Omi/HtrA2 在無鎂外液致癇海馬神經元凋亡中的作用機制及其特異性抑制劑UCF-101 的神經元保護作用
2015-03-10 09:03:18連亞軍張海峰謝南昌高延倫
中風與神經疾病雜志 2015年11期
金 迪,連亞軍,張海峰,謝南昌,高延倫
癲癇是多種原因導致的腦部神經元高度同步化異常放電所致的臨床綜合征[1],其發作形式復雜,反復發作給患者的生活帶來了極大的痛苦,因此明確癲癇的發病機制、尋找新的治療靶點尤為重要。近年來,癲癇導致神經元凋亡的相關分子機制受到人們的很大關注,外來因素致癇神經元后,壞死的、凋亡的以及抗凋亡的信號途徑均被激活,它們之間的平衡被破壞,促凋亡蛋白表達相對更高,進而促進神經元凋亡的發生[2]。如何精確調控神經元凋亡,有望成為抗癲癇治療的新靶點。Omi/HtrA2 是一種定位于線粒體膜間隙的絲氨酸蛋白酶,是細胞內線粒體凋亡途徑中重要的信號分子。既往有研究證明,Omi/HtrA2 在腦缺血/再灌注損傷[3]、腎臟缺血再灌注損傷以及高氧誘導的肺泡上皮細胞凋亡等疾病中,均發揮重要作用。但其是否參與體外癲癇模型中神經元的凋亡過程,目前國內外尚缺乏這方面的研究。本研究通過Sombati 法建立海馬神經元自發性癲癇樣放電的細胞模型,該模型與臨床顳葉癲癇較為貼近,旨在觀察Omi/HtrA2 在體外癲癇模型神經元凋亡中的作用機制,以及應用其特異性抑制劑UCF-101 后,對神經元凋亡以及XIAP、caspase3等蛋白表達的影響,探討UCF-101 的神經保護作用,為臨床上癲癇的治療提供理論依據。
1 材料和方法
1.1 動物和主要試劑 新生24 h 以內SD 大鼠,SPF 級[鄭州大學動物實驗中心提供,許可證號SCxK(豫)2010-0002];DMEM/HIGH GLUCOSE 培養基、PBS 液(HyClone);轉鐵蛋白、牛胰島素、L-多聚賴氨酸、胰蛋白酶(Sigma);特級胎牛血清、B27 及Glutamax-I 谷氨酰胺(Gibco);青鏈霉素混合液(北京索萊寶);NSE 多克隆抗體(北京博奧森生物技術有限公司);山羊抗兔IgG/Cy3 熒光二抗(北京博奧森生物技術有限公司);基礎培養基:DMEM+0.2%轉鐵蛋白+0.4%NaHCO3+1%青鏈霉素混合液;種植液:90%基礎培養基+10%FBS+1%Glutamax-I+2%牛胰島素;維持液:93%基礎培養基+2%B27 +0.25% Glutamax-I+5% FBS;無鎂細胞外液(pH 7.4):145 mmol/L NaCl、2.5 mmol/L KCl,10 mmol/L HEPES,2 mmol/L CaCl2,10 mmol/L 葡 萄 糖,0.002 mmol/L甘氨酸;正常細胞外液:無鎂外液各成分+1 mmol/L MgCl2;Anti-active caspase-3 抗體(abcam);Anti-HtrA2/Omi 抗體(abcam);Anti-XIAP 抗體(santa cruz);Anti-HAX-1 抗體(santa cruz)。線粒體和胞漿蛋白分離提取試劑盒(上海生工)。
1.2 原代海馬神經元的培養 新生24 h 內的SD 大鼠,常規消毒,斷頭取腦,置于加有預冷PBS的培養皿中,分離雙側海馬,剔除表面血管,并剪成約1 mm×1 mm×1 mm 的組織塊,轉移至15 ml 的離心管中,加入0.125%的胰酶后,置于37 ℃恒溫箱中消化15 min,期間每隔數分鐘輕微振蕩,消化完畢后1500 r/min 離心5 min,加入PBS 洗滌并終止消化,棄上清,加入種植液,吹打均勻制成細胞懸液。取少量細胞懸液用于臺盼藍染色計數,調整細胞密度為5 × 105/ml,接種于6 孔培養板中(預先以0.01%L-多聚賴氨包被),每孔約2 ml。置于37.5 ℃恒溫,5% CO2培養箱中培養24 h 后,等量換成含有B27 的維持液,以后每周2~3 次進行半量換液,并于倒置相差顯微鏡下觀察細胞生長情況,培養至7~10 d 用于后續實驗[4]。
1.3 NSE 染色鑒定海馬神經元 海馬神經元在放置有爬片的24 孔板中生長至7 d 后,進行NSE鑒定。吸棄培養基,PBS 漂洗1 遍,然后用4%多聚甲醛固定20 min,PBS 漂洗3 遍,每次5 min;0.25%Triton 穿孔15 min,吸棄Triton,PBS 漂洗3 次,每次5 min;1%BSA 室溫下封閉30 min,加入兔抗大鼠NSE 多克隆抗體(稀釋200 倍)4℃過夜,TBST 漂洗3 次,每次5 min,加入山羊抗兔IgG/Cy3 熒光二抗,以下操作步驟均需避光進行,37 ℃作用1 h;TBST漂洗3 次,每次5 min,5 μg/ml DAPI 染色2 min,吸棄DAPI,TBST 漂洗,抗熒光猝滅封片劑封片,放入濕盒避光保存,擇期在熒光顯微鏡下觀察。
400 倍熒光顯微鏡下隨機選擇10 個視野,以視野中陽性神經元數目占觀察細胞總數的百分率為神經元純度。
1.4 造模分組
1.4.1 海馬神經元培養至10 d,隨機分為對照組和模型組 對照組用正常細胞外液培養3 h 后,更換為正常維持液繼續培養。模型組以無鎂細胞外液作用3 h 后,更換為正常維持液,并分別于造模后3 h、8 h、24 h 終止細胞培養,用于后續試驗。
1.4.2 海馬神經元培養至10 d,隨機分為對照組、致癇24 h 組、UCF-101 干預組、DMSO 組 其中UCF-101 干預組,在無鎂細胞外液作用前,加入10 μmol/L UCF-101(溶于DMSO 中)事先作用24 h,再行無鎂細胞外液誘導,造模后繼續培養24 h,終止培養。DMSO 組致癇前給予相同濃度的DMSO 作用24 h,再行無鎂外液誘導,造模后繼續培養24 h,終止培養。
1.5 western blot 法檢測Omi/HtrA2、caspase3、XIAP、HAX-1 蛋白表達變化 分別提取各組海馬神經元線粒體蛋白、胞漿蛋白及細胞總蛋白。BCA 法測蛋白濃度,調整上樣量,每個泳道保證總蛋白量相等。SDS-PAGE 凝膠電泳(上層膠80 V 30 min;下層膠120 V 60 min),電泳結束后將膠上的蛋白質條帶轉移到PVDF 膜上(半干轉),5%脫脂奶粉封閉3 h。一抗孵育,兔抗大鼠Omi/HtrA2(1∶ 1000);兔抗大鼠caspase3(1 ∶ 200);兔抗大鼠XIAP(1 ∶200);兔抗大鼠HAX-1(1 ∶ 200),4 ℃冰箱過夜。TBST 洗膜每次10 min,共3 次,洗膜后加入辣根過氧化物酶標記山羊抗兔IgG(1∶ 8000),室溫搖床緩慢孵育1 h,ECL 顯影成像重復3 次。采用Image J測定條帶光密度,以目的蛋白與相應內參條帶光密度比值作為目的蛋白的相對表達量。
1.6 TUNEL 法檢測神經元凋亡 采用TUNEL法原位標記DNA 片段來檢測凋亡細胞數量,具體步驟按試劑盒提供的說明書進行操作,熒光素染色,熒光顯微鏡下進行觀察,每片選取10 個高倍視野,分別計數陽性細胞數,重復3 次,取其均數作為該例凋亡細胞數。
2 結果
2.1 海馬神經元形態學特征 海馬神經元剛接種時形態基本呈圓形,體積較小,單個分布。培養3 h 后逐漸開始貼壁,24 h 后絕大部分細胞貼壁,而且部分神經元長出短小突起,少數未能貼壁的細胞死亡。隨著培養時間的延長,神經元突起逐漸增多并延長,粗細不等,有的突起在遠端形成分支,3 d后明顯增長的突起開始相互連接成稀疏的網絡。神經元生長至7 d,細胞間連接更為緊密,立體感較強,神經元胞體大多呈橢圓形或三角形,此時細胞較為豐滿,突起清晰,神經元網絡密集(見圖1)。

圖1 倒置相差顯微鏡下觀察體外培養不同時間新生大鼠海馬神經元(A:24 h;B:3 d:C:5 d;D:7 d)
2.2 海馬神經元純度測定 海馬神經元培養至7 d,經神經元特異性烯醇化酶NSE 染色鑒定,純度為90%以上(見圖2)。

圖2 培養至7 d 的海馬神經元NSE 免疫熒光染色
2.3 Western Blot 檢測各組海馬神經元中Omi/HtrA2、caspase3、XIAP、HAX-1 蛋白表達的變化
2.3.1 Omi/HtrA2 在海馬神經元中的表達以及致癇后其位置的改變 Western Blot 結果顯示,正常對照組中Omi/HtrA2 主要位于線粒體中,即線粒體中的蛋白表達量較高,隨著造模后時間的延長,Omi/HtrA2 逐漸釋放入胞漿內,這種移動尤其在致癇后8 h、24 h 較為明顯,與正常對照組比較有統計學差異(P <0.05)。而致癇后3 h 線粒體中的Omi/HtrA2 表達量較正常組線粒體中表達量有所上升,存在統計學差異(P <0.05)。由此說明,海馬神經元致癇后,Omi/HtrA2 在線粒體中的含量短時間內有所上升,隨后發生了顯著的移位,即由線粒體移位到了胞漿中(見圖3)。
2.3.2 UCF-101 對各組海馬神經元中XIAP、active-caspase3、HAX-1 蛋白表達的影響 正常對照組中HAX-1 表達相對較高,致癇24 h 后,HAX-1 蛋白表達量顯著下降,與正常對照組相比有統計學差異(P <0.05)。預先使用UCF-101 作用后,HAX-1表達量有所上升,與模型組有統計學差異(P <0.05)。DMSO 組與模型組相比,無統計學差異(P>0.05)(見圖4)。XIAP 表達量在模型組或DMSO組中均有所升高,但與正常組比較均無統計學差異,使用UCF-101 預處理后,XIAP 表達量較另外3 組有明顯的升高,存在統計學差異(P <0.05)。正常對照組海馬神經元中active-caspase3 的蛋白表達量很低,模型組中此蛋白表達量明顯上調,與正常對照組比較有統計學意義(P <0.05)。UCF-101 處理組中此蛋白表達量明顯低于模型組(P <0.05),DMSO組與模型組相比無統計學差異(P >0.05)(見圖5)。

圖3 WB 檢測各組Omi/HtrA2 在線粒體和胞漿中的相對含量

圖4 WB 檢測各組海馬神經元中HAX-1 蛋白相對含量

圖5 WB 檢測各組海馬神經元中XIAP 蛋白以及activecaspase3 蛋白相對含量
2.4 UCF-101 對體外癲癇模型中神經元凋亡的影響 正常對照組熒光顯微鏡下觀察偶見少量TUNEL 染色陽性的細胞,模型組(致癇24 h 組)鏡下可見大量凋亡細胞,與正常對照組相比有統計學差異(P <0.05)。UCF-101 組凋亡細胞數較模型組顯著減少,與模型組相比有統計學差異(P <0.05)。DMSO 組凋亡細胞數與模型組相比無統計學差異(P >0.05)(見圖6)。
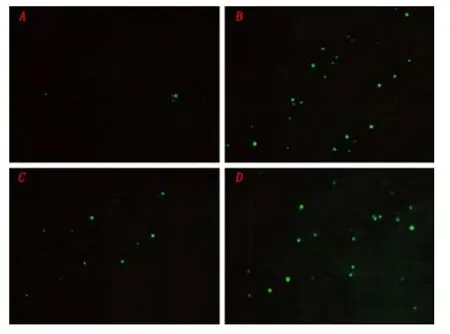
圖6 TUNEL 染色檢測神經元凋亡:A 正常組B 致癇24 h 組C UCF-101 處理組D DMSO 組
3 討論
癲癇是僅次于腦卒中的最常見的神經退行性疾病[5],近年來,線粒體在后天獲得性癲癇例如顳葉癲癇中的作用日益受到人們重視。線粒體中某些蛋白在癲癇發作過程中從線粒體釋放入胞漿,從而對促凋亡、抗凋亡、壞死途徑的激活產生一定影響。因此,癲癇發作過程中發生亞細胞移位的蛋白,有望成為癲癇治療的新靶點。
IAPs 是細胞內可以調控caspase 活性的凋亡抑制因子,這些因子包括XIAP、c-IAP1、c-IAP2 等。據報道這些凋亡抑制因子通過直接抑制起始caspase和效應caspase 來阻斷凋亡途徑[6]。IAPs 包含一個或多個保守區域,即BIRs,這些BIR 區域在抑制caspase 活性方面發揮重要作用。BIR 區域和它們之間的連接部分直接與caspase 結合,并抑制其活性,但IAPs 的抗凋亡活性又可以被一些蛋白所調控,這些蛋白連接在BIR 區域上,通過破壞caspase-IAPs 復合體或促使IAPs 泛素化降解來激活caspase活性,其中就包括Omi/HtrA2。Omi/HtrA2 是一種位于線粒體膜間隙的促凋亡的絲氨酸蛋白酶,據報道它主要通過兩種途徑促進細胞發生凋亡,即caspase 依賴性途徑和caspase 非依賴性途徑。和AIF、細胞色素C 一樣,Omi/HtrA2 在凋亡過程中,從線粒體膜間隙釋放到胞漿中,通過與IAPs 結合而激活caspase,發揮其促凋亡作用,此即caspase 依賴性促凋亡途徑[7]。但也有文獻報道,Omi/HtrA2 可以單獨依賴其蛋白酶活性發揮caspase 非依賴性促凋亡作用。
HAX-1 是一個分子量為35Kd 的抗凋亡蛋白,與BCL-2 家族的BH1 區和BH2 區有同源性。其可以在體內或體外條件下通過Omi/HtrA2 的蛋白酶活性被裂解,而且其降解在凋亡過程中發生較早,Cilenti 等[8]早在2004 年就研究得出在Omi/HtrA2尚位于線粒體時,線粒體中的HAX-1 就已經被其降解了。此項研究表明Omi/HtrA2 在線粒體中就可以通過裂解HAX-1 發揮其促凋亡作用,而這種作用與其在胞漿中通過裂解XIAP,激活caspase 發揮促凋亡作用,顯然是不同的。據相關文獻報道[9],來自mnd2 鼠的細胞系攜帶有突變的Omi/HtrA2 基因,從而影響其蛋白酶活性,當給這種細胞系誘導凋亡時,HAX-1 幾乎不發生降解。而當功能性的Omi/HtrA2 基因導入此種細胞系后,HAX-1 發生了降解。而用Omi/HtrA2 的特異性抑制劑UCF-101 預先作用于細胞后,HAX-1 的降解明顯減少,這些結論在Cilenti 等的研究中也都有提到。除此之外,既往有學者以出生14 d 的Wistar 大鼠為研究對象,以氯化鋰-匹魯卡品制造癲癇持續狀態模型,研究Omi/Htra2 在癲癇發生過程中的作用機制,免疫熒光雙標顯示幾乎所有caspase3 陽性的細胞胞漿中都高表達Omi/HtrA2,但并不是所有高表達Omi/HtrA2 的細胞都是caspase3 陽性[10],由此也提示Omi/HtrA2 可以通過caspase 非依賴性的方式促進細胞發生凋亡。
神經元體外原代培養模型是研究神經元發育、分化、神經再生、神經系統疾病的發生機制等眾多領域的重要模型,近年來在癲癇的研究中越來越受到重視。本次研究創新性的以體外培養的新生大鼠海馬神經元為研究對象,利用Sombati 法制造體外癲癇模型。進一步探索了Omi/HtrA2 在癲癇過程中的亞細胞移位,以及其對caspase3、XIAP、HAX-1 蛋白表達的影響。
研究結果得出,Omi/HtrA2 于細胞致癇后3h 在線粒體中的表達升高,隨后于致癇后8 h、24 h 發生了顯著的移位,胞漿中的Omi/HtrA2 含量在細胞致癇后3 h、8 h、24 h 均有提高,其中以8 h、24 h 最為顯著。其移位于胞漿中后,通過裂解XIAP 發揮其caspase 依賴性的致凋亡作用。但本次研究得出XIAP 表達在正常組與致癇24 h 組并無統計學差異,初步考慮可能是由于細胞遭遇應激反應后,為了維持細胞功能和結構的完整性,海馬神經元內的XIAP保護性的增多,但隨著時間的延長,Omi/HtrA2 逐漸從線粒體釋放入胞漿內,裂解并中和XIAP,又使其表達量下降,兩種作用綜合起來導致XIAP 表達量與正常組相比無明顯差異。但使用UCF-101 預處理后,其特異性的抑制Omi/HtrA2 的活性,而減弱了Omi/HtrA2 對XIAP 的降解作用,導致XIAP 表達量有明顯的提升。XIAP 被裂解后,解除了其對caspase3 的抑制作用,導致active-caspase3 蛋白的表達明顯上升。而HAX-1 在Omi/HtrA2 的作用下顯著減少,使用UCF-101 后HAX-1 表達量上升,差異有統計學意義,這與Lucia Cilenti 等人的研究結果是一致的。致癇24 h 后,TUNEL 染色結果顯示神經元凋亡數目顯著增多,而使用UCF-101 后凋亡細胞有所減少,就是由于UCF-101 特異性的抑制了Omi/HtrA2 的促凋亡活性,從而抑制了神經元的凋亡。
本次研究得出在無鎂外液誘導的神經元癇性放電過程中,Omi/HtrA2 發生了明顯移位,并且通過降解XIAP、HAX-1 發揮其caspase 依賴性和caspase 非依賴性的兩種促凋亡方式,使用其特異性抑制劑UCF-101 后,上述兩種蛋白降解明顯減弱,抑制了caspase 的激活及神經元的凋亡,對致癇海馬神經元發揮一定的保護作用。但更需要進一步的研究,明確HAX-1、XIAP 被其裂解的具體時相,且UCF-101作為其特異性抑制劑,還有沒有別的不為人知的作用與抗凋亡相關,尚待進一步研究。
[1]Ferriero DM.Protecting neurons[J].Epilepsia,2005,46(Suppl 7):45-51.
[2]Althaus J,Siegelin MD,Dehghani F et al.The serine protease Omi/HtrA2 is involved in XIAP cleavage and in neuronal cell death following focal cerebral ischemia/reperfusion[J].Neurochem Intern,2007,50(1):172-180.
[3]張 偉,崔艷艷,姜寰宇,等.UCF-lOl 對大鼠腦缺血再灌注損傷的神經保護作用[J].解剖科學進展,2010,16(4):335-337.
[4]徐祖才,徐 平,張 駿,等.新生大鼠海馬神經元原代培養及膜片鉗全細胞記錄[J].重慶醫學,2012,41(31):3241-3242.
[5]Sloviter RS.The neurobiology of temporal lobe epilepsy:too much information,not enough knowledge[J].Comptes Rendus Biol,2005,328(2):143-153.
[6]Deveraux QL,Leo E,Stennicke HR,et al.Cleavage of human inhibitor of apoptosis protein XIAP results in fragments with distinct specificities for caspases[J].EMBO J,1999,18(19):5242-5251.
[7]Verhagen AM,Silke J,Ekert PG,et al.Omi/HtrA2 promotes cell death through its serine protease activity and its ability to antagonize inhibitor of apoptosis proteins[J].J Biol Chem,2002,277(1):445-454
[8]Cilenti.L,Soundarapandian MM,Zervos AS.Regulation of HAX-1 anti-apoptotic protein by Omi/HtrA2 protease during cell death[J].J Biol Chem,2004,279(48):50295-50301.
[9]Jones JM,Datta P,Srinivasula SM,et al.Loss of Omi mitochondrial protease activity causes the neuromuscular disorder of mnd2 mutant mice[J].Nature,2003,425(6959):721-727.
[10]Rami A,Kim M.Translocation of the Serine Protease Omi/HtrA2 from Mitochondria into the Cytosol Upon Seizure-Induced Hippocampal Injury in the Neonatal Rat Brain[J].Neurochem Res,2010,35(12):2199-2207.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
中國民間療法(2021年5期)2021-06-09 09:21:04
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
飲食科學(2017年5期)2017-05-20 17:11:53
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
光學精密工程(2016年6期)2016-11-07 09:07:19
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
