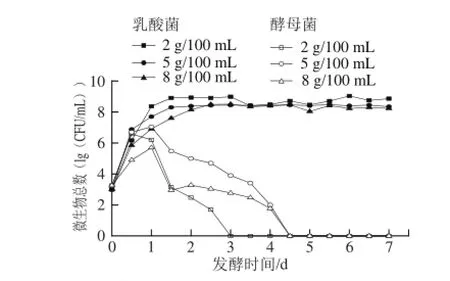
圖2 3 種食鹽質量濃度泡菜發酵過程中微生物總數的變化Fig.2 Changes in latic acid bacteria and yeast counts during the fermentation process of pickled cabbage with three addition amounts of salt
泡菜發酵初期不僅有有益微生物(如:乳酸菌、酵母菌和醋酸菌),還有有害微生物(如:丁酸菌、腐敗細菌、霉菌等),其中乳酸菌和酵母菌占主要部分[8]。如圖2所示,啟動發酵時乳酸菌數在103CFU/mL,且在2 d內快速生長至108CFU/mL,發酵中后期乳酸菌數始終保持在108CFU/mL。然而,在發酵前期乳酸菌在不同食鹽質量濃度下的生長情況存在較大差異,2、5、8 g/100 mL食鹽質量濃度泡菜中的乳酸菌含量分別在第1、1.5、2天時達到108CFU/mL;其中2 g/100 mL食鹽質量濃度泡菜在第1~3.5天均高于另外2種食鹽質量濃度泡菜,可能是2 g/100 mL的食鹽溶液產生的滲透壓對乳酸菌的代謝繁殖抑制作用最弱所致。在3種食鹽質量濃度泡菜發酵的中后期乳酸菌數始終保持在108CFU/mL,該結果與現有研究結果[8,14]一致。但發酵中后期5 g/100 mL食鹽質量濃度泡菜中乳酸菌數量保持比較穩定,而2 g/100 mL和8 g/100 mL食鹽質量濃度泡菜中乳酸菌數先升高后下降,可能是由鹵水的pH值和食鹽質量濃度的差異導致[13,15]。
泡菜發酵啟動時酵母菌數量為103CFU/mL,在發酵的第1天內均出現快速增長期。酵母菌在不同食鹽質量濃度的泡菜中繁殖速率有差異,2、5、8 g/100 mL食鹽質量濃度泡菜中的酵母菌數量分別在第0.5、1、1天時達到106、107、105CFU/mL,然后逐漸減少,最后消亡。可能是隨著異型乳酸菌和酵母菌代謝產生的二氧化碳造成了厭氧環境、鹵水pH值降低、酸度增加使酵母菌逐漸消亡[16-17]。2 g/100 mL食鹽質量濃度的泡菜中酵母在第3天消亡,而含鹽量為5 g/100 mL和8 g/100 mL泡菜的鹵水中酵母菌在第4.5天時才消亡,可能是在相對的高鹽環境下產生了一些耐鹽酵母[18-19]。這與曾駿等[20]研究認為“傳統四川泡菜發酵過程中酵母菌數量會先增加后減少,5 d后消失”的結果一致。
2.3 3種食鹽質量濃度泡菜發酵過程中底物含量的變化
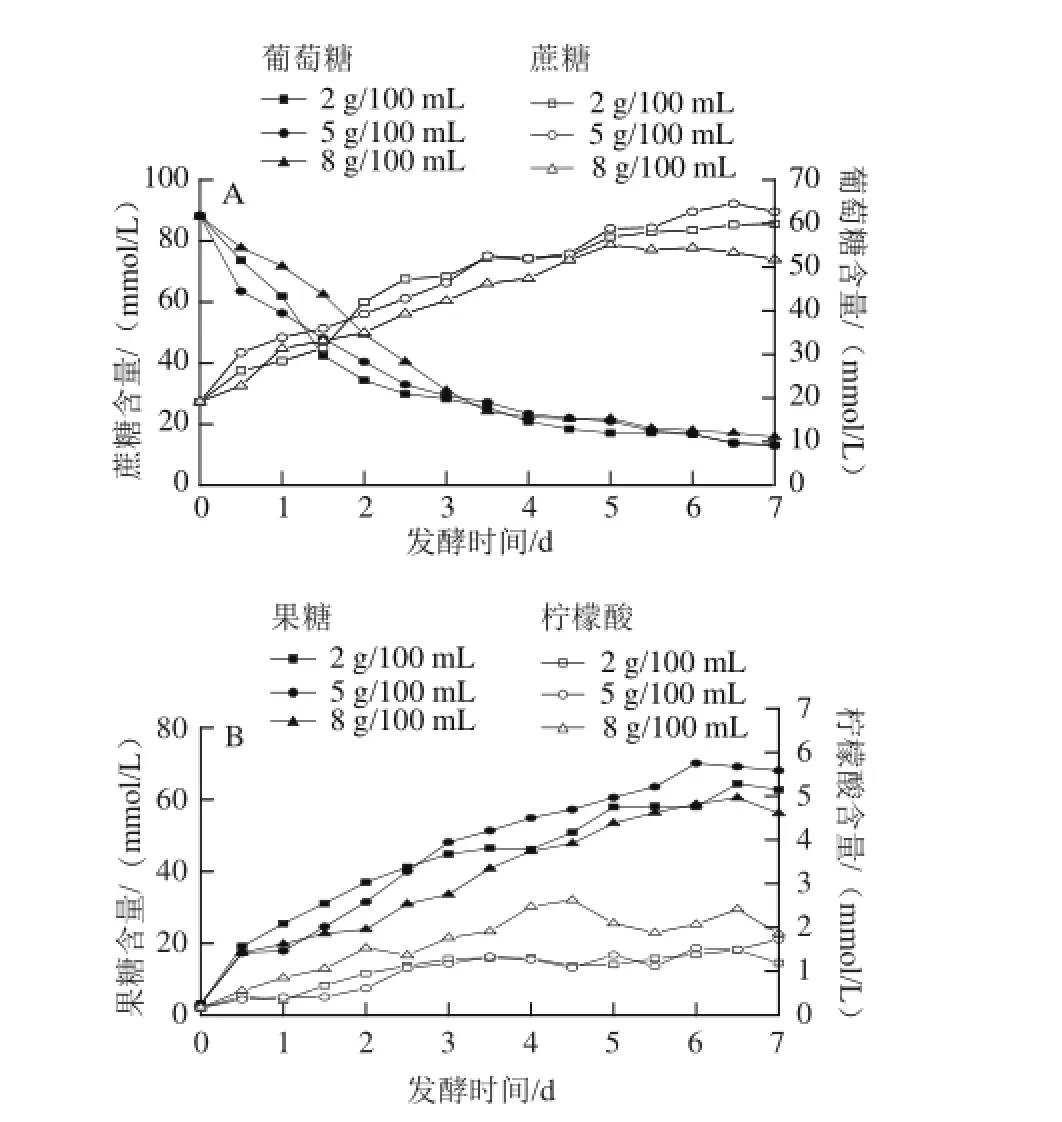
圖3 3 種食鹽質量濃度泡菜發酵過程中蔗糖、葡萄糖(A)、果糖、檸檬酸(B)含量的變化Fig.3 Changes in sucrose (A), glucose, fructose and citric acid (B) contents during the fermentation process of pickled cabbage with three addition amounts of salt
發酵過程中,蔗糖、葡萄糖、果糖為微生物提供碳源,其中的蔗糖主要源于發酵時加入的冰糖,蔗糖作為發酵的初始碳源,供給體系中微生物代謝。如圖3A所示,3種不同食鹽質量濃度泡菜鹵水中蔗糖在發酵前期被快速代謝,泡菜在發酵中后期對蔗糖的利用趨于穩定。泡菜發酵前期由腸膜明串珠菌主導,其可以產生高效的葡萄糖蔗糖酶,葡萄糖蔗糖酶能將蔗糖轉換化為葡萄糖和果糖[21];此外一些細菌(如:醋酸菌)和真菌(如:霉菌和酵母菌)的繁殖也會代謝蔗糖[21-22],導致了蔗糖前期被快速代謝。泡菜發酵中后期由同型乳酸發酵菌株(如:植物乳桿菌)主導,其對蔗糖的利用能力較異型發酵菌株弱[23];另一方面乳酸的大量積累、鹵水pH值的降低等抑制了菌的代謝活動,使蔗糖的利用減慢。然而在發酵的前2.5 d,2 g/100 mL食鹽質量濃度泡菜較另外2種泡菜蔗糖消耗量大,可能是在較低滲透壓條件下細菌、霉菌、酵母菌代謝較強,從而蔗糖的消耗較快。在泡菜發酵的中后期,食鹽質量濃度(2~8 g/100 mL)對菌株代謝蔗糖的影響較小,但發酵結束時,5 g/100 mL食鹽質量濃度泡菜中蔗糖利用率最高,8 g/100 mL食鹽質量濃度的最低。
鹵水中葡萄糖和果糖一部分源于蔗糖的分解,另一部分是從原料中滲透出來。如圖3所示,在整個發酵過程中葡萄糖和果糖的變化趨勢相似,在發酵的前5 d穩步上升,之后趨于穩定,這可能是在整個發酵過程中微生物對葡萄糖和果糖的消耗量低于產生量所致。其中5 g/100 mL食鹽質量濃度泡菜中葡萄糖和果糖的殘留量最多,可能是該食鹽質量濃度下的蔗糖利用率較高導致;8 g/100 mL食鹽質量濃度泡菜中葡萄糖和果糖的殘留量最少。檸檬酸是一種前體風味物質,對泡菜的風味存在重要的意義。3種食鹽質量濃度泡菜中檸檬酸含量在前期均穩步升高,之后處于穩定狀態。但不同食鹽質量濃度泡菜中的檸檬酸最高含量并不相同,其中2、5、8 g/100 mL食鹽質量濃度泡菜中檸檬酸的最高濃度分別為1.5、1.8、2.8 mmol/L,這可能是高滲透壓有利于圓白菜中的檸檬酸滲入到鹵水中導致。
2.4 3種食鹽質量濃度泡菜發酵過程中代謝產物產量的變化
乳酸是泡菜中主要的有機酸,其含量直接影響泡菜中菌系的穩定性。泡菜在發酵初期乳酸菌含量相對較低,由產酸能力較差的異型乳酸菌(如:腸膜明串珠菌)主導,在泡菜發酵的中后期乳酸菌含量較高,由耐酸性較強、產酸能力強的同型乳酸菌(如:植物乳桿菌)主導[24]。
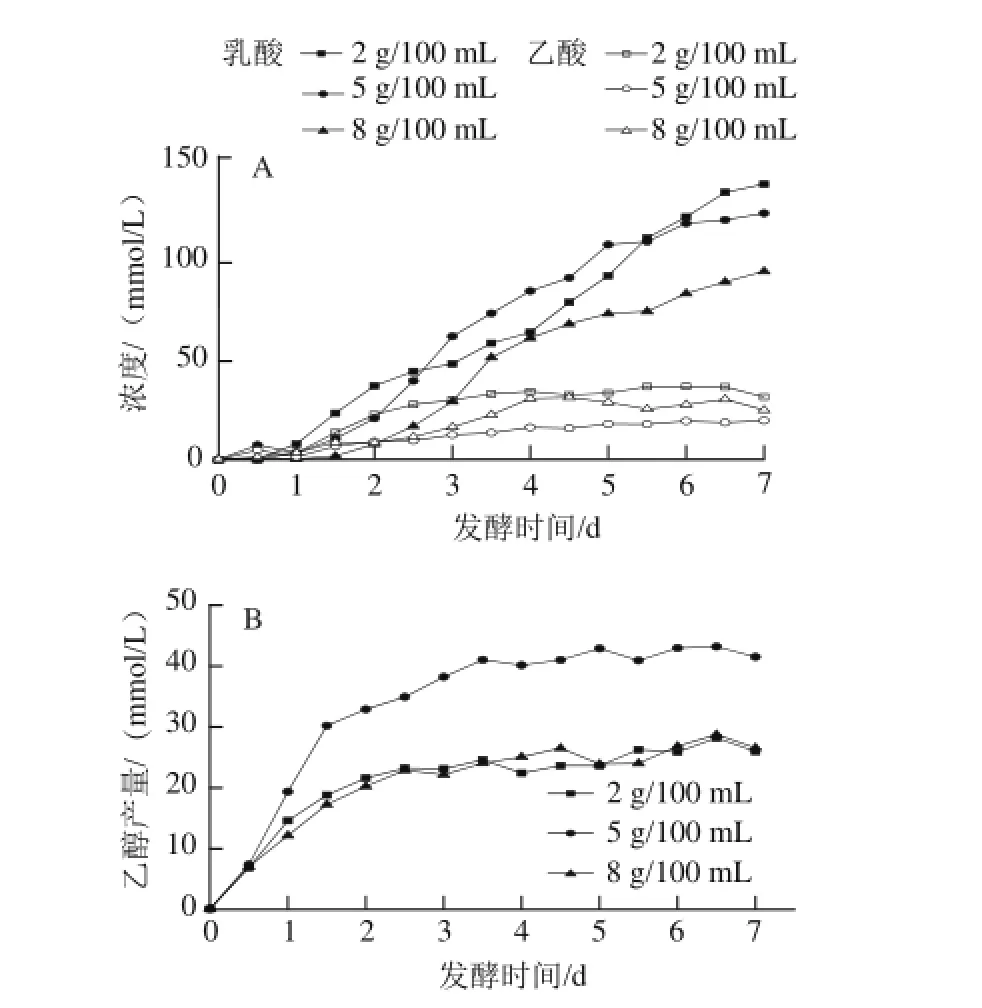
圖4 3種食鹽質量濃度泡菜發酵過程中乳酸和乙酸(A)、乙醇(B)產量的變化Fig.4 Changes in lactic acid, acetic acid (A) and ethanol (B) contents during the fermentation process of pickled cabbage with three addition amounts of salt
如圖4所示,乳酸在發酵前2 d緩慢升高,第3~5天快速積累,之后又緩慢升高。盡管乳酸菌在發酵的前2 d已生長至108CFU/mL,與此同時蔗糖也被快速消耗,然而乳酸的積累量并不大。一方面是因為泡菜發酵前期由產酸能力較弱的異型乳酸菌(如:腸膜明串珠菌)主導[10];另一方面是鹵水中存在較多能代謝蔗糖但不產乳酸的微生物(如:霉菌、酵母菌)。在泡菜發酵的中后期由產酸能力較強的同型乳酸菌(如:植物乳桿菌)主導[11],然而在隨著發酵的進行,鹵水中的乳酸和H+大量積累,乳酸菌需要消耗更多的能量來維持細胞內的pH值穩定,影響了菌體產酸[13],在發酵的第5~7天,乳酸積累較慢。其中2 g/100 mL食鹽質量濃度泡菜乳酸產量在第1~2.5天高于另外2種食鹽質量濃度泡菜,第3~5天5 g/100 mL食鹽質量濃度泡菜乳酸產量高于另外2種食鹽質量濃度泡菜,發酵后期乳酸的積累抑制乳酸菌生長和產酸,導致產乳酸速率下降。發酵結束時,2 g/100 mL食鹽質量濃度泡菜乳酸產量最高(140 mmol/L),8 g/100 mL食鹽質量濃度泡菜中乳酸產量最低(95 mmol/L)。總之,乳酸的產量與食鹽質量濃度呈反比關系,楊瑞鵬等[15]通過對不同食鹽質量濃度中乳酸菌區系的研究也證實了這一結論。
作為異型發酵乳酸菌的產物之一,適當濃度的乙酸可以有效地提高泡菜的感官性質[25]。異型乳酸發酵過程是葡萄糖經單磷酸己糖途徑降解,在有氧條件下發酵產生乙酸[26-27]。乙酸主要是在發酵第1.5~4天變化稍快,在發酵末期趨于穩定。不同食鹽質量濃度泡菜的乙酸含量并不相同,2 g/100 mL食鹽質量濃度泡菜的鹵水中乙酸產量最高,可能是2 g/100 mL食鹽質量濃度泡菜的乳酸菌含量在第1.5~4天均高于其他2種食鹽質量濃度泡菜,在此階段異型乳酸菌產生了較多乙酸所致。乙酸與乙醇結合產生乙酸乙酯能增強泡菜的風味。泡菜液中乙醇主要來源于酵母菌和異型乳酸菌,在發酵前期酵母菌數最高達到106CFU/mL,乳酸菌數達到108CFU/mL,發酵前期大多乳酸菌都是異型乳酸菌,在缺氧條件下異型乳酸菌發酵產生乙醇[27]。乙醇產量從發酵啟動開始到第2.5天急劇上升,在發酵中后期異型乳酸菌和酵母菌逐漸消亡[20,24],乙醇產量趨于穩定。此外,5 g/100 mL食鹽質量濃度泡菜中乙醇產量在發酵結束時達到43.2 mmol/L,遠高于另外2種食鹽質量濃度的泡菜。一方面是因為在發酵前期5 g/100 mL食鹽質量濃度泡菜中酵母菌數高于另外2種食鹽質量濃度的泡菜,而且酵母菌的下降量相對另外2種食鹽質量濃度的泡菜要緩慢,在第4.5天才消失;另一方面發酵啟動到第1天5 g/100 mL食鹽質量濃度泡菜中乳酸菌含量明顯高于另外2種食鹽質量濃度泡菜,在此階段異型乳酸菌代謝積累乙醇。
2.5不同食鹽質量濃度泡菜發酵過程中亞硝酸鹽含量變化
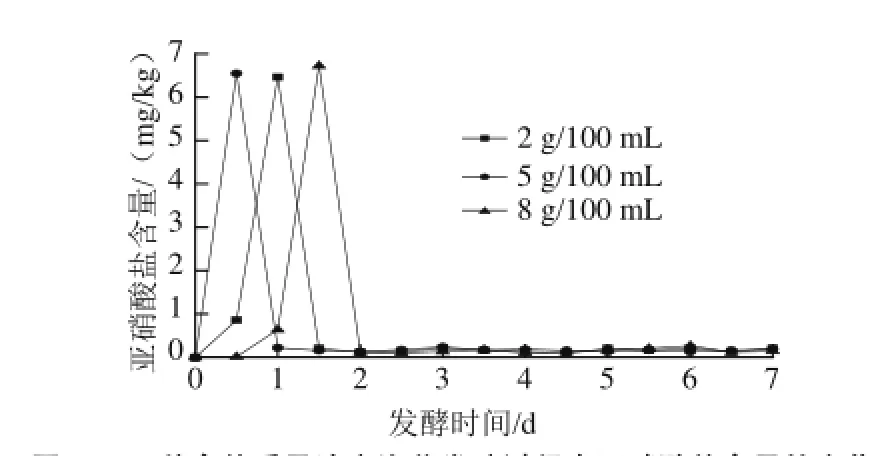
圖5 3 種食鹽質量濃度泡菜發酵過程中亞硝酸鹽含量的變化Fig.5 Changes in nitrite content during the fermentation process ofpickled cabbage with three addition amounts of salt
自然發酵泡菜是利用存在蔬菜表面的微生物群進行發酵的,這種方式生產出來的泡菜中亞硝酸鹽含量普遍偏高[28]。有研究表明硝酸還原酶的適宜pH值為7.10~7.15,發酵液pH值越小,亞硝酸鹽的降解速率越快[29]。如圖5所示,整個泡菜發酵過程鹵水中亞硝酸含量變化存在一定的波動,3種食鹽質量濃度的泡菜亞硝酸鹽含量的最高峰均在6.5 mg/kg左右,但不同食鹽質量濃度泡菜通過亞硝峰的時間不同,其中含5 g/100 mL食鹽質量濃度的泡菜最先通過了亞硝峰。一方面可能是因為含鹽量為5 g/100 mL的泡菜在0.5 d時pH值已經降至5.1,相對于其他食鹽質量濃度的泡菜pH值更低,從而對硝酸還原酶的抑制作用更加強烈;另一方面可能是5 g/100 mL的食鹽質量濃度能較好地抑制有害微生物的生長[2]。
3 結 論
通過對不同食鹽質量濃度的傳統自然發酵泡菜研究表明,乳酸菌在發酵前期快速繁殖,然后數量保持穩定;酵母菌數量先上升至106CFU/mL后逐漸消亡。食鹽質量濃度主要是對泡菜發酵前期有顯著影響,2 g/100 mL的食鹽添加量泡菜中產生的滲透壓最低,乳酸菌的繁殖代謝最快,酵母菌最先消亡;發酵結束時產生的乳酸含量最高(140 mmol/L),pH值最低(3.15),總酸含量最高(11.2 g/L)。5 g/100 mL的食鹽添加量能較好地抑制有害微生物的繁殖,最快通過亞硝峰,發酵結束時乙醇產量最高(43.2 mmol/L),蔗糖利用率最高,葡萄糖和果糖的殘留量最高,整個發酵過程中酸度穩定上升。8 g/100 mL食鹽添加量減緩了泡菜中乳酸菌的繁殖代謝,使泡菜的成熟期延遲,對蔗糖的利用率最低。食鹽質量濃度的改變引起鹵水滲透壓的變化,導致了發酵過程中菌系結構的不同,微生物的代謝出現差異,最終引起了泡菜中各指標的差異化。傳統自然發酵泡菜中適宜的食鹽添加量能提高泡菜品質,可進一步研究食鹽添加量對泡菜感官和風味的影響,進而為我國傳統自然發酵泡菜工業化生產提供可靠依據。
[1]陳功,夏有書,張其圣,等.從中國泡菜看四川泡菜及泡菜壇[J].中國釀造, 2010, 29(8): 5-8.
[2]楊瑞,張偉,陳煉紅,等.發酵條件對泡菜發酵過程中微生物菌系的影響[J].食品與發酵工業, 2005, 31(3): 90-92.
[3]付莎莉,陳安均,蒲彪,等.食鹽濃度對傳統四川泡菜發酵過程中乳酸菌菌相的影響[J].食品與發酵工業, 2013, 39(8): 102-107.
[4] ZHAO Dayun, DING Xiaolin. Studies on the low-salt Chinese potherb mustard (Brassica juncea, Coss.) pickle. I: the effect of a homofermentativeL(+)-lactic acid producerBacillus coagulanson starter culture in the low-salt Chinese potherb mustard pickle fermentation[J]. LWT-Food Science and Technology, 2008, 41: 474-482.
[5]張銳.低鹽榨菜自然發酵微生物群落動態的變化與功能分析[D].寧波:寧波大學, 2010.
[6]楊瑞,張偉,徐小會.泡菜發酵過程中主要化學成分變化規律的研究[J].食品工業科技, 2005, 26(2): 96-98.
[7] XIONG Tao, LI Xiao, GUAN Qianqian, et al. Starter culture fermentation of Chinese sauerkraut: growth, acidification and metabolic analyses[J]. Food Control, 2014, 41: 122-127.
[8]李文斌,宋敏麗,唐中偉,等.自然發酵泡菜微生物群落變化的研究[J].中國食物與營養, 2008(11): 22-24.
[9]李幼筠.泡菜與乳酸菌[J].中國釀造, 2001, 20(4): 7-9.
[10]尹曦.四川宜賓芽菜中乳酸菌菌相分析[D].雅安:四川農業大學, 2012.
[11] WOUTERS D, BERNAERT N, CONJAERTS W, et al. Species diversity, community dynamics, and metabolite kinetics of spontaneous leek fermentations[J]. Food Microbiology, 2013, 33: 185-196.
[12]天津輕工業學院,無錫輕工業學院.食品工藝學[M].北京:中國輕工業出版社, 1993: 66.
[13]閆征,王昌祿,顧曉波. pH值對乳酸菌生長和乳酸產量的影響[J].食品與發酵工業, 2002, 29(6): 35-38.
[14]翁佩芳,陳希,沈錫權,等.榨菜低鹽腌制細菌群落多樣性的分析[J].中國農業科學, 2012, 45(2): 338-345.
[15]楊瑞鵬,趙學慧.溫度與食鹽濃度對乳酸菌區系的影響[J].中國調味品, 1991, 16(8): 15-17.
[16]潘靜,王昌祿.多耐性酒精酵母菌的選育及特性研究[J].中國釀造, 2011, 30(5): 113-116.
[17]單藝,張蘭威.優選糯米酒腐木生假絲酵母菌Y-2生長性能及其釀造條件優化的研究[J].食品科技, 2007, 32(11): 40-45.
[18] TOKUOKA K,侯希君.耐糖和耐鹽的酵母菌[J].齊齊哈爾師范學院學報, 1993, 13(4): 39-43.
[19]衛永華,齊威,王春玲,等.耐鹽酵母在醬油中的應用及其耐鹽機理的研究[J].中國釀造, 2012, 31(4): 25-28.
[20]曾駿,陳安均,蒲彪,等.傳統四川泡菜中酵母菌的動態變化規律[J].食品科學, 2014, 35(7): 81-85. doi: 10.7506/spkx1002-6630-201407017.
[21] HYUN J E, DONG M S, NAM S H. Selecttion of psychrotrophicLeuconnstocspp. producing highly active dextransucrase from lactate fermented vegetables[J]. International Journal of Food Microniology, 2007, 117: 61-67.
[22] GULLO M, CAGGIA C, de VERO L, et al. Characterization of acetic acid bacteria in“traditional balsamic vinegar”[J]. International Journal of Food Microbiology, 2006, 106: 209-212.
[23] GOBBETTI M, CORSETTI A, ROSSI J. The sourdough microflora: interactions between lactic acid bacteria and yeasts: metabolism of carbohydrates[J]. Applied Microbiology and Biotechnology, 1994, 41(4): 456-460.
[24] XIONG Tao, GUAN Qianqian, SONG Shuhua, et al. Dynamic changes of lactic acid bacteria flora during Chinese sauerkraut fermentation[J]. Food Control, 2012, 26: 178-181.
[25] OLIVEIRA R P S, PEREGO P, OLIVEIRA M N, et al. Growth, organic acids profile and sugar metabolism ofBifidobacterium lactisin co-culture withStreptococcus thermophilus: the inulin effect[J]. Food Research International, 2012, 48: 21-27.
[26] GANZLE M G, VERMEULEN N, VOGEL R F. Carbohydrate, peptide and lipid metabolism of lactic acid bacteria in sourdough[J]. Food Microbiology, 2007, 24: 128-138.
[27] KANDLER O. Carbohydrate metabolism in lactic acid bacteria[J]. Antonie van Leeuwenhoek, 1983, 49: 209-224.
[28]郝明玉.直投式發酵泡菜與自然發酵泡菜的比較研究[D].南昌:南昌大學, 2013.
[29]唐愛明,夏廷斌.乳酸菌降解豬血培養基中亞硝酸鹽的研究[J].肉類工業, 2004(9): 34-36.
Effect of Salt Concentration on Microbial Community Composition and Metabolism in Traditional Pickled Cabbage
XIONG Tao, LI Junbo, PENG Fei, GUAN Qianqian (State Key Laboratory of Food Science and Technology, College of Food Engineering, Nanchang University, Nanchang 330047, China)
In this paper, traditional pickled cabbages obtained by natural fermentation with different addition amounts of salt (2, 5 and 8 g/100 mL) were studied by comparing the dominant microorganisms, pH, total acid, substrates and metabolites and nitrite in pickling juices during fermentation. The results indicated that lactic acid bacteria were the dominant bacteria during the fermentation process, sucrose was steadily consumed, glucose and fructose were gradually accumulated, and lactic acid was accumulated to a high level at the end of fermentation. Salinity had a significant effect on the early stage of fermentation, and 2 g/100 mL salt could result in the highest reproduction and metabolism rate of lactic acid bacteria, the lowest pH and the highest total acid at the end of fermentation. In contrast, 5 g/100 mL salt had obvious inhibitory effect on the growth of harmful microorganisms and resulted in the highest speed to reach the nitrite peak. The addition of 8 g/100 mL salt could result in the lowest reproduction and metabolism rate of lactic acid bacteria and the lowest utilization rate of sucrose.
pickled cabbage; salt concentration; lactic acid bacteria; fermentation; metabolism
TS255.54
1002-6630(2015)11-0172-05
10.7506/spkx1002-6630-201511033
2014-07-10
“贛鄱英才555工程”領軍人才培養計劃項目(18000063);國家重點實驗室自由探索課題(SKLF-ZZB-201309);國家高技術研究發展計劃(863計劃)項目(2011AA100904)
熊濤(1970—),男,教授,博士,研究方向為益生菌及大宗果蔬高值化利用。E-mail:xiongtao0907@163.com
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
