低溫保存對4種作物種子超弱發光特性影響規律的研究
2015-03-10 11:05:41張文蘭田茜李群賈文斌戴雙段乃彬
山東農業科學 2014年12期
張文蘭 田茜 李群 賈文斌 戴雙 段乃彬
摘要:為探索超弱發光特性(UWL)與種子活力之間的相關性,并為研究種子活力無損測定新方法提供技術依據,對不同保存年限的玉米、小麥、大豆、水稻種子進行超弱發光和延遲發光測定,并通過數據統計、曲線擬合及作圖,分析種子超弱發光特性與低溫保存時間的關系及延遲發光的變化特征。結果表明,同一品種不同保存年限的種子超弱發光能力差異明顯,隨保存延長超弱發光強度逐漸下降,與發芽勢變化呈現極強的正相關;種子的延遲發光呈現隨貯藏年限延長強度降低而衰減系數先增大后減小的特征。
關鍵詞:低溫保存;作物種子;超弱發光;種子活力
中圖分類號:S510.1文獻標識號:A文章編號:1001-4942(2014)12-0038-05
種子活力檢測方法的種類多達數十種,歸納起來不外乎直接法和間接法,即通過發芽試驗測定田間出苗率的方法和生化方法,這些方法繁瑣、耗時長,會破壞組織以及物理、化學結構,且要求操作人員具備較高的專業操作水平,屬有損檢測方法。特別對于稀有種子,發芽試驗測定將造成不可挽回的資源損失,因此在種質資源保存領域需要一種快速、高效、無損的種子活力檢測方法。
生物組織或細胞在生命活動的代謝過程中,都自發地輻射出一種極其微弱的光子流,這是一種極微弱的準連續光子輻射[1],是生物在生命活動中出現的一種機體自身發光,其強度為每秒每平方厘米上幾個至幾千個光子,波長180~800 nm,稱為生物系統的代謝超微弱發光(Ultra-Weak Luminescence,UWL),是所有活的生物都具有的一種普遍現象[2]。生物代謝的超弱發光廣泛存在于動植物中,反映了生物體與生命活動過程的有關信息。生物超弱發光通常包括兩部分:一種為自發的超微弱發光,與生物體的代謝有關;另一種為外因誘導發光稱為延遲發光(Delayed Luminescence,DL),如光、電離輻射、超聲、化學藥物等外界激勵因素。自身發光和延遲發光與生物的新陳代謝狀態和水平,細胞的分裂、死亡、變異以及細胞間信息的傳遞等許多基本的生命過程都有著內在的聯系,是反映生物體內部機能的一個重要窗口[3]。基于以上,通過超微弱發光特性檢測種子基礎代謝(反映種子的活力)與保存時間的關系,在不破壞被檢測種子的物理和化學結構情況下,對種子進行無損、快速檢測,將為種質資源的安全保存與及時更新提供更為準確的技術依據[4,5]。
1材料與方法
1.1試驗材料
從種子資源庫(中期庫)選取玉米鄭單958、小麥濟南16、大豆齊黃27、水稻香粳9407共4種作物種子,其保存時間分別為1、3、5、7、9年,保存溫度為0℃。種子由中國農業科學院種質資源研究所國家種質中期庫提供。
1.2試驗方法
1.2.1發芽勢及發芽率的測定取以上玉米、小麥、大豆、水稻凈種子,按照《農作物種子檢驗規程》(GB/T 3543.3-1995)操作規程進行發芽試驗,統計發芽勢和發芽率。
1.2.2自身發光的測量從0℃中期庫中取出種子,置于4℃保鮮箱中24 h緩慢升溫,再置于25℃生化培養箱中緩慢升溫并保持恒溫,使之不因升溫破壞種子的基礎代謝,盡量保持種子保存時的自然狀態。每個保存年限的種子樣品分成3個平行組,其中玉米每組10粒、小麥50粒、大豆15粒、水稻70粒,以便使種子數量能夠完全覆蓋測量杯底并略有盈余。在進行試驗前種子先置于暗室中30 min,取出后放入BPCL-2-ZL超微弱發光測量儀樣品室的測量杯中進行自身發光的測定,時間為100 s,取每種樣品3個平行組測量數據的平均值作為該樣品的測量值,樣品室溫度設定為恒溫25℃。
1.2.3延遲發光的測量由于自身發光主要反映種子的基礎代謝特征,這對于判斷種子活力并不全面,本試驗進一步測量了種子的延遲發光特征。前人的研究發現新鮮種子采取的激勵方法是日光燈或LED燈珠照射,照度只有幾十勒克斯[7]。而本試驗是經過較長時間保存的種子,較低的照度顯然不足以對樣品產生足夠激勵,為深入研究種子對外界激勵的響應,需要較強的“喚醒”信號,故選擇波長較寬的日光燈為光源,光照強度為4 000 lx。樣品分組方式與自身發光測量相同,將待測樣品依次放在日光燈下照射2 min后,迅速測定延遲發光光子數,每次測量100 s,樣品室設定為恒溫25℃。
2結果與分析
2.1發芽率、發芽勢測定結果
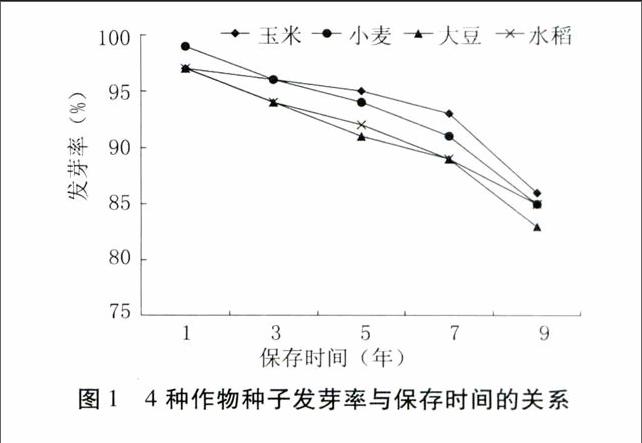
由圖1可知,玉米、小麥、大豆、水稻4種作物種子的發芽率隨保存時間的延長逐漸降低,降低趨勢一致,保存1~7年間緩慢降低,7年后顯著降低。以玉米種子為例,保存1年的種子發芽率為97%,保存3、5、7、9年后的發芽率分別降低為96%、95%、93%和86%。
4種作物種子發芽勢降低趨勢也接近一致。圖2反映出發芽勢的降低幅度比發芽率大。以小麥為例,保存1年后種子發芽勢為99%,保存3、5、7、9年后分別降低為95%、90%、78%和70%。
以上結果表明,在中期庫保存對4種作物種質生活力變化趨勢的影響是一致的,同時說明,在中期庫保存過程中,玉米、小麥、水稻和大豆的耐貯性相差不大。
2.2自身發光檢測結果分析
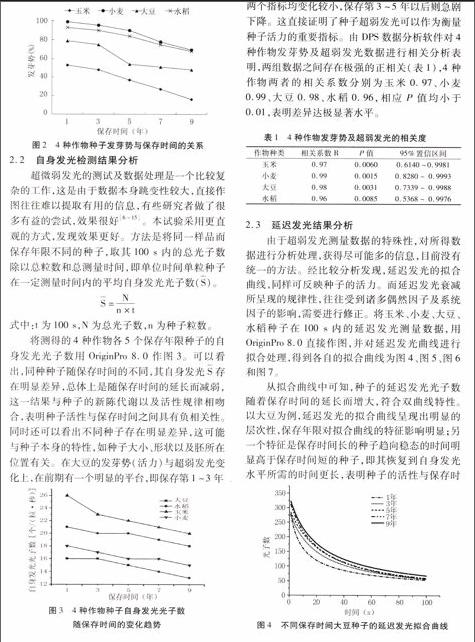
超微弱發光的測試及數據處理是一個比較復雜的工作,這是由于數據本身跳變性較大,直接作圖往往難以提取有用的信息,有些研究者做了很多有益的嘗試,效果很好[6~15]。本試驗采用更直觀的方式,發現效果更好。方法是將同一樣品而保存年限不同的種子,取其100 s 內的總光子數除以總粒數和總測量時間,即單位時間單粒種子在一定測量時間內的平均自身發光光子數(S)。
S=Nn×t
式中:t為100 s,N為總光子數,n為種子粒數。
將測得的4種作物各5個保存年限種子的自身發光光子數用OriginPro 8.0作圖3。可以看出,同種種子隨保存時間的不同,其自身發光S存在明顯差異,總體上是隨保存時間的延長而減弱,這一結果與種子的新陳代謝以及活性規律相吻合,表明種子活性與保存時間之間具有負相關性。同時還可以看出不同種子存在明顯差異,這可能與種子本身的特性,如種子大小、形狀以及胚所在位置有關。在大豆的發芽勢(活力)與超弱發光變化上,在前期有一個明顯的平臺,即保存第1~3年兩個指標均變化較小,保存第3~5年以后則急劇下降。這直接證明了種子超弱發光可以作為衡量種子活力的重要指標。由DPS數據分析軟件對4種作物發芽勢及超弱發光數據進行相關分析表明,兩組數據之間存在極強的正相關(表1),4種作物兩者的相關系數分別為玉米0.97、小麥0.99、大豆0.98、水稻0.96,相應P值均小于0.01,表明差異達極顯著水平。
2.3延遲發光結果分析
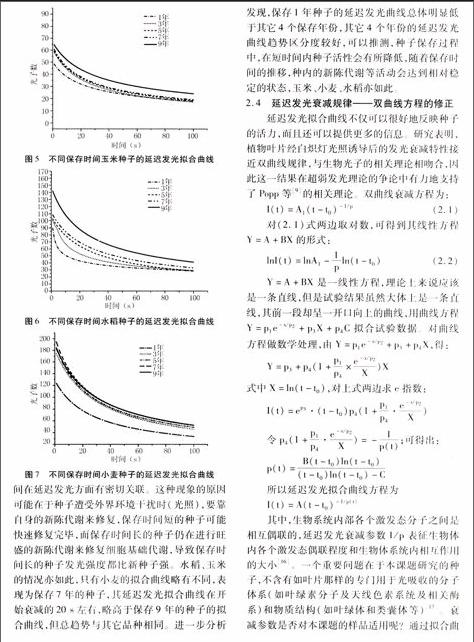
由于超弱發光測量數據的特殊性,對所得數據進行分析處理,獲得盡可能多的信息,目前沒有統一的方法。經比較分析發現,延遲發光的擬合曲線,同樣可反映種子的活力。而延遲發光衰減所呈現的規律性,往往受到諸多偶然因子及系統因子的影響,需要進行修正。將玉米、小麥、大豆、水稻種子在100 s內的延遲發光測量數據,用OriginPro 8.0直接作圖,并對延遲發光曲線進行擬合處理,得到各自的擬合曲線為圖4、圖5、圖6和圖7。
從擬合曲線中可知,種子的延遲發光光子數隨著保存時間的延長而增大,符合雙曲線特性。以大豆為例,延遲發光的擬合曲線呈現出明顯的層次性,保存年限對擬合曲線的特征影響明顯;另一個特征是保存時間長的種子趨向穩態的時間明顯高于保存時間短的種子,即其恢復到自身發光水平所需的時間更長,表明種子的活性與保存時間在延遲發光方面有密切關聯。這種現象的原因可能在于種子遭受外界環境干擾時(光照),要靠自身的新陳代謝來修復,保存時間短的種子可能快速修復完畢,而保存時間長的種子仍在進行旺盛的新陳代謝來修復細胞基礎代謝,導致保存時間長的種子發光強度都比新種子強。水稻、玉米的情況亦如此,只有小麥的擬合曲線略有不同,表現為保存7年的種子,其延遲發光擬合曲線在開始衰減的20 s左右,略高于保存9年的種子的擬合曲線,但總趨勢與其它品種相同。進一步分析發現,保存1年種子的延遲發光曲線總體明顯低于其它4個保存年份,其它4個年份的延遲發光曲線趨勢區分度較好,可以推測,種子保存過程中,在短時間內種子活性會有所降低,隨著保存時間的推移,種內的新陳代謝等活動會達到相對穩定的狀態,玉米、小麥、水稻亦如此。
2.4延遲發光衰減規律——雙曲線方程的修正
延遲發光擬合曲線不僅可以很好地反映種子的活力,而且還可以提供更多的信息。研究表明,植物葉片經白熾燈光照誘導后的發光衰減特性接近雙曲線規律,與生物光子的相關理論相吻合,因此這一結果在超弱發光理論的爭論中有力地支持了Popp等[9]的相關理論。雙曲線衰減方程為:
Y=A+BX是一線性方程,理論上來說應該是一條直線,但是試驗結果雖然大體上是一條直線,其前一段卻呈一開口向上的曲線,用曲線方程Y=p1e-x/p2+p3X+p4C擬合試驗數據。對曲線方程做數學處理,由Y=p1e-x/p2+p3+p4X,得:
Y=p3+p4(1+p1p4×e-x/p2X)X
式中X=ln(t-t0),對上式兩邊求e指數:
I(t)=ep3·(t-t0)p4(1+p1p4·e-x/p2X)
令p4(1+p1p4·e-x/p2X)=-1p(t);可得出:
p(t)=B(t-t0)ln(t-t0)(t-t0)ln(t-t0)-C
所以延遲發光擬合曲線方程為
I(t)=A(t-t0)-1/p(t)
其中,生物系統內部各個激發態分子之間是相互偶聯的,延遲發光衰減參數1/p表征生物體內各個激發態偶聯程度和生物體系統內相互作用的大小[16]。一個重要問題在于本課題研究的種子,不含有如葉片那樣的專門用于光吸收的分子體系(如葉綠素分子及天線色素系統及相關酶系)和物質結構(如葉綠體和類囊體等)[17]。衰減參數是否對本課題的樣品適用呢?通過擬合曲線方程,可以比較方便地求出衰減參數,作圖后發現保存年限也與衰減參數存在相關性,但是關系較復雜,如圖8所示。表現為在1~5年間衰減參數隨保存時間延長而增大,5年時達到最大,隨后減小。由于衰減參數相對于自身發光及延遲發光的測量而言,是以擬合曲線方程為前提的,經模擬處理后,可能會造成信息的丟失,因此雖然衰減參數與保存時間并不呈現為簡潔的關系,但是它們之間的關聯性,使得衰減參數仍可以作為檢測種子活力的輔助參數。
由于測量樣品數量還不太豐富,同時旁證材料較少,相關的分析有待于進一步深入研究。
3結論
通過測量4種作物種子的20個樣品的自身發光和光照誘導條件下的延遲發光特性,用作圖和曲線擬合分析試驗數據,發現儲藏時間會導致玉米、小麥、大豆、水稻種子組織結構及生理生化上的變化,相應地反映在其自身發光強度和延遲發光變化上。
在自身發光(UL)方面,保存的時間越長,種子自身的發光強度越弱,體現了它們之間的負相關性,單位時間單粒種子的平均自發光子數,可很好地反映種子的新陳程度,這可作為無損傷檢測種子保存時間的主要指標。
在延遲發光(DL)方面,保存時間的不同會導致延遲發光特性的差異。在強光激勵對種子“喚醒”后,保存時間長的種子趨向穩態的時間要明顯高于保存時間短的種子,表明了它們之間的正相關性。這種特性可以很好地將延遲發光作為反映種子內部新陳代謝變化的一個窗口,擬合曲線能夠很好地反映種子的活力,衰減參數也與保存時間存在關聯性,可作為無損傷檢測種子活力的輔助參數。
總之,相對于傳統檢測方法及其它有損傷檢測,自身發光和延遲發光方法可以很好地反映種子活力,這對于提高檢測效率,降低種子損耗,是一種有效快速簡便的方法。
參考文獻:
[1]聶繼云,彭運生. 生物超弱發光及其應用研究概述[J]. 激光生物學報,1998,7(2):124-130.
[2]陳勝,黃楚云,李默然. 生物超微弱發光及其應用[J]. 黃石理工學院學報,2006,22(4):82-84.
[3]習崗. 植物超弱發光及其在農業上的應用[J]. 中國糧油學報,2013,26(1):100-102.
[4]李光,陳靜偉,楊海蓮,等. 雙氧水對葉片延遲發光的影響[J]. 河北大學學報,2006,26(4):366-371.
[5]陳文利. 生物光子學技術在水稻種子活力和植物應激反應中的應用研究[D]. 廣州:華南師范大學,2002.
[6]焦煒. 谷物新陳度的快速測定[J]. 糧食流通技術,2011(3):36-39.
[7]鮑杰,吳才章. 基于虛擬儀器的小麥新陳度檢測[J].中國糧油學報,2011,26(6):102-105.
[8]于林平,朱京立,李勇,等. 小麥新陳度快速判斷方法探討[J]. 糧油倉儲科技通訊,2003(6):49-50.
[9]Popp F A. Coherent photon storage of biological systems[C]// In: Popp F A, Becker G,Knig H L,Peschka W.(Hrsg.):Electromagnetic Bio-information. Proceedings of the symposium,Marburg,5. September 1977. Urban & Scharzenberg, München-Wien-Baltimore 1979.
[10]王毅,冀圣江,司建中. 小麥新陳度鑒別方法探討[J]. 糧油倉儲科技通訊,2009(1):48-49.
[11]吳才章,王繼偉. 小麥超弱延遲發光測試系統[J]. 光子學報,2014,43(2):0217001.
[12]梁義濤,朱遠坤,王鋒,等. 不同結構小麥籽粒延遲發光特性[J]. 河南科技大學學報,2013,34(5):69-72.
[13]李德紅,邢達,譚石慈,等. 綠豆和花生的超弱發光[J]. 植物生理學報,1998,24(2):177-182.
[14]李韶山,王艷,郭周義. 萌發花生種子超弱發光的研究[J].光子學報,2000,29(11):966-968.
[15]呂家根,占達東,王周平,等. 酸雨脅迫下小麥微弱延遲發光及其生理生態變化相關性研究[J].化學學報,2003,61 (5):760-764.
[16]陳江麗,陸治國. 蒜的延遲發光[J]. 光電子·激光,1999(4):365-367.
[17]郭海軍,陳靜偉,黃艷賓. 葉片延遲發光衰減方程的研究[J]. 河北大學學報,2010,30(3):255-258.
