家蠅新型抗菌肽A3基因的克隆與分析
2015-01-22 07:13:22耿文一柳峰松
產(chǎn)業(yè)與科技論壇 2015年12期
關鍵詞:實驗
□ 耿文一 楊 雪 唐 婷 柳峰松
一、引言
隨著抗生素的濫用和超級耐藥細菌的產(chǎn)生,昆蟲抗菌肽已成為新型抗菌藥物設計和開發(fā)的重要資源。家蠅中已經(jīng)報道的抗菌肽有攻擊素(attacin),天蠶素(cecropin),防御素(defensin)和溶菌酶(lysozyme)[1]。抗菌肽的獲得可以通過許多方法,例如采用提取純化的方式獲得,但是提取工藝繁瑣;有研究是通過真/原核表達來表達純化抗菌肽[2],由于抗菌肽的抑菌能力較強,該方法的成功率較低;也有研究運用抑制性消減雜交方法成功獲得免疫相關基因[3]。本文通過分析家蠅基因數(shù)字表達譜和轉錄組數(shù)據(jù),發(fā)現(xiàn)細菌刺激后多條未知基因表達量顯著升高。分析其中1條基因編碼的多肽發(fā)現(xiàn)具有抗菌肽的典型特征,同時實驗證實根據(jù)其成熟肽合成的多肽具有廣譜抑菌活性。此基因是在家蠅中首次被發(fā)現(xiàn)的,并且在GenBank中找不到同源基因,將其命名為A3。
二、材料與試劑
(一)實驗材料。家蠅Musca domestica(Linnaeus)種蠅由中國科學院動物研究所何鳳琴老師惠贈,幼蟲由本實驗室飼養(yǎng),飼養(yǎng)溫度為25℃,相對濕度為50% ~70%。
(二)主要試劑。實驗室所用試劑是TaKaRa公司產(chǎn)品,有RNAiso Plus、瓊脂糖凝膠DNA回收試劑盒、pMD19-T載體、SYBR Green;南京金斯瑞科技有限公司產(chǎn)品有M-MLV反轉錄酶、Taq DNA聚合酶;蘇州金唯智生物有限公司合成引物。由上海默悉生物科技有限公司合成A3合成肽(純度大于95%)。
(三)主要菌種。攻毒實驗所用大腸桿菌(Escherichia coli ATCC25922)和金黃色葡萄球菌(Staphylococcus aureus)為本實驗室所保存。抑菌實驗所用菌種由中科院動物研究所朱順義研究員饋贈。革蘭氏陽性菌:巨大芽孢桿菌(Bacillus megaterium),枯草芽孢桿菌(Bacillus subtilis),藤黃微球菌(Micrococcus luteus),脲芽孢八疊菌(Sporosarcina ureae);革蘭氏陰性菌:嗜水汽單胞菌(Aeromonas hydrophila),根癌農桿菌(Agrobacterium tumefaciens),大腸桿菌(Escherichia coli),銅綠假單胞菌(Pseudomonas aeruginosa),鼠傷寒沙門氏菌(Salmonella typhimurium),黏質沙雷氏菌(Serratia marcescens)。
三、實驗方法
(一)總RNA的提取和反轉錄。按照使用說明書,用RNAiso Plus試劑提取家蠅幼蟲的總RNA,經(jīng)瓊脂糖凝膠電泳檢測其完整性后,利用核酸定量儀測定其濃度和純度,各取1μg 總 RNA,以 AOLP(5'-GGCCACGCGTCGACTAGTACT16(G/A/C)-3')為引物反轉錄合成cDNA。
(二)A3基因cDNA序列的克隆。參照家蠅轉錄組序列信息,設計A3基因特異正向引物A3-F1(5'-AACCACATTACGCTAACACG-3')和特異反向引物 A3-R1(5'-CATTAAATCGCGAGCCCTTTTTGA-3'),以家蠅幼蟲 cDNA為模板,進行 PCR 擴增,PCR 反應條件為:94℃、4min;94℃、30s,55℃、40s,72℃、1min,35 個循環(huán);72℃延伸,10min。PCR 產(chǎn)物經(jīng)1.0%瓊脂糖凝膠電泳分離、切膠回收后,連接pMD19-T載體,轉化大腸桿菌DH5α化學感受態(tài)細胞,PCR篩選陽性克隆后測序。
(三)A3抗菌肽的生物信息學分析。將上述測序結果進行生物信息學分析,利用Bioedit軟件搜索開放閱讀框(Open reading frame,ORF),翻譯成氨基酸序列。利用Signal 4.1 Server(http://www.cbs.dtu.dk/services/SignalP/)查找信號肽。成熟肽分子量、等電點及氨基酸組成在Antimicrobial Peptide Database(http://aps.unmc.edu/AP/)中預測。在NCBI網(wǎng)站上利用 Blastp程序(http://www.ncbi.nlm.nih.gov/blast)進行相似性比對。
(四)A3成熟肽合成。將A3成熟肽氨基酸序列VQRVLFPICTKGPFLHSLRDTDSLGRQLDPYVIR YQKKFGST發(fā)送到上海默悉生物科技有限公司進行合成。
(五)A3合成肽的抑菌活性實驗。將實驗菌株在37℃,220 rpm條件下震蕩過夜。將活化后的菌種以1%接種量接種到新鮮的液體培養(yǎng)基中,當OD600=0.2左右時,停止搖菌。將各菌株菌液用新鮮培養(yǎng)基稀釋100倍,備用。將A3合成肽用無菌水進行梯度稀釋,濃度分別是1 mM、200 μM、100 μM、20 μM、2 μM、0.2 μM、20 nM 和 2 nM。
我們對本實驗室的細菌菌譜進行大范圍的檢測。取其中一株細菌的稀釋菌液加入96孔板中,每孔加入菌液90 μl;多肽稀釋液以不同的濃度分別加入細菌稀釋液中,加入量為10 μl,23℃,100 rmp 震蕩培養(yǎng) 12 h。同時以分別加入 10 μl無菌水和10 μl氨芐青霉素(0.1 g/ml)作為陰性和陽性對照組。在OD600下檢測實驗組和對照組菌液的濁度,來判斷其是否具有抑菌活性,并測定最小抑菌濃度(minimal inhibitory concentration,MIC),MIC定義為與陰性對照組相比實驗組達到85%以上細菌死亡的多肽濃度。所有不同濃度的樣品要進行3次平行實驗。
(六)A3合成肽的溶血活性測定。抽取1 ml的兔靜脈血,立即與阿氏液(Alsever's Solution)混勻。用生理鹽水洗滌血細胞,2000 rmp,離心5 min,棄上清。用50倍體積生理鹽水重懸血細胞,并以200 μl每支分裝到1.5 ml離心管中。將多肽A3用生理鹽水稀釋成濃度梯度,分別為1 mM、204 μM、102 μM、20.4 μM、2.04 μM、0.204 μM、20.4 nM 和 2.04 nM。200 μl血細胞加入 4 μl肽液,37℃孵育30 min。3000 rmp,離心5 min,收集上清,在分光光度計上測定416 nm處吸光度值。陽性對照為用生理鹽水稀釋的10%Triton-X-100;陰性對照為生理鹽水。溶血率計算方法:溶血率(%)=[(AB)/(C-B)]×100%(A:實驗組吸光度值;B:陰性對照組吸光度值;C:陽性對照組吸光度值)。
四、結果
(一)家蠅A3基因cDNA序列及分析。經(jīng)PCR克隆,獲得家蠅A3基因cDNA序列254 bp,包含198 bp完整的開放閱讀框,5'末端非翻譯區(qū)12 bp,3'末端非翻譯區(qū)44 bp。推導的A3多肽序列由65個氨基酸殘基組成(圖1),圖中劃線部分為信號肽。成熟肽富含Leu(11.90%)和Arg(9.52%)(圖2),疏水性氨基酸為14個,帶正電荷的氨基酸為8個,理論分子量為4881.4 D,等電點為9.86。Blastp分析后未發(fā)現(xiàn)A3基因的同源性序列。
通過對抗菌肽二級結構分析,A3成熟肽是由β-折疊和α-螺旋構成(見圖3)。從三級結構預測圖中可以看出,在C端有1個α螺旋,N端有2個反向平行的β片層(見圖4)。

圖1 家蠅抗菌肽A3基因cDNA序列及其推導的氨基酸序列
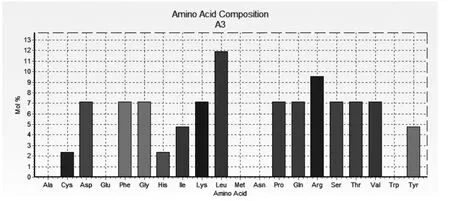
圖2 A3成熟肽的氨基酸組成
(二)A3合成肽的抑菌實驗結果。我們用實驗室已有的菌株來進行抑菌實驗。A3合成肽對多種細菌都有抑制作用,其中抑制作用最高的為革蘭氏陰性菌Serratia marcescens,其MIC 為30.73 μM。
(三)A3合成肽溶血實驗結果。從溶血實驗結果可以看出,合成肽的終濃度為1 mM時,其溶血效率為19.1%,說明A3對哺乳動物血細胞具有較低的溶血活性。
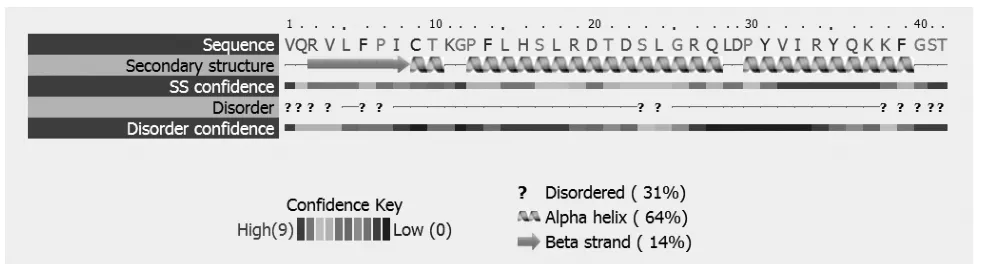
圖3 A3成熟肽的二級結構

圖4 A3成熟肽的三級結構
五、結語
本文根據(jù)家蠅EST序列篩選到一種新型的家蠅抗菌肽A3基因,合成A3成熟肽對革蘭氏陰性菌和革蘭氏陽性菌具有抑制作用,且?guī)缀鯖]有溶血活性,因此A3在醫(yī)藥領域具有一定應用潛力。本研究不但從家蠅體內發(fā)現(xiàn)了一種新型抗菌肽,在新抗菌肽的發(fā)現(xiàn)方法方面也有獨到之處,在基因組學和轉錄組學迅速發(fā)展的今天,利用生物信息學作為指導,結合分子生物學實驗驗證,獲得新基因是一種方便可行的方法,可突破生物化學方法分離蛋白的難關,會加快新型抗菌肽的研發(fā)進程。
[1]Boman HG,F(xiàn)aye I,Gudmundsson GH,Lee JY,Lidholm DA.Cell-free immunity in Cecropia.A model system for antibacterial proteins[J].Eur J Biochem,1991,201(1):23 ~ 31
[2]耿華,安春菊,郝友進,李德森,杜榮騫.家蠅攻擊素(Attacin)基因的克隆與表達[J]. 遺傳學報,2004,12:1344~1350
[3]Ito Y,Nakamura M,Hotani T,Imoto T.Insect lysozyme from house fly(Musca domestica)larvae:possible digestive function based on sequence and enzymatic properties[J].J Biochem,1995,118(3):546~551
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
