IL-31RA基因敲除小鼠純合子模型的建立*
2015-02-19 03:56:16江濤,高婧,岳歡等
貴州醫(yī)科大學(xué)學(xué)報(bào) 2015年3期
關(guān)鍵詞:小鼠
?
IL-31RA基因敲除小鼠純合子模型的建立*
網(wǎng)絡(luò)出版時(shí)間:2015-03-19網(wǎng)絡(luò)出版地址:http://www.cnki.net/kcms/detail/52.5012.R.20150319.0956.015.html
江濤, 高婧, 岳歡, 高雅倩, 黃俊瓊**
(遵義醫(yī)學(xué)院附屬醫(yī)院, 貴州 遵義563000)
[摘要]目的: 建立IL-31RA基因敲除小鼠純合子模型,為IL-31RA基因相關(guān)研究提供動(dòng)物模型。方法: IL-31RA基因敲除小鼠嚴(yán)格按照SPF級(jí)要求的動(dòng)物飼養(yǎng)標(biāo)準(zhǔn)進(jìn)行飼養(yǎng)繁殖,采用聚合酶鏈?zhǔn)椒磻?yīng)(PCR)法鑒定子代小鼠的基因型,RT-PCR法鑒定小鼠IL-31RA mRNA的表達(dá),Western blot鑒定IL-31RA蛋白的表達(dá),HE染色觀察小鼠重要臟器的形態(tài)學(xué)變化。結(jié)果: PCR法成功檢測(cè)出子代小鼠的3種基因型,純合子基因敲除小鼠未檢測(cè)出IL-31RA mRNA和IL-31RA蛋白的表達(dá),IL-31RA基因敲除小鼠的重要臟器的形態(tài)學(xué)特征與野生型小鼠比較無(wú)明顯變化;基因敲除小鼠可成功飼養(yǎng)繁殖,亦可獲得較多的基因敲除純合子小鼠。結(jié)論: 成功構(gòu)建了IL-31RA基因敲除小鼠純合子模型。
[關(guān)鍵詞]白細(xì)胞介素31; 基因敲除; 聚合酶鏈反應(yīng)(PCR); 反轉(zhuǎn)錄PCR; Western blot; 模型,動(dòng)物
白細(xì)胞介素31(Interleukin 31,IL-31)是2004年Dillon等發(fā)現(xiàn)的細(xì)胞因子,屬于白細(xì)胞介素6細(xì)胞因子家族成員,主要由活化的CD4+Th2細(xì)胞產(chǎn)生。IL-31受體(IL-31R)是由IL-31RA與制瘤素M受體B(oncostatin M receptor B, OSMRB)組成的異源二聚體,在皮膚及支氣管上皮組織中表達(dá)水平較高。國(guó)內(nèi)外研究發(fā)現(xiàn)IL-31及其受體與皮膚損傷、禿發(fā)癥、搔癢、特應(yīng)性皮炎、氣道超敏反應(yīng)及炎癥性腸病等疾病相關(guān),本課題組前期研究發(fā)現(xiàn)IL-31及其受體在過(guò)敏性哮喘小鼠肺組織中表達(dá)增高,IL-31可誘導(dǎo)肺組織分泌趨化因子。為進(jìn)一步研究IL-31在過(guò)敏性哮喘中的作用,本研究成功構(gòu)建并鑒定出IL-31RA基因敲除小鼠純合子模型,報(bào)告如下。
1材料與方法
1.1 材料
1.1.1實(shí)驗(yàn)動(dòng)物IL-31RA基因敲除小鼠品系為C57BL/6J,4周齡,6雄2雌,購(gòu)自突變小鼠區(qū)域資源中心(MMRRC),在遵義醫(yī)學(xué)院醫(yī)學(xué)與生物學(xué)研究中心繁殖擴(kuò)群。
1.1.2主要試劑Premix Taq(TaKaRa TaqTMVersion 2.0 plus dye)(TaKaRa公司),DL 500 Marker,GoldView,動(dòng)物組織基因組DNA提取試劑盒(上海生工),總RNA提取試劑盒(天根生物),引物由上海生工合成,DEPC(Sigma公司),兔抗鼠IL-31RA抗體、HRP標(biāo)記羊抗兔IgG抗體(北京博奧森公司) , 化學(xué)發(fā)光試劑及硝酸纖維素膜(GE公司)。
1.2 方法
1.2.1基因敲除小鼠雜合子模型由MMRRC進(jìn)行設(shè)計(jì)及完成。基因敲除過(guò)程簡(jiǎn)述如下:IL-31RA基因組克隆來(lái)源于129SvEvBrd基因組文庫(kù),構(gòu)建一個(gè)替換型載體替換IL-31RA的第4外顯子,即IL-31RA基因組序列(MGI:2180511)中8093-8172堿基,共80 bp;采用電穿孔法將此打靶載體轉(zhuǎn)入C57BL/6J小鼠胚胎干細(xì)胞(ES細(xì)胞),經(jīng)G418 篩選得到陽(yáng)性克隆的ES細(xì)胞,并將此ES細(xì)胞植入假孕的C57BL/6J小鼠子宮內(nèi),發(fā)育得到雜合子的IL-31RA基因敲除小鼠。所有小鼠均飼養(yǎng)在遵義醫(yī)學(xué)院醫(yī)學(xué)與生物學(xué)研究中心的屏障系統(tǒng)(SPF)小鼠飼養(yǎng)室,飼料購(gòu)自重慶第三軍醫(yī)大學(xué)生產(chǎn)的SPF級(jí)小鼠飼料【生產(chǎn)許可證號(hào)SCXK(渝)2012-0004】,飲用水為消毒純凈水,小鼠所接觸物品均高壓預(yù)真空滅菌。小鼠的性成熟期約35 d,妊娠期約20 d,采用雄雌鼠各1只合籠繁殖。
1.2.2基因敲除小鼠的鑒定因引進(jìn)的IL-31RA基因敲除小鼠均為雜合子,繁殖出的子代可出現(xiàn)野生型(Wild-type,WT)IL-31RA+/+、雜合子IL-31RA+/-及突變型(Knockout,KO)IL-31RA-/-3種表型,對(duì)子代IL-31RA基因敲除小鼠作為模型需先鑒定其基因型。IL-31RA基因敲除小鼠基因組DNA的提取:剪取小鼠尾巴末端約0.6 cm,按說(shuō)明書(shū)操作抽提DNA,雙蒸水溶解, 4 ℃保存。聚合酶鏈?zhǔn)椒磻?yīng)(PCR)及瓊脂糖凝膠電泳鑒定IL-31RA基因敲除小鼠子代基因型:按照參考文獻(xiàn)設(shè)計(jì)引物,WT上游系列為5′-GGGTGTGAACGCTGGAATAATG-3′、KO上游系列為5′-GCAGCGCATCGCCTTCTATC-3′(含有插入的neo基因序列,可鑒定插入的載體片段)、共用下游引物系列為5′-GATCATCTCTCCAAATTTACATG-3′。WT型目的片段長(zhǎng)274 bp,KO型目的片段長(zhǎng)348 bp。反應(yīng)體系(25 μL):Premix Taq 12.5 μL, WT和KO上游及下游引物各1 μL,模板DNA 2 μL,加滅菌蒸餾水補(bǔ)足體積。循環(huán)條件為95 ℃預(yù)變性3 min;95 ℃變性30 s、60 ℃退火 30 s, 30個(gè)循環(huán),最后72 ℃延伸10 min。4 ℃保存?zhèn)溆谩CR 反應(yīng)終產(chǎn)物5 μL進(jìn)行2%瓊脂糖凝膠電泳鑒定IL-31RA基因敲除小鼠子代基因型,判斷標(biāo)準(zhǔn)為WT型和KO型分別在274 bp與348 bp處出現(xiàn)單一的目的條帶,雜合子則在274 bp和348 bp處同時(shí)出現(xiàn)目的條帶。
1.2.3IL-31RA基因敲除小鼠IL-31RA mRNA的鑒定采用RT-PCR法,IL-31RA基因敲除小鼠總RNA的提取及cDNA的合成:取已經(jīng)鑒定的WT及KO小鼠肺組織,按照總RNA提取試劑盒操作說(shuō)明書(shū)抽提RNA,逆轉(zhuǎn)錄合成cDNA。逆轉(zhuǎn)錄體系(20 μL):5×Prime ScriptTMBuffer 4 μL,Prime Script TM RT Enzyme Mix I 1 μL,Oligo dT Primer(50 μM) 1 μL, Random 6 mers(100 μM) 1 μL,Total RNA 10 μL,RNase Free dH2O補(bǔ)足體積。逆轉(zhuǎn)錄反應(yīng)條件:37 ℃,15 min;85 ℃,5 s; cDNA放于-20 ℃保存。PCR反應(yīng)及瓊脂糖凝膠電泳鑒定mRNA,設(shè)計(jì)及合成含有IL-31RA基因第4號(hào)外顯子的基因序列引物,上游系列為5′-TCATTTCTCAGCGTCAGTGG-3′、下游系列為5′-TCCTTCTCTGGTCTCCAAGTG-3′;擴(kuò)增的目的基因片段長(zhǎng)為389 bp。 反應(yīng)體系(25 μL):Premix Taq 12.5 μL,上、下游引物各1 μL,模板cDNA 2 μL,滅菌蒸餾水補(bǔ)足體積。反應(yīng)條件: 94 ℃預(yù)變性2 min; 94 ℃變性30 s、61 ℃退火30 s、72 ℃延伸30 s, 30個(gè)循環(huán);最后72 ℃延伸10 min。4 ℃保存?zhèn)溆谩HCR反應(yīng)終產(chǎn)物5 μL進(jìn)行2%瓊脂糖凝膠電泳鑒定小鼠IL-31RA mRNA。
1.2.4IL-31RA基因敲除小鼠IL-31RA蛋白的鑒定采用Western Blot法,提取已經(jīng)鑒定的KO及WT小鼠肺組織蛋白質(zhì),蛋白定量后以4∶1比例與Buffer混合,100 ℃水浴6 min,常規(guī)行SDS-PAGE電泳后轉(zhuǎn)印至硝酸纖維素膜,用封閉液 (含50 g/L脫脂奶粉的TBST)室溫振搖封閉2 h。依次加入兔抗鼠IL-31RA抗體及 HRP標(biāo)記羊抗兔IgG,化學(xué)發(fā)光法曝光顯色, 觀察結(jié)果。
1.2.5IL-31RA基因敲除對(duì)小鼠重要臟器的影響臟器系數(shù)測(cè)定:頸椎脫臼法處死小鼠,稱(chēng)量小鼠體重及臟器(心臟、肝臟、腎臟、肺、胸腺及脾臟)的重量,臟器系數(shù)=100×臟器重量(g)/體重(g)。HE染色:心臟、肝臟、腎臟、肺、胸腺及脾臟于4%多聚甲醛中固定48 h后,常規(guī)石蠟切片,HE染色。在顯微鏡下觀察各個(gè)臟器的形態(tài)學(xué)變化。
1.3 統(tǒng)計(jì)學(xué)處理
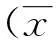
2結(jié)果
2.1 IL-31RA基因敲除小鼠的生長(zhǎng)繁殖情況
IL-31RA基因敲除小鼠(IL-31RA-/-)能成功繁殖(已繁殖5代),子代小鼠生長(zhǎng)狀況良好(已觀察1年零6個(gè)月)。小鼠約35 d性成熟,性周期為4~5 d,妊娠期約20 d,哺乳期約20 d,每胎產(chǎn)仔數(shù)為7~12只,成活率>98%。
2.2 IL-31RA基因敲除小鼠基因型鑒定
小鼠基因型的鑒定結(jié)果如圖1,其中1、2號(hào)小鼠為野生型(IL-31RA+/+),僅出現(xiàn)274 bp條帶;3號(hào)小鼠為雜合子(IL-31RA+/-),同時(shí)出現(xiàn)274 bp 和348 bp 2個(gè)條帶,4、5號(hào)小鼠為突變型(IL-31RA-/-),僅出現(xiàn)1個(gè)348 bp條帶。

圖1 IL-31RA基因敲除小鼠基因型鑒定Fig.1 Identification results of genotype of IL-31RA gene knockout mice
2.3 IL-31RA基因敲除小鼠IL-31RA mRNA的鑒定
RT-PCR結(jié)果示W(wǎng)T小鼠(1、2號(hào))檢測(cè)出389 bp處的條帶,而KO小鼠(3、4、5號(hào))未檢測(cè)出。見(jiàn)圖2。

圖2 小鼠IL-31RA mRNA的RT-PCR鑒定Fig.2 Identification results of IL-31RA mRNA of the mice
2.4 IL-31RA基因敲除小鼠IL-31RA蛋白鑒定
Western Blot結(jié)果示IL-31RA基因敲除小鼠IL-31RA蛋白的相對(duì)分子質(zhì)量為83 kDa,WT小鼠檢測(cè)到相對(duì)分子量約為83 kDa片段,KO小鼠未檢出。見(jiàn)圖3。

圖3 小鼠IL-31RA蛋白的Western Blot鑒定Fig.3 Identification results of IL-31RA protein of the mice
2.5 L-31RA基因敲除小鼠重要臟器系數(shù)及HE染色
純合子IL-31RA基因敲除小鼠與野生型間各臟器的臟器系數(shù)比較,差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05),見(jiàn)表1;兩者間形態(tài)學(xué)也無(wú)明顯改變,見(jiàn)圖4。

表1 L-31RA基因敲除小鼠心臟、肝臟、腎臟、

圖4 小鼠各臟器的HE染色結(jié)果(×200)Fig.4 HE staining results of each organ of mice
3討論
IL-31的功能包括調(diào)節(jié)細(xì)胞增殖分化,調(diào)節(jié)造血和免疫反應(yīng),誘導(dǎo)炎性細(xì)胞因子和趨化因子分泌,參與調(diào)節(jié)炎癥反應(yīng),是重要的炎癥因子。IL-31可誘導(dǎo)皮膚炎癥,刺激表皮角質(zhì)細(xì)胞分泌趨化因子、巨噬細(xì)胞炎性蛋白3β、胸腺活化調(diào)節(jié)的趨化因子、T細(xì)胞活化蛋白-3[4-6]。Lei Z 等發(fā)現(xiàn)過(guò)敏性哮喘患者血清IL-31和外周血單核細(xì)胞IL-31mRNA均明顯高于正常對(duì)照組,Shah SA等研究發(fā)現(xiàn)IL-31可上調(diào)氣道上皮細(xì)胞黏液素基因MUC5AC的表達(dá);IP WK等采用IL-31在體外刺激人支氣管上皮細(xì)胞,通過(guò)熒光定量PCR檢測(cè)發(fā)現(xiàn)單核細(xì)胞趨化蛋白-1(MCP-1)、血管內(nèi)皮生長(zhǎng)因子(VEGF)、上皮生長(zhǎng)因子(EGF)表達(dá)增高;Stott B 等[10]發(fā)現(xiàn)IL-31可由IL-4誘導(dǎo)表達(dá),可以促進(jìn)Th2型炎癥反應(yīng),誘導(dǎo)促炎基因的表達(dá)。Liu W等[11]研究發(fā)現(xiàn)兒童過(guò)敏性鼻炎(AR)伴有哮喘患者血清和鼻灌洗液中IL-31mRNA和IL-31比正常對(duì)照組及單純患AR組顯著增高,并且增高趨勢(shì)與Th2細(xì)胞因子及黏蛋白5AC(MUC5AC)的表達(dá)一致。近年,越來(lái)越多的國(guó)內(nèi)外研究證實(shí)IL-31在氣道過(guò)敏性疾病中發(fā)揮重要作用,有望成為該疾病的診斷和(或)監(jiān)測(cè)指標(biāo)。
IL-31在過(guò)敏性哮喘中發(fā)揮重要作用,但其具體機(jī)制尚不清楚。基因敲除小鼠技術(shù)是建立在同源重組理論基礎(chǔ)上的一種技術(shù),借助細(xì)胞生物學(xué)、分子生物學(xué)、和動(dòng)物胚胎學(xué)的方法,使小鼠體內(nèi)某種基因的功能缺失,是闡明該基因功能最直接的手段之一,因此基因敲除技術(shù)成為后基因組時(shí)代最重要的研究方法和內(nèi)容[12]。本研究采用同源重組的方法敲除小鼠IL-31RA基因包含在編碼信號(hào)肽的基因序列內(nèi)外顯子4,使IL-31RA失去功能,通過(guò)觀察IL-31RA基因敲除小鼠生長(zhǎng)繁殖情況,發(fā)現(xiàn)IL-31RA基因敲除小鼠與野生型小鼠具有一樣的生長(zhǎng)繁殖能力,并且IL-31RA基因敲除對(duì)小鼠的重要臟器沒(méi)有明顯影響。雜合子小鼠交配后子代可能出現(xiàn)WT、KO及雜合子3種基因型,故必須對(duì)它們進(jìn)行基因型鑒定。本研究采用PCR法對(duì)4周齡基因敲除小鼠進(jìn)行基因型篩選,進(jìn)一步采用RT-PCR法及Western Blot法對(duì)篩選出的WT、KO小鼠進(jìn)行IL-31RA mRNA及IL-31RA蛋白的表達(dá)進(jìn)行鑒定,結(jié)果證明成功構(gòu)建了IL-31RA基因敲除小鼠的純合子模型,該模型的成功構(gòu)建,為更好的探討IL-31-IL-31R信號(hào)在過(guò)敏性哮喘中的作用提供了一個(gè)很好的動(dòng)物模型。
參考文獻(xiàn)4
[1]Stacey RD, Cindy S, Angela H, et al. Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat Immunol, 2004(7):752-760.
[2]杜清波,黃俊瓊.IL-31在哮喘小鼠肺組織中的表達(dá)及對(duì)肺組織細(xì)胞ck的影響.遵義:遵義醫(yī)學(xué)院, 2012.
[3]Bilsborough J, Mudri S, Chadwick E, et al. IL-31 receptor (IL-31RA) knockout mice exhibit elevated responsiveness to oncostatinM. Immunol, 2010(10):6023-6030.
[4]Raap U, Weiβmantel S, Gehring M, et al. IL-31 significantly cor-relates with disease activity and Th2 cytokine levels in children with atopic dermatitis. Pediatr Allergy Immunol, 2012(3):285-288.
[5]Nobbe S, Dziunycz P, Mühleisen B, et al. IL-31 expression by inflammatory cells is preferentially elevated in atopic dermatitis. Acta Derm Venereol, 2012(1):24-28.
[6]黃俊瓊,謝宇鋒,盛偉華,等. IL-31對(duì)人表皮角化細(xì)胞的影響. 中國(guó)免疫學(xué)雜志, 2007(4):369-373.
[7]Lei Z, Liu G, Huang Q, et al. SCF and IL-31 rather than IL-17 and BAFF are potential indicators in patients with allergic asthma. Allergy, 2008(63):327-332.
[8]Shah SA, Ishinaga H, Hou B, et al. Effects of interleukin-31 on MUC5AC gene expression in nasal allergic inflammation. Pharmacology, 2013(3-4):158-164.
[9]Ip WK, Wong CK, Li ML, et al. Interleukin-31 induces cytokine and chemokine production from human bronchial epithelial cells through activation of mitogen-activated protein kinase signalling pathways: implications for the allergic response. Immunology, 2007(122):532-541.
[10]Stott B, Lavender P, Lehmann S, et al. Human IL-31 is induced by IL-4 and promotes TH2-driven inflammation. Allergy Clin Immunol, 2013(2):446-454.
[11]Liu W, Luo R, Chen Y, et al. Interleukin-31 promotes helper T cell type-2 inflammation in children with allergic rhinitis. Pediatr Res, 2014(6):1038-1051.
[12]Guan C, Ye C, Yang X, et al. A review of current large-scale mouse knockout efforts. Genesis, 2010(48):73-85.
(2015-01-08收稿,2015-02-27修回)
中文編輯: 吳昌學(xué); 英文編輯: 周凌
·基礎(chǔ)研究·
Establishment of IL-31RA Gene Knockout Homozygous Mice Model
JIANG Tao, GAO Jing, YUE Huan, GAO Yaqian, HUANG Junqiong
(TheAffiliatedHospitalofZunyiMedicalCollege,Zunyi563000,Guizhou,China)
[Abstract]Objective: To establish the IL-31RA gene knockout homozygous mice model and lay the foundation for further study on IL-31 gene. Methods: IL-31RA gene knockout mice were bred and reproduced according to the SPF class animal feeding standard. PCR was used to identify the genotype of the offspring, the expression of IL-31RA mRNA was detected by RT-PCR, expression of IL-31RA protein was detected by Western blot, and morphological changes of vital organs were observed by HE staining. Result: Three genotypes of the offspring of IL-31RA gene knockout mice were successfully identified; expression of IL-31RA mRNA and IL-31RA protein was not detected in IL-31RA gene knockout homozygous mice. Compared with the wild type mice, morphological characteristics of vital organs of had no significant changes in IL-31RA gene knockout homozygous mice. IL-31RA gene knockout mice could be dred and reproduced successfully. Conclusion: The IL-31RA gene knockout homozygous mice model has been successfully established.
[Key words]interleukin 31; gene knockout; polymerase chain reaction; reverse transcription-PCR; Western blot; model, animal
[中圖分類(lèi)號(hào)]R-332
[文獻(xiàn)標(biāo)識(shí)碼]A
[文章編號(hào)]1000-2707(2015)03-0229-05
通信作者**E-mail:junqiongh@aliyun.com
[基金項(xiàng)目]*國(guó)家自然科學(xué)基金資助項(xiàng)目(No:81260266); 貴州省優(yōu)秀科技教育人才省長(zhǎng)專(zhuān)項(xiàng) (No:黔省專(zhuān)合字200951)
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫(huà)刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
 貴州醫(yī)科大學(xué)學(xué)報(bào)2015年3期
貴州醫(yī)科大學(xué)學(xué)報(bào)2015年3期
- 貴州醫(yī)科大學(xué)學(xué)報(bào)的其它文章
- 醫(yī)學(xué)院校不同專(zhuān)業(yè)畢業(yè)生醫(yī)學(xué)人文素質(zhì)調(diào)查分析*
- 藥物結(jié)合心理干預(yù)對(duì)高血壓患者血壓、抑郁情緒和生活滿(mǎn)意度的影響*
- 成分輸血培訓(xùn)教育對(duì)血漿臨床合理使用率的影響
- 真皮下血管網(wǎng)皮瓣成活質(zhì)量的實(shí)驗(yàn)研究
- 子宮內(nèi)膜異位癥與TNF-β G252A 基因的相關(guān)性研究
- 貴陽(yáng)市農(nóng)村初中學(xué)生膳食營(yíng)養(yǎng)狀況調(diào)查*