鎘脅迫下鋅對水稻植物螯合肽合成的影響
2015-03-07 06:42:28于莎莎卞英芳孫偉華曹趙云
浙江農業科學 2015年11期
于莎莎,卞英芳,楊 歡,孫偉華,曹趙云
(中國水稻研究所,農業部稻米及制品質量監督檢驗測試中心,浙江杭州 311400)
鎘脅迫下鋅對水稻植物螯合肽合成的影響
于莎莎,卞英芳,楊 歡,孫偉華,曹趙云
(中國水稻研究所,農業部稻米及制品質量監督檢驗測試中心,浙江杭州 311400)
以水稻品種9311為試驗材料,采用水培試驗研究了鎘(Cd)離子脅迫下水稻幼苗根部和地上部生物量、重金屬含量、植物螯合肽(PCs)及谷胱甘肽(GSH)含量的變化,及添加鋅(Zn)離子對Cd脅迫作用的調節。結果顯示,Cd脅迫降低了水稻生物量,下降幅度達27.6%~37.0%;隨Cd處理濃度的增加,水稻根部和地上部PCs含量分別增加了93.7%和84.7%;Zn可以降低Cd脅迫水稻根部PCs和GSH的合成水平以及Cd的含量。一定濃度范圍內,Cd?Zn高量表達PCs,PCs與重金屬Cd結合,緩解了重金屬Cd的毒害。
鎘;鋅;水稻;植物螯合肽;谷胱甘肽
隨著工礦業的發展、農業污水灌溉、農藥以及化肥的施用,農田土壤鎘(Cd)污染日趨嚴重[1]。水稻是我國主要的糧食作物,也是一種極容易積累鎘的作物,受Cd污染嚴重[2]。
Cd對水稻的毒害是多方面的,相應地,水稻會采取多方面的策略來抵抗或減弱Cd脅迫的程度,如固定化、區域化及合成植物螯合肽、金屬硫蛋白等[3]。絡合是植物細胞解除Cd毒害的主要方式,它主要通過生物體內有生物活性的配位體與金屬離子發生專性較強的生物化學反應,形成螯合物從而緩解重金屬的毒害。植物螯合肽(PCs)是在外界重金屬誘導脅迫下,細胞質中合成的一類低分子量、富含半胱氨酸的巰基多肽化合物,以GSH為合成前體[4],基本的結構式為(γ?Glu?Cys)n?Gl y,(n=2~11,植物中常為2~4),對重金屬離子的螯合能力強,在重金屬的累積和解毒過程中發揮重要作用。研究表明,植物螯合肽的生物合成及解毒機制在重金屬環境治理污染中具有良好的應用前景[5-6]。
植物在天然環境中具有合成PCs的潛能,但在修復重金屬污染過程中需要通過修飾或高量表達植物螯合肽合成酶獲得高產PCs,或者借助重金屬的激活作用促使植物螯合肽合成酶催化合成PCs[7]。研究表明,Cd是激活植物螯合肽合成酶最有效的金屬[8],為了緩解有毒重金屬的毒害作用,最有效的方式是通過Cd的激活誘導合成足量的PCs,但Cd通常是有毒的,如何解決上述矛盾已引起廣泛的關注。鋅(Zn)與Cd具有相同的核外電子構型,化學性質相似,但Zn是植物生長必需的微量元素之一,在體內發揮重要的生理作用,被稱為人體的智慧元素[9]。雖然它們對植物的作用相反,但二者之間存在復雜的交互影響關系。目前Cd?Zn之間相互作用主要有協同作用、拮抗作用兩種觀點。He等[10]的田間試驗表明施Zn使萵苣中Cd含量顯著升高,二者之間為協同作用。Bunluesin等[11]曾經報導了高Zn/Cd比例有助于降低Cd的積累,同樣Cd的存在也導致Zn含量的降低。Hart等[12]認為Zn的施用抑制了小麥對Cd的吸收,從而減少了植株各部位Cd的濃度,可能是Zn能誘導PCs的產生,PCs降低了Cd的有效性,從而降低了Cd的毒害。Cd,Zn交互作用對植物的影響是復雜的,能否通過二者的交互作用高量表達PCs從而緩解Cd的毒害尚未有定論。
目前,大田水稻受Cd污染后的評價研究較多,通常以籽粒中的含Cd量作為主要的指標[13],但是對水稻苗期受Cd污染的評價研究較少。本研究采用水培試驗,探討Zn/Cd比例對水稻幼苗的生物量、重金屬含量、PCs及GSH含量的影響,為水稻Cd,Zn競爭吸收機理提供理論依據,也為利用Zn緩解Cd污染提供理論指導。
1 材料與方法
1.1 材料與試劑
供試水稻品種為9311。
乙腈、甲醇(色譜純,德國Merck公司)、甲酸(色譜純)、GSH(≥97%)、DTT(≥99%)均購自美國Sigma?Aldrich公司;PC2,PC3和PC4均購自美國AnaSpec Inc.公司,純度均≥95%;Bond Elut Plexa PAX(60 mg×3 mL)購自Agilent Technologies公司;試驗用水為Milli?Q超純水。
1.2 儀器與設備
Survryor系列液相色譜儀,TSQ Quantum Access Max三重四極桿質譜儀,X?series 2電感耦合等離子體質譜儀(ICP?MS,美國ThermoFisher公司),MS3 basic漩渦振蕩儀(德國IKA公司),KQ?800TED型超聲波清洗器和臺式冷凍離心機Biofuge Primo R(美國Thermo Fisher公司)。
1.3 處理設計
水稻種子于35℃黑暗條件下催芽,露白后播種于沙床,于光照培養箱(白天35℃,夜晚25℃,相對濕度80%)中培養。待水稻長至3葉1心期,選取健壯、長勢一致的水稻幼苗轉移至容積為6 L、內裝有5 L培養液的塑料黑桶中,每桶5穴,每穴3株,以樹脂塑料板分隔各穴,海綿固定幼苗。營養液每3 d換1次,調節pH值為5.0~5.3。水培1周后進行單金屬以及復合金屬處理。各處理編號依次為CK,A,B,C,D,E,F,G,H,I,其中Cd添加濃度依次為0,1,10,50,1,1,1,50,50,50μmol·L-1,Zn添加濃度依次為0,0,0,0,1,10,50,1,10,50μmol·L-1。
1.4 測定指標
1.4.1 生物量的測定
處理10 d后收獲水稻幼苗,先用自來水沖洗根表面吸附的雜質,再用去離子水沖洗2遍,用吸水紙將其表面的水分吸干,分為地上部和根部,置于恒溫干燥箱中,105℃殺青0.5 h,85℃烘干至恒重,稱地上部和根部干物質的質量。
1.4.2 重金屬含量的測定
參照許萍等[14]的方法,分別稱取地上部和根部0.25 g試樣(精確至0.001 g),置于50mL聚丙烯消解管,加入8mL硝酸,于石墨消解爐中65℃緩慢加熱使樣品與酸完全混合,加熱30 min后,將消解儀溫度調至120℃,繼續消解至消化液約1 mL,取出消解管,加入3 mL雙氧水,繼續消解至消化液1 mL,取出消解管,冷卻后,用高純水定容至50.0 mL,混勻。用ICP?MS分析Cd和Zn的含量。
1.4.3 PCs等巰基化合物的提取和測定
參照Cao等[15]的方法,取水稻鮮樣放入研缽中,加入液氮研磨至粉末狀,稱取0.2 g(精確至0.001 g)于15 m l塑料離心管中,加入2 mL 4℃預冷的水(含100 mmol·L-1 dithiothreitol),渦旋振蕩1 min,超聲波破碎5 min,4℃8 000 r· min-1離心10 min,收集上清液,過0.22μm濾膜后,采用液質聯用儀進行分析測定。
1.5 數據統計
試驗數據用Excel進行統計,采用SPSS統計分析軟件進行方差分析(P<0.05)。
2 結果與分析
2.1 Cd脅迫對水稻生長的影響
由圖1可知,根部和地上部的生物量隨著Cd處理水平的提高而減少,在50μmol·L-1 Cd脅迫下,水稻幼苗根系和地上部生長明顯受到抑制,干質量明顯偏低,分別為對照(CK)的64.5%和61.7%。在低Cd處理下,隨著Zn添加濃度的增加,水稻根部和地上部的生物量均表現出先升高后下降的趨勢,其中尤其以添加10μmol·L-1 Zn效果最好,根部和地上部干質量明顯高于單獨Cd處理,分別為單獨Cd處理的1.07和1.12倍。在高Cd處理下,添加1μmol·L-1Zn可促進水稻根部和地上部生物量的增加,分別比單獨Cd處理增加了13%和15%;與單獨Cd處理相比,添加10和50μmol·L-1 Zn,根部和地上部生物量顯著下降。表明Cd處理對水稻幼苗產生了一定的生理毒害,Zn在一定程度上能夠緩解Cd的毒害,但緩解作用有限。
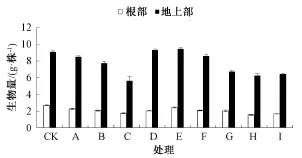
圖1 不同Cd離子濃度及Zn/Cd比例對水稻幼苗生物量的影響
2.2 水稻體內Cd和Zn的含量
由表1可知,在重金屬Cd脅迫下,水稻根部和地上部Cd的積累隨著Cd處理濃度的增加而增大,即Cd處理濃度為1,10和50μmol·L-1時,根系Cd含量分別為對照的236.7,511.6和679.8倍,地上部Cd含量分別是對照的36.9,68.3和122.4倍。低濃度Cd處理下,Zn的添加顯著地降低了水稻根部和地上部Cd的含量,且隨Zn添加濃度的增加,Cd含量在水稻根部和地上部的水平顯著降低,當Zn濃度為50μmol·L-1時,根部Cd含量為22.97mg·kg-1,與未添加Zn相比,Cd含量降低了35.3%,同時地上部分Cd含量降低了14.2%。在高濃度Cd處理下,Zn濃度的增加導致水稻根部和地上部Cd含量的協同增大,根部Cd含量由108.55 mg·kg-1增加到113.71 mg·kg-1,地上部Cd含量由15.67 mg·kg-1增加到23.79 mg·kg-1。不同濃度的Cd也會影響水稻根部和地上部中Zn的積累。結果表明,低濃度的Cd處理有利于Zn的吸收;高濃度Cd處理則抑制Zn的積累。當Zn添加濃度為50μmol·L-1時,1μmol·L-1 Cd脅迫處理的水稻根部中Zn含量為69.04 mg·kg-1,地上部分為36.77 mg·kg-1;當Zn添加濃度為50μmol·L-1,Cd處理濃度為50μmol·L-1時,根部和地上部分的Zn含量分別為46.22和18.11 mg·kg-1。表明一定范圍內二者在重金屬吸收水平上表現出一定的互作效應,在低Cd脅迫處理時,Zn對Cd的吸收表現出拮抗效應,高Cd脅迫處理時表現出協同效應;而Cd對Zn的吸收表現出協同效應;超過一定范圍結果相反。
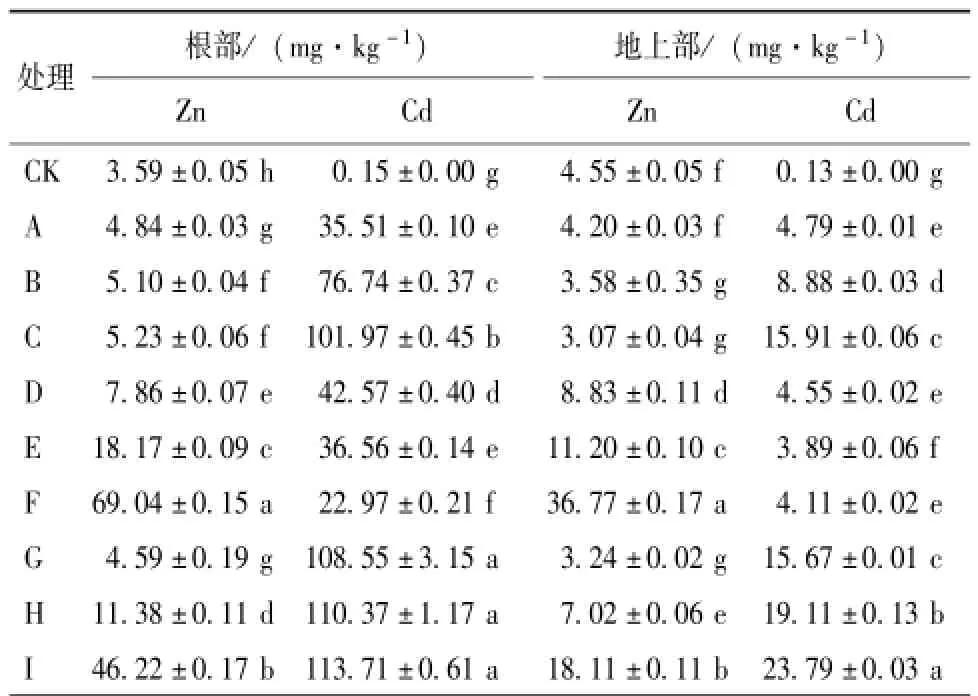
表1 不同濃度的Cd及Cd+Zn處理下水稻根部和地上部重金屬的含量
2.3 水稻幼苗植物螯合肽和GSH的含量
2.3.1 Cd脅迫下Zn對水稻幼苗中PCs含量的影響
由圖2中A可以看出,隨著Cd處理水平的提高,水稻根部PCs總量逐漸增大,由94.33 mg· kg-1增大到256.96 mg·kg-1,其中PC2與PC3增加的尤為顯著,分別比對照增加了15.3,15.9倍。與單獨Cd處理相比,Zn的添加降低了水稻根部PCs含量,并隨Zn添加濃度的增加降低作用更顯著,其中,低濃度Cd處理中隨Zn添加濃度的增加,水稻根部PCs含量從115.82 mg·kg-1降低到58.26 mg·kg-1;高濃度Cd處理中隨Zn添加濃度的增加,水稻根部PCs含量從323.43 mg·kg-1降低到226.15 mg·kg-1。低Cd處理時,隨Zn添加濃度的增大,PC3和PC4含量顯著降低,分別降低了58.28%和80.21%。高Cd處理時,隨Zn添加濃度的增大,PC2和PC3含量顯著降低,分別降低了13.95%和33.43%。
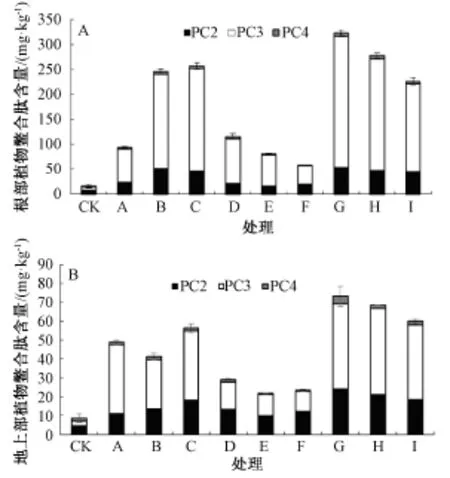
圖2 不同Cd離子濃度及Zn/Cd比例對水稻根部和地上部植物螯合肽含量的影響
由圖2中B可知,隨著Cd處理水平的提高,水稻地上部PCs總量呈先降低后升高的趨勢,當Cd處理濃度為10μmol·L-1時,降低尤為明顯。低濃度Cd處理中Zn的添加使水稻地上部PCs含量與單獨Cd處理相比降低了40.3%~55.6%;高濃度Cd處理中Zn的添加降低了水稻地上部PCs含量,由73.05 mg·kg-1降低到59.88 mg·kg-1,但與單獨Cd處理相比,增加了3.52 mg·kg-1。低Cd處理時,隨Zn添加濃度的增大,PC3和PC4含量顯著降低,分別降低了25.58%和71.94%。高Cd處理時,隨Zn添加濃度的增大,PC2,PC3和PC4含量顯著降低,分別降低了23.16%,12.14%和54.81%。
由圖2可以看出,在Cd脅迫下,地上部PCs的含量低于根部;在Cd脅迫下地上部和根部PC3含量高于其他巰基多肽的含量。以上結果表明,Zn的添加使得Cd與Zn在吸收上出現競爭效應,Zn降低了Cd的吸收,使得水稻根部和地上部PCs的含量降低,表現出抑制效應。
2.3.2 Cd脅迫下Zn對水稻幼苗谷胱甘肽含量的影響
GSH與PCs具有相似的結構,是PCs合成的前體[4],具有很強的氧化還原特性,可有效地清除活性氧及自由基,在植物抗逆境脅迫中起重要作用。由圖3可知,在Cd脅迫下,水稻幼苗地上部GSH含量顯著高于根部,當Cd脅迫濃度為50μmol· L-1時,地上部GSH含量是根部的5倍;在對照中,水稻幼苗根部和地上部中的GSH含量較高,分別為121.52和202.63 mg·kg-1;水稻幼苗根系在單獨Cd脅迫下GSH含量一直低于對照值,最低下降了84.5%,差異顯著;在低濃度Cd處理下,添加Zn可促進GSH的合成,且隨著Zn濃度的增大GSH的合成呈上升趨勢;高濃度Cd處理下,GSH含量隨Zn濃度的增大呈先上升后下降的趨勢,當Zn濃度為10μmol·L-1時,GSH含量達到最大值。表明Zn可顯著增強水稻抗逆境脅迫的能力,一方面水稻幼苗內產生大量的GSH,有效地清除活性氧和自由基,另一方面合成PCs,螯合重金屬緩解重金屬的毒害。
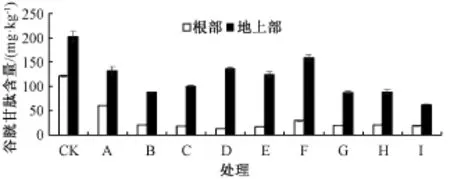
圖3 Cd脅迫下各處理對水稻幼苗谷胱甘肽含量的影響
3 小結與討論
Cd脅迫影響水稻幼苗的正常生長,引起水稻幼苗生物量的下降。隨著Cd處理濃度的增加,毒害作用基本呈增強的趨勢;Zn對低濃度Cd脅迫的水稻幼苗有緩解效應,這種效應隨著Zn濃度的升高呈先升高后下降的趨勢,當Zn濃度為10μmol· L-1時,能夠極大地減輕Cd脅迫對水稻幼苗生物量的影響,效應最為明顯。Zn緩解Cd毒害的效應受Cd濃度的制約,當Cd濃度過高(50μmol· L-1)時,Zn不僅沒有緩解Cd對水稻幼苗的毒害效應,還產生協同毒害效應。原海燕等[16]研究也發現單獨施加Cd處理導致馬藺生物量降低,Zn的施加一定程度上緩解了Cd的毒害作用。
對于大多數作物而言,根系是重金屬積累的主要部位[17]。本研究表明,重金屬脅迫下,水稻根部重金屬含量明顯高于地上部,這與Wang等[18]研究認為的重金屬脅迫下PCs以及重金屬在根部大量積累相一致。可能是水稻根系直接吸收營養物質,在吸收營養物質的同時也吸收大量的重金屬元素,且這些重金屬容易在根中積累,導致根中重金屬含量高。這對于水稻忍耐Cd的傷害是有利的,可以減少重金屬向地上部乃至籽粒的運輸,減少其通過食物鏈危害人類健康的可能性。
谷胱甘肽具有很強的氧化還原特性,在防衛植物細胞免受活性氧攻擊方面起著重要作用[19];植物螯合肽是重金屬誘導產生的一類多肽,能與重金屬結合并在細胞的液泡內形成區室化以減少重金屬對細胞的損害[20-22],二者在高等植物耐受重金屬脅迫機制中具有重要作用[8]。本試驗表明,水稻根部和地上部PCs和GSH對Cd脅迫響應敏感;隨脅迫濃度的增大,GSH水平相應下降,PCs誘導量相應增加,此現象說明在根部已受氧化脅迫的情況下,水稻幼苗啟動了合成PCs的機制,使Cd在根部細胞內形成區室化阻止對根部的損傷,且GSH和PCs二者表現出相互消長的關系,表明GSH是PCs合成的前體。Cd脅迫對水稻產生了一定的毒害,Zn的添加一定程度上緩解了Cd的毒性并伴隨著水稻根內PCs誘導量的降低,Cd含量降低,可能是Cd,Zn在吸收上出現競爭效應,Zn顯著降低了Cd的吸收,因Cd對PCs的誘導效果遠大于Zn,所以PCs的合成相應地降低。植物體內植物螯合肽的產生與重金屬的濃度有關,其合成水平的高低與細胞內脅迫金屬的數量存在顯著相關關系[23-24],細胞內PCs的誘導量可反映金屬的實際毒性,因此PCs和GSH可作為生物標記物用于Cd污染的早期預警和生態風險評價。
綜上所述,Zn能夠緩解Cd脅迫對水稻的毒害,但這種作用有一定的局限性,當超過一定濃度時,將與重金屬Cd一起協同破壞水稻的生長發育,加重對水稻的毒害。只有在掌握Zn濃度的前提下,才能有效發揮Zn元素對有害離子的緩解毒害效應,促進水稻的生長發育。
[1] 黃秋嬋,黎曉峰,沈方科,等.硅對水稻幼苗鎘的解毒作用及其機制研究[J].農業環境科學學報,2007,26(4):91-94.
[2] Yao W,Sun L,Zhou H,et al.Additive,dominant parental effects control the inheritance of grain cadmium accumulation in hybrid rice[J].Molecular Breeding,2015,35(1):1-10.
[3] 康浩,石貴玉,潘文平,等.鎘對植物毒害的研究進展[J].安徽農業科學,2008,36:11200-11201.
[4] Yadav SK.Heavymetals toxicity in plants:an overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants[J].South African Journalof Botany,2010,76(2):167-179.
[5] Bhargava P,Srivastava A K,Urmil S,et al.Phytochelatin plays a role in UV?B tolerance in N2-fixing cyanobacterium Anabaena doliolum[J].Journal of Plant Physiology,2005,162(11):1220-1225.
[6] Petra J,Gudrun K,Sieglinde M,et al.Cadmium induces a novelmetallothionein and phytochelatin 2 in an aquatic fungus[J].Biochemical and Biophysical Research Communications,2005,333(1):150-155.
[7] 胡朝華,張蕾,朱端衛.植物螯合肽的生物合成與解毒機制及在重金屬修復中的應用前景[J].華中農業大學學報:自然科學版,2006,25(5):575-580.
[8] Grill E,Loffler S,Winnacker E L,et al.Phytochelatins,the heavy?metal?binding peptides of plants,are synthesized from glutathione by a specificγ?glutamylcysteine dipeptidyl transpeptidase(phytochelatin synthase)[J].Proceedings of the National Academy of Sciences,1989,86(18):6838-6842.
[9] Dhankhar R,Sainger PA,Sainger M.Phytoextraction of Zinc:physiological and molecularmechanism[J].Soil and Sediment Contamination,2012,21(1):115-133.
[10] He P P,Lv X Z,Wang G Y.Effects of Se and Zn supplementation on the antagonism against Pb and Cd in vegetables[J].Environment International,2004,30(2):167-172.
[11] Bunluesin S,Pokethitiyook P,Lanza G R,et al.Influences of Cadmium and Zinc interaction and humic acid on metal accumulation in Ceratophyllum demersum[J].Water Air and Soil Pollution,2007,180(1/4):225-235.
[12] Hart J J,Welch R M,Norvell W A,et al.Zinc effects on cadmium accumulation and partitioning in near?isogenic lines of durum wheat that differ in grain cadmium concentration[J]. New Phytologist,2005,167(2):391-401.
[13] 羅瓊,王昆,許靖波,等.我國稻田鎘污染現狀·危害·來源及其生產措施[J].安徽農業科學,2014(30):10540-10542.
[14] 許萍,陳銘學,牟仁祥,等.ICP?MS混合模式測定植物性農產品中的9種痕量元素[J].分析測試學報,2011,30(10):1138-1142.
[15] Cao Z Y,Sun L H,Mou R X,et al.A novelmethod for the simultaneous analysis of seven biothiols in rice(Oryza sativa L.)using hydrophilic interaction chromatography coupled with electrospray tandem mass spectrometry[J].Journal of Chromatography B,2014,976-977:19-26.
[16] 原海燕,黃蘇珍,郭智,等.鋅對鎘脅迫下馬藺生長、鎘積累及生理抗性的影響[J].應用生態學報,2007,9(18):2111-2116.
[17] 邵國勝,謝志奎,張國平.雜草稻和栽培稻氮代謝對鎘脅迫反應的差異[J].中國水稻科學,2006,20(2):189-193.
[18] Wang H O,Zhong G R,Liu X F,et al.The study on sulfhydryl compound to themechanism of detoxification in wheat under copper and cadmium stress[J].Acta Agriculturae Boreali?Sinica,2008,23(3):158-161.
[19] Wójcik M,Tukiendorf A.Phytochelatin synthesisand cadmium localization in wild type of Arabidopsis thaliana[J].Plant Growth Regulation,2004,44(1):71-80.
[20] Ovecˇka M,TakácˇTM.Managing heavymetal toxicity stress in plants:biologicaland biotechnological tools[J].Biotechnology Advances,2014,32(1):73-86.
[21] Luo Z B,Wu C,Zhang C,etal.The role of ectomycorrhizas in heavymetal stress tolerance of host plants[J].Environmental and Experimental Botany,2014,108(12):47-62.
[22] Wood B A,Feldmann J.Quantification of phytochelatins and their metal(loid)complexes:critical assessment of current analytical methodology[J].Analytical and Bioanalytical Chemistry,2012,402(10):3299-3309.
[23] Anjum N A,Ahmad I,Mohmood I,et al.Modulation of glutathione and its related enzymes in plants’responses to toxic metals and metalloids?a review[J].Environmental and Experimental Botany,2012,75:307-324.
[24] 丁戈,彭佳師,張國斌,等.植物螯合肽合酶基因AtPCS2的表達調控[J].中國科學:生命科學,2013,43(12):1112-1118.
(責任編輯:侯春曉)
S 511
A
0528?9017(2015)11?1731?05
文獻著錄格式:于莎莎,卞英芳,楊歡,等.鎘脅迫下鋅對水稻植物螯合肽合成的影響[J].浙江農業科學,2015,56(11):1731-1735.
DOI 10.16178/j.issn.0528?9017.20151111
2015?09?14
浙江省科技專項重大農業項目(2014C02002?1)
于莎莎(1991-),女,山東威海人,在讀碩士研究生,研究方向為農產品質量與食物安全。E?mail:yushasha91@126.com。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
