紅殼文蛤雙列雜交及雜種優勢分析*
2015-04-10 03:51:18吳楊平陳愛華姚國興張志偉蔡永祥
海洋與湖沼 2015年1期
關鍵詞:差異
吳楊平 陳愛華 張 雨 姚國興 張志偉 蔡永祥
(江蘇省海洋水產研究所 南通 226007)
雜交是創造變異的重要途徑, 是動植物遺傳改良的重要手段, 其主要作用是利用雜種優勢為育種制備中間材料(張國范等, 2004)。一個世紀以來, 雜種優勢一直是學術界關注的焦點之一, 主要原因在于其對生物產生的巨大作用、輔助的生物學機制及應用前景(張國范等, 2009)。采用雜交育種技術, 進而利用雜種優勢, 在動植物育種中已取得廣泛成功。海洋經濟貝類種類繁多, 是我國水產養殖產量最多的類群(約占70%)(李加琦, 2011)。在自然狀態下, 貝類的活動范圍較小, 因而比較容易形成具有不同遺傳特色的群體。因此, 貝類種內、種間雜交的研究較多。而隨著研究的不斷深入和擴大, 發現大多數種間雜交由于親緣關系相距較遠, 經常會出現雜交不親和、幼體不成活、雜種 F1不育、育性差和“瘋狂分離”等現象(張國范等, 2009)。盡管國內外對一些貝類研究過種間雜交, 但目前僅雜交蚌(李家樂等, 2007)等極少數種類成功應用; 而種內雜交卻不存在配子間的親和、發育障礙和育性等問題。因此, 自20世紀70年代以來, 貝類種內雜交的研究逐漸增多, 并且取得了較大進展(Newkirket al, 1977; Manziet al, 1991; Cruzet al, 1997; 張國范等, 2002; 常亞青等, 2002; 游偉偉等, 2010)。
文蛤(Meretrix meretrix)素有“天下第一鮮”之美譽,廣泛分布于我國各海區, 遼寧遼河口、山東萊州灣、江蘇南部沿海及廣西北部灣等資源最為豐富, 形成了我國文蛤的主產區。文蛤養殖歷史悠久, 其養殖方式逐漸由過去單一的自生自長、自然采捕的原始狀態發展成如今的移苗增殖、灘涂蓄水暫養、圍網精養、池塘混養等多種養殖方法, 但是不管養殖方式有多大的改變, 缺少高產抗逆品種一直掣肘著文蛤產業的發展。為此, 積極開展文蛤系統育種工作, 為產業發展注入新的活力是非常必要的。鑒于此, 本研究開展了不同文蛤地理群體間紅殼個體的雙列雜交, 旨在為文蛤雜交育種及雜種優勢在生產中的利用提供基礎資料。
1 材料與方法
1.1 親貝來源與3×3雙列雜交試驗設計
親貝分別采自江蘇如東(S)、廣西北海(G)和日本本州(J)。在位于呂四港的江蘇省海洋經濟貝類研發中心的試驗基地暫養促熟后用于家系構建。2013年 8月6—8日, 采用完全雙列雜交方式構建選育家系(見表1), 共成功構建 6組(Gg、Gs、Gj、Sg、Ss、Sj), 每組3個平行家系, 每個家系均由不同的1雌1雄文蛤配對繁殖產生。

表1 文蛤3×3雙列雜交試驗設計Tab.1 Design of diallel cross for Meretrix meretrix
1.2 雜交家系的構建及培育
采用陰干和流水刺激方法分別處理江蘇如東、廣西北海及日本本州文蛤群體中的紅殼個體, 親貝陰干12h后經流水刺激1h, 為防止意外受精, 將單個文蛤放入1L的燒杯中產卵和排精。分別收集卵子和精液后按照雙列雜交設計構建表1各組家系。
1.3 幼蟲及稚貝培育
將上述建立的各家系組轉移至300L的圓柱形塑料白缸中培育。經計數后受精卵密度調節為30ind/10ml。約24h后, D形幼蟲孵出, 將各家系幼蟲轉移至另一300L圓柱形塑料缸中培育, 初始密度控制在20ind/10ml, 保證各個家系的密度基本一致。早期每天半量換水1次, 培育水溫27.0—29.6°C; 以湛江叉鞭金藻(Dicrateria zhanjiangensis)、亞心形扁藻(Platymonas subcordiformis)和綠色巴夫藻(Pavlova viridis)為主要餌料, 連續充氣培養。附著后移苗至1000L的圓柱形塑料白缸中培育, 以自然海水為餌料來源, 適當補充人工單胞藻, 每天換水1次, 每次2/3。每3—5d沖洗培育過程中塑料白缸1次。上述培育過程中, 所用器具用砂濾海水浸泡后沖洗干凈, 保持各家系間的隔離,避免幼蟲交叉污染。
1.4 生長數據測量及雜種優勢計算
卵徑、幼蟲和殼長≤2000μm的稚貝均采用顯微鏡目微尺測量(目微尺精度±5μm), 殼長>2000μm的稚貝采用電子游標卡尺測量(精度±0.02mm), 每組均隨機取樣30個測量。日生長率的計算公式為:r(%)=[(Lt/L0)1/t?1]×100,Lt為結束時殼長(μm),L0為開始時殼長(μm),t為試驗天數(d)。
幼蟲存活率為單位體積幼蟲數與D形幼蟲數的百分比; 稚貝和幼貝階段的存活率為移池時收獲的數量與布入此池的數量的比值。
參照Zheng等(2011)使用的方法, 計算雜種優勢潛力指數hp(heterosis potence index,hp)和雜種獲得hg(hybrid gain,hg):hp=Q/L, 其中Q=2F1?(P1+P2),L=P1–P2;hg=[(F1?MP)/MP]×100, 式中,F1是雜種子代的平均值, MP是中親值: MP=(P1+P2)/2。當hp>1.0時,雜交產生顯著的雜種優勢; 當hp<–1.0時, 雜交產生雜交衰退; 當–1.0<hp<1.0時, 雜交效果不明顯, 雜交沒有產生顯著的雜交效果, 也沒有產生雜交衰退。
參照Cruz等(1997)和Lamkey(1998)等的計算公式,計算中親雜種優勢(mid-parent heterosis,HMP)和單親雜種優勢(single-parent heterosis,Hx):HMP=[(FC?P)/P]×100,HX=[(FC?PX)/PX] ×100, 式中FC是雜交組合(正交或反交)平均殼長,P為雙親自繁組的平均殼長,PX是某一親本自繁組的平均殼長。
參照曹景林等(2000)公式計算超親優勢Hph=[(F1?Pm)/Pm] ×100,F1為雜交子代某一性狀的平均值,Pm為雙親中某一性狀優良的親本的平均值。
參照Zhang等(2007), 采用普通線性模型分析母本效應(母本G和母本S)及交配方式(自繁、正交和反交)對殼長的影響: 模型如下:Yijk=μ+Pi+Mj+ (P×M)ij+eijk, 模型中,Yijk是來自第i個卵源j種交配方式下的第k個個體殼長, μ是常數項,Pi是卵源效應(母本效應)(i=1, 2),Mj是交配方式影響(j=1, 2, 3), (P×M)ij是卵源與交配方式的交互影響,eijk是隨機誤差。
1.5 數據處理
生長數據均以平均值±標準差(Mean±SD)表示,采用SPSS17.0統計軟件及EXCEL 2003對數據進行分析處理并作圖, 進行單因素方差分析(One-Way ANOVA)并采用 LSD多重比較各家系間殼長的差異,差異顯著性設置為P<0.05, 差異極顯著性設置為P<0.01。
2 結果與分析
2.1 浮游期、稚貝和幼貝階段的生長比較
構建成功的6個組合分別以江蘇文蛤、廣西文蛤為母本, 而兩者的卵徑存在顯著差異(P<0.05), 前者為 90±3.62μm, 后者為 77.05±7.05μm。因此, 卵源的大小差異造成受精卵的大小差異。以廣西文蛤為母本的 Gg、Gs和 Gj在 1日齡時 D形幼蟲大小在123.00—124.25μm, 而以江蘇文蛤為母本的 Sg、Ss和Sj在1日齡時D形幼蟲大小在141.25—143.75μm。1—5日齡的浮游期內, 以江蘇文蛤為母本的3個組合的殼長均顯著大于以廣西文蛤為母本的 3個組合(P<0.05)。表2反映了浮游期內各組的生長差異, 說明浮游期存在母本效應。從7月29日到8月2日的整個浮游期而言, 6個組合的日生長率存在大小差異,依次為 Gj>Gs>Sg>Sj>Gg>Ss(見表3)。由此可見, 雜交組合的日生長率大于自繁組合。
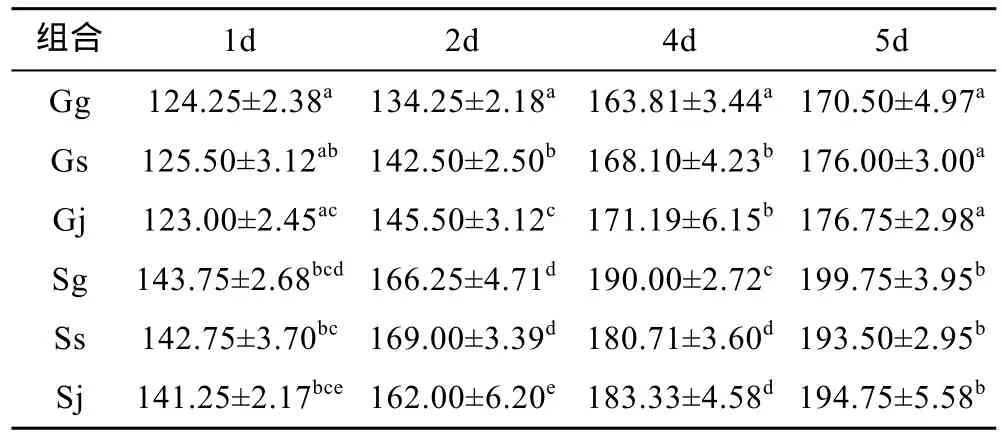
表2 浮游期的殼長比較Tab.2 Comparison of shell length during the pelagic period
13—43日齡的稚貝隨著日齡的增加, 生長差異逐漸顯現(圖1)。13日齡時, 在前期日生長率的累積作用下, 各組合殼長大小依次為: Gj>Sg>Sj>Gs>Gg>Ss, 雜交組合的殼長均大于自繁組合。可見, 來自先天卵源間的差異逐漸被后天生長所打破。43日齡時,各組合殼長大小依次為: Gs>Gj>Sg>Sj>Gg>Ss, Gg 和Ss差異不顯著(P>0.05), 與 Gs、Gj、Sg和 Sj均顯著(P<0.05)。從13—43日齡這一階段的日生長率來看(表3), Gs的日生長率最大, 其次分別為Sj、Gj和Sg, 最小分別為Ss和Gg。
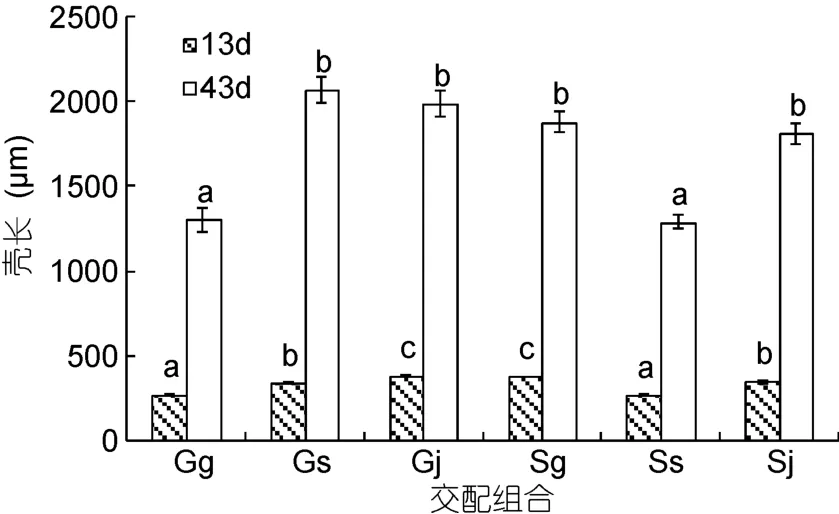
圖1 稚貝的殼長比較Fig.1 Comparison in shell length during the early juvenile period

圖2 幼貝的殼長比較Fig.2 Comparison in shell length during the juvenile period
圖2為73—149日齡的幼貝殼長比較。73日齡、104日齡和 149日齡的殼長大小依次為 Gs>Sj>Gj>Sg> Gg>Ss, Gs>Gj>Sj>Sg>Gg>Ss, Gs>Gj>Sj>Sg>Gg>Ss,可見, 在這3個時期Gs的殼長均最大, Gg和Ss依然最小。由表3的73—149日齡日生長率可見, 6個組合日生長率的大小依次為: Gs> Gj> Sg >Sj> Ss> Gg。1—149 日齡的日生長率大小依次為: Gs>Gj>Sj>Sg>Gg>Ss。因此, 雜交組合的生長快于自繁組。
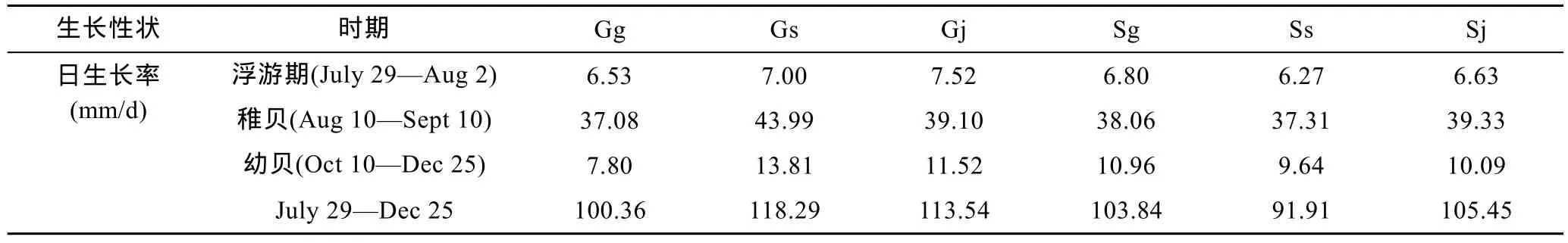
表3 各階段的日生長率比較Tab.3 Comparison of daily growth rates in different stages
由此可見, 在浮游期、稚貝和幼貝階段, 雜交組合(Gj、Gs、Sg、Sj)在日生長率方面均表現出比自繁組(Gg、Ss)快的生長優勢。浮游期受卵源因素的影響,尚未顯現出雜交組和自繁組殼長區別, 但在 13日齡以后的稚貝和幼貝階段, 雜交組比自繁組生長快的優勢迅速顯現, 并通過殼長明顯可以區別兩者的生長差異。由表4可見, 浮游期雜交組總體上未表現出生長優勢, 因其大小主要受卵源與交配方式的影響,此時兩者的交互作用對殼長無顯著影響(P>0.05); 其次是卵源的影響, 這時的大小差異主要來自卵源因素(P<0.01), 而交配方式未起到影響作用(P>0.05)。隨著日齡的增加, 交配方式對大小有顯著影響(P<0.01),表現出雜交組比自繁的生長優勢, 即雜種優勢。在整個 1—149日齡階段, 未見有卵源與交配方式對殼長的交互作用, 除4日齡、104日齡和149日齡分別對殼長有顯著影響(P<0.05)外, 均單獨對殼長有顯著影響(P<0.05)。因此, 殼長早期主要受到卵源因素的影響, 之后主要受到交配方式影響。

表4 卵源與交配方式對生長的影響Tab.4 Impacts of egg origination and mating strategy on shell growth
2.2 各日齡存活率的比較
各交配組合各個培育日齡的成活率見表5。D形幼蟲首日存活率設定為100%, 1—5日齡內幼蟲的存活率均在80%以上。5日齡時存活率大小順序為Gs>Sg>Gj>Sj>Gg>Ss, 相互間差異不顯著(P>0.05)。幼蟲變態附著后, 經倒換池重新布苗, 并將此時的稚貝存活率定義為 100%; 13日齡時, 各交配組合均保持100%的存活率。43—73日齡期間, 除自繁組Ss的存活率最低為 79.83%外, 其余存活率均在 80%以上,表現出較高的存活率。104—149日齡期間, 各組幼貝的存活率大小依次為 Gs>Sg>Gj>Sj>Gg>Ss。從 1—149日齡整個培育期間來看, 除 4日齡時 Gs的存活率低于其他交配組合, 但差異不顯著(P>0.05), 其余日齡Gs的存活率均最高; 且43日齡后, 雜交組的存活優勢更明顯。
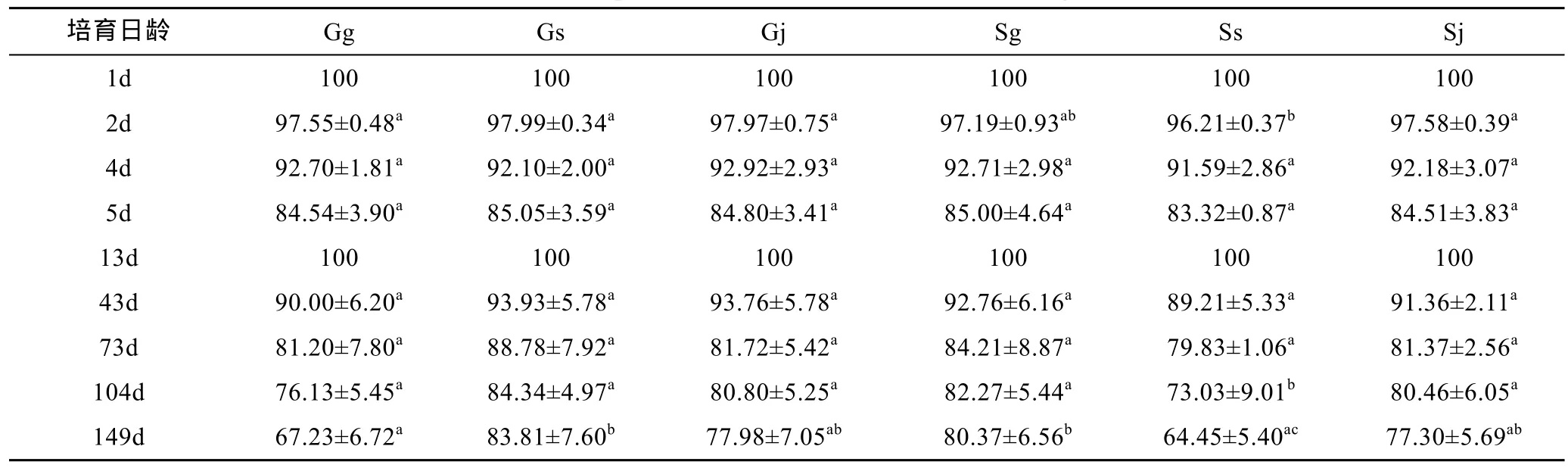
表5 各階段成活率比較Tab.5 Comparison in survival rates in different stages
2.3 雜種優勢比較分析
圖3中, 各雜交組殼長的雜種優勢整體上隨日齡的增大而增大。除Sg、Sj和Gj分別在2日齡、1—2日齡和1日齡為負值, 其余日齡均為正值; Gs在1—149日齡均呈正值, 且雜種優勢越來越明顯。由于只有Gs和Sg有雙親自繁組Gg和Ss, 因此只測算廣西群體G和江蘇群體S的中親雜交優勢HMP, 其值一直大于 0, 且始終介于單親雜種優勢HGs和HSg之間。如表6所示, 稚貝和幼貝階段, 中親雜種優勢HMP和單親雜種優勢HGs、HSj均顯著大于浮游期(P<0.05);單親雜種優勢HSg和HGj在浮游期、稚貝和幼貝這3個階段均有顯著差異(P<0.05), 其中稚貝階段>幼貝階段>浮游期。就整個發育階段而言, 稚貝階段的雜種優勢均大于幼貝階段, 浮游期最小。
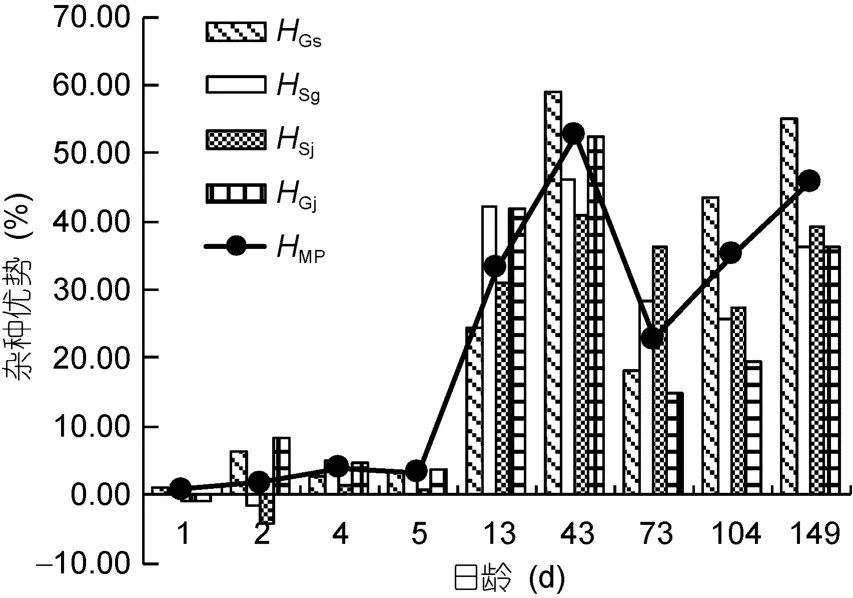
圖3 在不同日齡時的雜種優勢Fig.3 Heterosis shown on different days
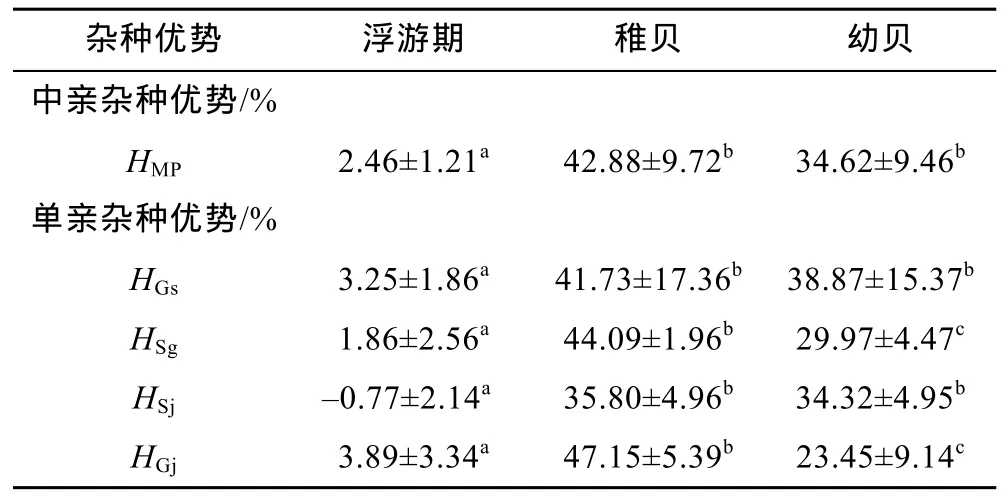
表6 不同發育階段的雜種優勢Tab.6 Heterosis shown in different stages
在1—149日齡, 只有早期1—5日齡的雜種優勢潛力指數為負值, 為–0.80—-0.12, 介于–1和 1之間,說明沒有雜種優勢產生, 且未出現負的雜種優勢, 未產生雜種衰退; 其余均為正值, 且均大于1, 說明有正的雜種優勢產生, 獲得了雜種優勢。圖4還可見, 殼長性狀的雜種獲得均為正值, 特別是稚貝以后殼長的雜種獲得都大于 22.80%, 說明殼長性狀得到了顯著改良。表7統計了Gs和Sg的超親優勢率, 13日齡之后Gs和Sg均有超親優勢率。

圖4 在不同日齡時的雜種優勢潛力指數及雜種獲得Fig.4 Potence index and hybrid gain on different days
2.4 子代的殼色表現
在自然界中, 廣西、江蘇和日本這3個群體中紅殼個體均只有1%—3%。3個群體的紅殼個體經雜交和自繁, 子代中紅殼個體的比例均有顯著提高, 數值介于72.80%—76.49%(表8)。說明紅殼可以得到一進步純化。

表7 不同發育階段的超親優勢率Tab.7 Super-parent heterosis shown in different stages
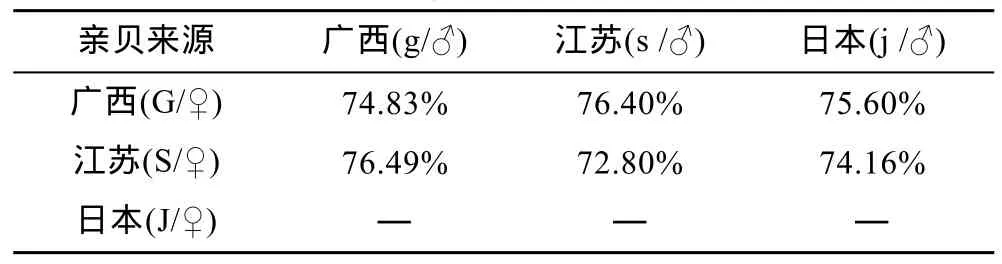
表8 雜交、自繁子代的殼色表現Tab.8 Shell color between hybrid and self-fertilized generations
3 討論
3.1 母本效應
母本效應對動物早期發育階段的生長是一個非常重要的影響因素。個體在其生長發育的早期階段,比較容易受到母本效應的影響, 而后母本效應減弱或消失, 雜種優勢得以充分表達。牡蠣的種間雜交、海灣扇貝(Argopecten irradians)的群體雜交、菲律賓蛤仔(Ruditapes philippinarum)的雜交等研究中均有類似發現(Newkirket al, 1977; 閆喜武等, 2008; 張國范等, 2009)。大量研究認為, 在種間或同種的不同群體間雜交時, 雜交個體在不同環境下的遺傳變異程度很難確定, 由于不同種間、群體間個體的營養積累程度、卵徑大小、攜帶的卵內營養物質多少均不同,故在胚胎發育早期表現顯著的母本效應。本試驗中,1—5日齡浮游期內各組合的生長主要受到母本效應影響。造成這種情況的原因在于母本來自不同群體,且母本間存在著顯著的卵徑大小差異。卵徑大小的差異反映出卵所儲存的能量存在差異, 而這些能量恰恰是幼蟲早期發育階段的主要營養來源(Cruzet al,1997), 這可能是本試驗中產生母本效應的原因。
3.2 雜種優勢
本研究表明, 文蛤各雜交組的雜種優勢在各個生長階段所有差異。其他研究也表明不同生長階段雜種優勢存在差異, 如閆喜武等(2008)在菲律賓蛤仔不同殼色品系間雜交中發現, 在不同階段雜種優勢的平均水平有所不同; Cruz等(1997)發現兩個扇貝群體雜交幼蟲在0—11日齡生長的雜種優勢為0, 17日齡時卻能達到6.8%。此外, 本研究表明正交組Gs雜種優勢與反交組Sg存在著明顯差異。在不同貝類群體、品系間進行雜交研究普遍都認識到正反交組合的雜種優勢有差異, 如海灣扇貝、長牡蠣(Crassostreagigas)、美洲牡蠣(Crassostrea virginica)等中這種差異是一種比較常見的現象(Malletet al, 1984; Hedgcocket al, 1995; Cruzet al, 1997)。
為了有效評估雜種優勢, 在本研究的完全2×2雙列雜交中引入了雜種潛力指數這一概念(Zhenget al,2010)。這種判斷方法先后在評定亞種間、種間雜交時具有高度適用性, 取得了較為理想的評估效果(張國范等, 2009)。雜種潛力指數可作為判斷雜交是否真正產生雜種優勢的依據, 因為只有當雜種子代均值超過最大親本值時, 雜種潛力指數才大于1.0(張國范等, 2009)。紅殼文蛤不同地理群體間的雜交, 雜種獲得均為正值, 特別是稚貝以后, 殼長的雜種獲得顯著高于22.80%。73日齡時, 雜交優勢率較低, 造成此結果的原因在于自繁組的生長增加較快, 大于同期雜交組的生長增幅; 但 73日齡之后的各日齡, 雜交組的生長快于自繁組, 雜種優勢明顯。73日齡時的雜交優勢率波動的具體原因還有待進一步研究分析, 但雜交組的雜種優勢確實存在, 這種正的雜種優勢潛力指數及雜種獲得應歸因于這兩個不同地理群體間存在著一定的遺傳差異, 林志華等(2008)利用 AFLP等分析結果表明廣西文蛤和江蘇文蛤確實存在較大的遺傳差異。總之, 本研究的結果充分表明紅殼文蛤不同地理群體雜交能獲得正的雜種優勢, 雜交組在生長速度方面優于對照組, 說明不同地理群體間的雜交可能是文蛤遺傳改良的一條有效途徑。
閆喜武, 張躍環, 霍忠明等, 2008.不同殼色菲律賓蛤仔品系間的雙列雜交.水產學報, 32(6): 864—875
李加琦, 2011.皺紋盤鮑配套雜交體系的建立、評價及應用. 青島: 中國科學院海洋研究所博士學位論文
李家樂, 白志毅, 2007.淡水養殖新品種——康樂蚌. 中國水產, (10): 44—45
張國范, 劉曉, 闕華勇等, 2004.貝類雜交及雜種優勢理論和技術研究進展. 海洋科學, 28(7): 54—60
張國范, 王繼紅, 趙洪恩等, 2002.皺紋盤鮑中國群體和日本群體的自交與雜交F1的RAPD標記. 海洋與湖沼, 33(5):484—491
張國范, 鄭懷平, 2009.海灣扇貝養殖遺傳學. 北京: 科學出版社, 76—102
林志華, 董迎輝, 李寧等, 2008.基于形態參數和 AFLP標記的文蛤(Meretrix meretrix)不同地理群體遺傳變異分析.海洋與湖沼, 39(3): 245—251
常亞青, 劉小林, 相建海等, 2002.櫛孔扇貝中國種群與日本種群雜交子一代的早期生長發育. 水產學報, 26(5):385—390
曹景林, 汪周, 2000.烤煙雜種一代不同生育時期株高和葉數的優勢. 安徽農業技術師范學院學報, 14(3): 23—26
游偉偉, 駱軒, 王德祥等, 2010. “東優1號”雜色鮑及其親本群體的形態特征和養殖性能比較. 水產學報, 34(12):1837—1843
Cruz P, Ibarra A M, 1997. Larval growth and survival of two catarina scallop(Argopecten circularis) populations and their reciprocal crosses. Journal of Experimental Marine Biology and Ecology, 212(1): 95—110
Hedgcock D, McGoldrick D J, Bayne B L, 1995. Hybrid vigor in pacific oysters: an experimental approach using crosses among inbred lines. Aquaculture, 137(1—4): 285—298
Lamkey K R. Edwards J W, 1998. Heterosis: theory and estimation. Proceediongs 34th Illinois Corn Breeders’School ,Urbana, IL, 2-3 Mar 1998, 62—77
Mallet A L, Haley L E,1984. General and specific combining abilities of larval and juvenile growth and viability setimated from narural oyster populations. Marine Biology,81(1):53—59
Manzi J J, Hadley N H, Dillon R T, 1991. Hard clam,Mercenaria mercenaria, broodstocks: growth of selected hatchery stocks and their reciprocal crosses. Aquaculture, 94(1):17—26
Newkirk G F, Waugh D L, Haley L E, 1977. Genetic of larval tolerance to reduced salinities in two populations of oysters,Crassostrea virginica. Journal of the Fisheries Research Board of Canada, 34:383—387
Zhang H, Liu X, Zhang Get al, 2007. Growth and survival of reciprocal crosses between two bay scallops,Argopecten irradiansconcentricus Say and A. irradians irradians Lamarck.Aquaculture, 272(S1): 88—93
Zheng H P, Xu F, Zhang G F, 2011. Crosses between two subspecies of bay scallopArgopecten irradiansand heterosis for yield traits at harvest. Aquaculture Research, 42(4):602—612
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
