穩定轉染CD151 或CD163 的Marc145 細胞系的建立及其在PRRS疫苗生產中的應用
2015-03-18 07:48:28劉開武吳建英王蓮芳胡小明黃海軍
中國獸醫雜志 2015年6期
阮 征 ,劉開武,吳建英,華 娟,夏 瑜,王蓮芳,胡小明,劉 杰,,黃海軍
(1.武漢市農科院畜牧獸醫科學研究所,湖北 武漢430208 ;2.武漢市重大動物疫病防控中心,湖北 武漢 430023;3.華中農業大學農業動物遺傳育種與繁殖教育部重點實驗室,湖北 武漢 430070)
豬繁殖與呼吸綜合征(Porcine reproductive and respiratory syndrome,PRRS)是豬群中流行的重要傳染病之一,可引起母豬的繁殖障礙(包括流產、早產、死胎、木乃伊胎等)、仔豬存活率急劇下降和育成豬呼吸道疾病。病原為豬繁殖與呼吸綜合征病毒(PRRSV),該病毒為單股正鏈RNA病毒,屬于動脈炎病毒屬。PRRSV有嚴格的宿主和細胞嗜性,豬肺泡巨噬細胞(Porcine alveolar macrophage,PAM)、Marc145 細胞和MA-104 細胞是其重要的靶細胞,PRRSV感染細胞需要首先與細胞受體結合,通過內吞作用進人細胞內,在胞內完成病毒的脫殼與基因組釋放[1-3]。
目前發現的PRRSV感染相關受體有硫酸乙酰肝素(Heparin Sulphate,HS)、唾液酸粘附素(Sialoadhesin,Sn)、波形蛋白(Vimentin)、CD163(Cluster of Differentiation 163)分子、CD151(Cluster of Differentiation 151)分子和NMMHCIIA(非肌肉肌球蛋白重鏈ⅡA)分子[4]。有關研究結果表明,CD151 能與PRRSV3′端非轉錄區(3′URT)發生結合,進而導致PRRSV與細胞發生相互作用進入細胞內產生感染。通過轉染使PRRSV非敏感細胞BHK-21 表達CD151會使該細胞允許PRRSV的感染,增殖程度高達100倍[5]。CD163 為一種SRCR 超家族的Hapten Hemoglobin 清道夫受體,是130 000 的細胞表面糖蛋白,在PRRSV感染巨噬細胞過程中介導病毒在細胞質中的脫衣殼和病毒基因組的釋放[6-8]。
在實際生產中,PRRSV的體外增殖主要在Marc145細胞中進行。本研究采用脂質體介導方法,通過G418 篩選穩定轉染Marc145 細胞,并利用單個細胞克隆技術,獲得了穩定超表達CD151蛋白和(或)CD163 蛋白的Marc145 細胞株,并進一步探討其在疫苗生產中的應用,為最終獲得PRRSV高效增殖的新途徑提供依據。
1 材料與方法
1.1 質粒和細胞 去內毒素純化質粒p IRES2-EGFP-CD151 和p IRES2-EGFP-CD163 由本實驗室保存。Marc145 細胞由武漢中博生物股份有限公司提供,在本實驗室傳代保存。
1.2 試劑 脂質體2000 和G418,購自Invitrogen 公司;胎牛血清,購自Gibco 公司;DMEM培養基,購自清大天一公司;胰蛋白酶、PBS(PH7.4),購自北京全式金生物技術有限公司;RIPA裂解液、50*cooktail、PMSF(100 mmol/L)、磷酸化蛋白酶抑制劑、SDS-PAGE凝膠制備試劑盒、5*蛋白上樣緩沖液、ECL、顯影定影試劑,購自谷歌生物技術有限公司;BCA蛋白定量檢測試劑盒,購自Bio-rad 公司;蛋白Marker,購自Therm(Fermentas)公司;PVDF膜(0.22 μm、0.45 μm),購自Millipore 公司;BSA,購自Roche 公司;TWEEN 20,購自Amresco 公司;actin,購自Santa cruz 公司;GAPDH,購自Epitomics 公司;CD151、CD163 抗體,購自Abcam 公司。
1.3 穩定轉染細胞株的建立 待Marc145 細胞融合至70%~80%,按照Lipofectamine 2000 轉染試劑盒操作說明書進行操作。轉染48 h 后,開始用G418(300 μg/mL)篩選3 d,逐漸增加篩選濃度至絕大部分細胞死亡,改為800 μg/mL 維持篩選6周。將獲得細胞采用極限稀釋法接種于96 孔細胞培養板,交替使用常規培養基和G418(600 μg/mL)培養基培養單個細胞,至其長出單個克隆,挑選單個細胞克隆繼續培養至分別建立穩定轉染CD151、CD163 和CD151/CD163 共轉染的細胞株。
1.4 熒光免疫細胞化學分析 分別培養未轉染的Marc145 細胞和穩定轉染的細胞(CD151、CD163 和CD151/CD163 共轉染),制作細胞爬片,進行熒光免疫細胞化學分析。用PBS按一定比例稀釋好的一抗(CD163,Rabbit,1∶50;CD151,Rabbit,1∶50)覆蓋組織,4°C孵育過夜。用PBS洗滌3 次。滴加與一抗相應種屬的熒光二抗(cy3 標記山羊抗兔1∶100;cy3標記山羊抗小鼠1∶100),覆蓋組織,避光室溫孵育50 min。滴加DAPI 染液,避光室溫孵育10 min。用PBS洗滌。稍甩干后將有細胞的一面朝下用抗熒光淬滅封片劑將玻片封固在載玻片上,于倒置熒光顯微鏡下觀察并采集圖像。
1.5 Western Blot 分析 分別收集4 種細胞并提取總蛋白,將樣品加入點樣孔中,5%濃縮膠電泳,轉膜;5%的脫脂牛奶脫色;加一抗(CD163,Rabbit,1∶1 000;CD151,Rabbit,1∶1 000;Actin,Rabbit,1∶1 000),4 ℃過夜;再加二抗(1∶3 000),室溫下孵育30 min;膠片拍照。
1.6 細胞接種病毒 按照文獻[12]的方法,用細胞生長液(含體積濃度為8%的小牛血清的DMEM培養基)對超表達CD151、CD163 或CD151/CD163共表達的Marc145 細胞在培養瓶中進行擴繁培養,待細胞量足夠后,以0.25%胰蛋白酶消化細胞接種到轉瓶,置于轉瓶培養箱中繼續培養,轉速10 r/h。待細胞覆蓋率達到80%以上,移除細胞生長液,按照0.001MOI 劑量接種PRRS病毒Ch-1R株,加入病毒維持液(含體積濃度為2%的小牛血清的DMEM),在37 ℃下繼續培養。細胞病變達到80%以上時收獲病毒液,隨后補入不含病毒的維持液,12 h 后再收毒,反復收毒5 次。按Reed-Muench 法計算TCID50值。將收獲的病毒液置于2 ℃~8 ℃備用。
1.7 疫苗制備 將上述收獲的病毒液用100 kD截留分子量的超濾膜包濃縮10 倍,滅活劑與病毒液的體積比按1∶2 000 加入質量濃度為37% 甲醛溶液進行滅活后,再經凝膠層析柱Sepharose 4FF柱層析得到純化疫苗,按抗原和油佐劑1∶1 比例加入油佐劑,乳化配制成滅活疫苗(油佐劑的配制為94%(V/V)白油、6%(V/V)的Span-80、2%(g/V)硬脂酸鋁)。
1.8 疫苗效力檢驗 疫苗免疫接種:3 個試驗組分別將疫苗按2 mL/頭的劑量接種4~6 周仔豬10頭,同時設對照組(非免疫10 頭)。血清抗體檢測:在免疫前和免疫后28 d 采集豬血清,檢測對PRRS病毒中和抗體效價。攻毒保護試驗:免疫后28 d,用PRRS病毒(含量為105.5TCID50/mL 的病毒液)1∶10 稀釋后給每頭豬肌肉接種3 mL,繼續飼養觀察21 d,撲殺剖檢。
2 結果與分析
2.1 穩定轉染細胞系的建立 在熒光顯微鏡下穩定轉染CD151、CD163 和CD151/CD163 共轉 染的細胞均表達強烈的綠色熒光蛋白(見中插彩版圖1)。擴繁培養的細胞經微生物學檢測合格后,在液氮中進行保藏,建立了種子細胞庫和工作細胞庫。
2.2 病毒受體在細胞轉染前后Marc145 細胞中的表達情況 結果表明,無論是單一轉染還是共轉染,穩定轉染后的Marc145 細胞表面紅色熒光強度增加明顯,預示其表達CD151 或(和)CD163 的水平明顯提高(見中插彩版圖2)。
2.3 Western Blot 分析病毒受體在細胞轉染前后Marc145 細胞中的表達情況 利用Western Blot 檢測相關細胞表面受體蛋白CD151 和CD163 的表達情況(圖3)。結果顯示,跟對照組(未轉染組)相比,穩定轉染單一受體蛋白或CD151、CD163 共轉染細胞中目的蛋白表達水平均高于未轉染細胞組,共轉染組較單一轉染組同一目的蛋白水平略低。
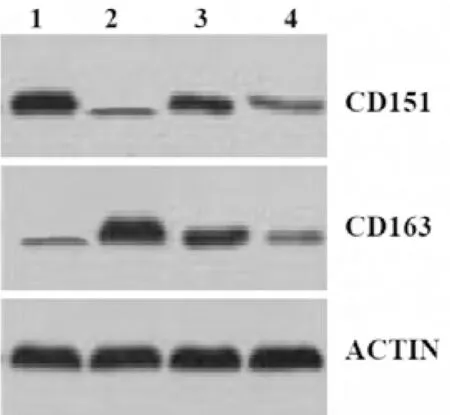
圖3 Western blot分析結果
表1 疫苗效力檢測結果
2.4 超表達受體蛋白的細胞制備疫苗效果分析利用超表達受體蛋白CD151 或(和)CD163 的Marc145 細胞接種PRRS病毒Ch-1R 株后,收獲病毒效價可達到108.0以上,較對照組(未轉染細胞組)提高10 倍以上。所制備的疫苗免疫接種動物后對PRRS病毒中和抗體效價與對照組(未轉染組)水平相當。攻毒保護試驗中通過剖檢未見病變情況,且疫苗吸收完全,達到部頒標準(1∶16)。
3 討論
PRRS病毒與細胞上的受體結合是實現其侵染靶細胞的先決條件。目前已經確定的PRRS病毒的細胞受體有6 種,分別是硫酸乙酰肝素(Heparin Sulphate,HS)、唾液酸粘附素(Sialoadhesin,Sn)、波形蛋白(Vimentin)、CD163(Cluster of Differentiation 163)分子、CD151(Cluster of Differentiation 151)分子和NMMHCIIA(非肌肉肌球蛋白重鏈ⅡA)分子。
已有多個研究小組報道,實驗室條件下通過構建超表達載體提高了病毒感染的滴度,但是尚未在疫苗制備上進行驗證。PRRSV非敏感細胞系BHK-21 和CRFK 在轉染重組Vimentin 后接種PRRSV,發現轉染Vimentin 的陽性細胞都能檢測到病毒RNA,而轉入半乳糖苷酶的陰性對照細胞中都檢測不到病毒RNA,可見PRRSV非敏感細胞由于Vimentin 的轉入獲得了易感性,表明Vimentin 可以 與PRRSV的N 蛋白結 合,是PRRSV受體復合物的一部分,Vimentin 可能與PRRSV感染Marc145 細胞有關。Welch 等報道轉染CD163 分子至PRRSV非敏感細胞系PK-15 中可以使此細胞獲得感染PRRSV和產生子代病毒粒子的能力。通過轉染使PRRSV非敏感細胞BHK-21 表達CD151 會使該細胞允許PRRSV的感染,增殖程度高達100 倍。最近有研究發現,將CD209 的基因導入BHK-21 細胞后能使BHK-21 細胞中產生的子病毒對Marc145 的感染能力增強。
本研究將CD151 和CD163 穩定轉染到Marc145細胞中,并通過其感染PRRS病毒Ch-1R(Ch-1a)株,實現了病毒培養滴度的顯著提高,最終在GMP 生產車間進行小規模試驗,所生產的不同類型疫苗均能達到部頒標準,為下一步將該技術更好地應用到生產實踐中去提供了依據。
[1]韓明遠,沈青春,張志剛,等.影響豬繁殖與呼吸綜合征病毒感染與增殖的因素[J].動物醫學進展,2010,31(1):87-91.
[2]蒲靜.PRRSV對豬肺泡巨噬細胞天然免疫功能影響的分子機制[D].北京:中國農業大學,2005.
[3]安同慶.豬繁殖與呼吸綜合征與宿主細胞受體之間相互作用的研究及病毒遺傳變異分析[D].北京:中國農業科學院,2007.
[4]蔣成蘭,邢鳳,徐云華,等.豬NMMHCⅡA基因功能區的克隆及表達特征分析[J].畜牧獸醫學報,2012,43(10):1519-1524.
[5]Shanmukhappa K,Kim JK,Kapil S.Role of CD151,Atetraspanin in porcine reproductive and respiratory syndrome virus infection[J].Virol J,2007,4:62.
[6]Welch SK,Calvert JG.Abrief reviewof CD163 and its role in PRRSVinfection[J].Virus Research,2010,154(1-2):98-103.
[7]Lee Y J,Park CK,Nam E,et al.Generation of a porcine alveolar macrophage cell line for the growth of porcine reproductiveand respiratory syndrome virus[J].Journal of Virological Methods,2010,163:410-415.
[8]De Vries W,Haasnoot J,van der Velden J,et al.Increased virus replication in mammalian cells by blocking intracellular innate defense responses [J].Gene Therapy,2008,15:545-552.
[9]薛慶善.體外培養的原理與技術[M].北京:科學出版社,2001.
[10]Kim JK,Fahad AM,Shanmukhappa K,et al.Defining the cellular target(s)of porcine reproductive and respiratory syndrome virus blocking monoclonal antibody 7G10[J].JVirol,2006,80(2):689-696.
[11]王偉偉,張璐,馬曉春,等.波形蛋白介導豬繁殖與呼吸綜合征病毒感染Marc145 細胞作用的初步研究[J].病毒學報,2011,27(5):456-461.
[12]Calvert JG,Slade DE,Shields SL,et al.CD163 expression confers susceptibility to porcine reproductive and respiratory syndrome viruses[J].JVirol,2007,81(14):7371-7379.
[13]Van Gorp H,Van Breedam W,Van Doorsselaere J,et al.Identification of the CD163 protein domains involved in infection of the porcine reproductive and respiratory syndrome virus[J].JVirol,2010,84(6):3101-3105.
[14]Lee Y J,Lee C.Deletion of the cytoplasmic domain of CD163 enhances porcine reproductive and respiratory syndrome virus replication[J].Arch Virol,2010,155(8):1319-1323.
[15]Huang Y W,Dryman BA,Li W,et al.Porcine DCSIGN:molecular cloning,gene structure,tissue distribution and binding characteristics[J].Dev Comp Immunol,2009,33(4):464-480.
