光照強度對狐臭柴葉片形態與結構的影響
2015-07-01 08:02:00劉詩雅張明生高曉峰劉貴賢韋紅邊
貴州農業科學 2015年6期
劉詩雅,張明生,高曉峰,劉貴賢,韋紅邊,呂 享
(貴州大學 生命科學學院,貴州 貴陽550025)
狐臭柴(PremnapuberulaPamp.)系馬鞭草科豆腐柴屬多年生落葉灌木,又名神仙豆腐柴,分布于我國貴州、四川、廣西、湖北、湖南和陜西等省區海拔700~1 800 m 山坡叢林中[1]。狐臭柴是一種藥食兼用植物,其葉能清濕熱解毒、治水腫毒瘡,根具有調經壯陽、治月經不調、風濕關節炎和陽痿等功效[2-3];且葉片富含果膠、蛋白質、氨基酸和纖維素等營養成分[4-5],民間用葉揉碎制作清涼可口、色香味俱全的“神仙豆腐”,深受人們的喜愛,具有較大的經濟利用價值。狐臭柴因其自然繁殖能力較差[4],加之生態環境被破壞,該物種野生資源日益減少,分布零散,采集困難[6],故開展其人工種植十分必要。狐臭柴葉片是主要的食用部位,作為光合作用的主要器官,是產生同化物的重要場所,直接影響植株生物量的形成[7]。光是植物生長的主要能源,也是影響植株葉片形態、解剖結構、生理代謝及生長發育的主要因子之一[8-13]。而目前尚無有關光照對狐臭柴葉片形態結構產生影響的研究報道。對此,筆者通過設置不同光照強度,探索其對狐臭柴葉片形態和解剖結構的影響,以期為狐臭柴人工種植技術的構建與開發利用提供理論依據。
1 材料與方法
1.1 材料
1.1.1 試驗植物 狐臭柴,來源于貴州省務川仡佬族苗族自治縣,由貴州大學鑒定。
1.1.2 遮陽網 不同目數的黑色遮陽網,市售。
1.2 試驗設計
以不同目數的黑色遮陽網調節光照強度,分別設重度遮光、中度遮光和不遮光3種光照強度處理。處理1,重度遮光,透光率5%~10%;處理2,中度遮光,透光率40%~50%;處理3,不遮光為對照(CK)。使用85%的壤土混合15%的沙土作為栽培基質,以排水良好的塑料桶(桶高32cm,上口徑35cm,下口徑32cm)栽植狐臭柴一年生扦插苗,裝土深度為27cm,每桶栽植1 株,每個處理栽植36桶。
1.3 葉片形態指標測定
于7月初采摘狐臭柴植株一級枝條健康的倒三葉,用FAA 進行固定,石蠟切片法制片,切片厚度10μm。用OLYMPUS-BX40顯微鏡在20倍和40倍物鏡下觀察并攝像,使用Photoshop 7.0以20倍和40倍物鏡下的顯微測微標尺為標準,對葉片厚度、葉脈直徑、柵欄組織厚度、海綿組織厚度、上下表皮厚度及上下表皮細胞密度等進行統計(表皮裝片為撕取上、下表皮制作的臨時裝片),葉片形態指標按照植物學常規描述。
1.4 數據處理
以Excel和DPS統計軟件進行數據分析。
2 結果與分析
2.1 光照強度對狐臭柴葉片形態的影響
從表1和圖1看出,隨著光照強度的增加,葉片表面積(葉片大小)逐漸減小;葉形由心形、近心形過渡到卵形;葉緣由鋸齒、部分鋸齒到全緣;葉尖由長尾尖、尾尖到急尖;葉色由深綠色、綠色到淺綠色;葉質由紙質逐漸變為堅紙質。說明,光照強度是狐臭柴生長的重要環境因子,蔭蔽條件下葉面積增大(包括鋸齒、心形、長尾尖)、葉色變深(葉綠素含量增加)、葉質變軟(角質層減少)等均是狐臭柴植株對陰生環境的良好適應。
2.2 不同光照強度下狐臭柴葉片的解剖結構
狐臭柴葉片上、下表皮各有一層細胞,細胞大小不等,形狀不規則,排列緊密,上表皮氣孔極少,下表皮氣孔較多,上、下表皮均有表皮毛和腺體(圖2~3)。遮光處理下葉片的上表皮細胞形態與對照差異較大(圖2-A~C),而下表皮細胞形態在不同處理間無明顯差異(圖2-D~F)。
不同光照強度下狐臭柴葉片組織結構有不同程度的變化(表2,圖3~4)。上表皮細胞密度、葉脈直徑隨光照強度的增加呈極顯著正相關,而下表皮細胞密度受光照強度變化的影響不明顯;葉片總厚度、上表皮厚度、柵欄組織厚度和海綿組織厚度在2種遮光條件下變化不大,但與對照均有極顯著差異,而葉片下表皮厚度受光照強度的影響很小。狐臭柴是典型的陰生植物,強光條件下,上表皮變厚且細胞密度增大、氣孔極度減少,有利于降低水分蒸騰;柵欄組織和海綿組織增厚,有助于充分回收光呼吸釋放的胞間CO2;葉脈變粗,可保障光合產物的瞬時轉運,有效地降低光呼吸消耗。

表1 不同光照強度下狐臭柴葉片的形態指標變化Table 1 Variation of leaf morphology indices under different sunlight intensity

圖1 不同光照強度下狐臭柴葉片的形態變化Fig.1 Leaf shape changes under different sunlight intensity

表2 不同光照強度下狐臭柴葉片的組織結構變化Table 2 Changes of leaf tissue structure under different sunlight intensity

圖2 不同光照強度下狐臭柴葉片的表皮細胞形態Fig.2 Leaf epidermal cell morphology under different sunlight intensity
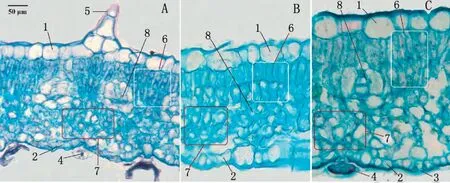
圖3 不同光照強度下狐臭柴葉片的解剖結構(橫切)Fig.3 Leaf anatomical structure under different sunlight intensity(transection)
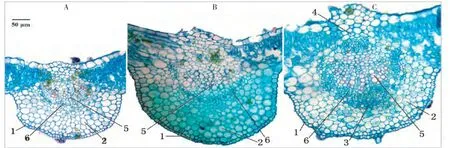
圖4 不同光照強度下狐臭柴葉脈的解剖結構(橫切)Fig.4 Leaf vein anatomical structure under different sunlight intensity(transection)
3 小結與討論
1)研究結果表明,隨著光照強度的增加,狐臭柴葉片表面積(葉片大小)逐漸減小;葉形由心形、近心形過渡到卵形;葉緣由鋸齒、部分鋸齒到全緣;葉尖由長尾尖、尾尖到急尖;葉色由深綠色、綠色到淺綠色;葉質由紙質逐漸變為堅紙質。表明,狐臭柴屬于陰生植物的特征。
2)地球上的每一種植物,在長期進化過程中形成了適應于特定自然環境、有利于自身生長發育的形態結構,而且這些特定的形態結構會隨著環境的變化而發生一定的改變,這就是植物的生態適應性。本研究所用的狐臭柴屬于陰生植物,光照強度是影響其生長發育的重要生態因子,強光下,其葉面積減小,葉片上表皮角質增加、細胞密度增大、柵欄組織和海綿組織增厚,這些變化充分反映了狐臭柴對光逆境的適應性,也符合大多數植物在高光強下葉片解剖結構的變化[14-18]。然而,植物的逆境適應能力有限,例如,適度的強光照可促進葉片表面角質層的形成,進而提高葉片反光度以減輕光對葉肉細胞的灼傷,同時通過薄膜干涉和改變光反射方式影響光量子吸收[19];若光照過強,則使葉綠體變小、類囊體結構破壞,光合作用強烈受阻而導致植株死亡[20-23]。
[1]中國科學院植物研究所.中國高等植物圖鑒[M].北京:科學出版社,1985:87-88.
[2]中國科學院中國植物志編輯委員會.中國植物志:第65卷[M].北京:科學出版社,1982:86-87.
[3]徐 利,史夢娜,張明生.狐臭柴扦插繁殖技術初步研究[J].山地農業生物學報,2011,30(1):82-86.
[4]史夢娜,張明生,徐 利,等.溶液培養對狐臭柴植株生長及葉片神仙豆腐的影響[J].中南林業科技大學學報,2012,32(03):70-74.
[5]Zhang M S,Shi M N,Xu B R,et al.A key component determination on forming Fairy Tofu from the leaf of Premna Puberula(Verbenaceae)[J].Journal of Applied Pharmaceutical Science,2012,2(9):31-35.
[6]許邦銳,張明生,李祥棟,等.光照強度對狐臭柴植株生長及葉片生理生化指標的影響[J].中南林業科技大學學報,2013,33(12):82-85,95
[7]趙立華,胡中會,李成云,等.不同遮光程度對玉米葉片結構的影響[J].中國 農 學通報,2012,28(6):43-46.
[8]Philip M P,Fraser J G M.Effects of shade on growth biomass allocation and leaf morphology in European yew (Taxus baccata L.)[J].European Journal of Forest Research,2013,132:211-218.
[9]Zhang S B,Yin L X.Plasticity in photosynthesis and functional leaf traits of Meconopsis horridula var.racemosa in response to growth irradiance[J].Botani-cal Studies,2012,53:335-343.
[10]Lusk C,Reich P B,Montgomery R A,et al.Why are evergreen leaves so contrary about shade[J].Trends in Ecology and Evolution,2008,23:299-303.
[11]許邦銳,張明生,李祥棟,等.光照強度對狐臭柴植株生長及葉片生理生化指標的影響[J].中南林業科技大學學報,2013,33(12):82-85,95.
[12]Chen J W,Zhang Q,Li X S,et al.Steady and dynamic photosynthetic responses of seedlings from contrasting successional groups under low-light growth conditions[J].Physiologia Plantarum,2011,141:84-95.
[13]李芳蘭,包維楷.植物葉片形態解剖結構對環境變化的影響與適應[J].植物學通報,2005,22(增刊):118-127.
[14]江 莎,胡 陽,鄭書馨,等.不同光強與光質對“達賽萊克特”草莓葉片形態結構的影響[J].電子顯微學報,2009,28(5):453-461.
[15]史剛榮,蔡慶生.白三葉葉片解剖可塑性及其對光強的響應[J].草地學報,2006,14(4):301-305.
[16]Ashton P M S,Berlyn G P.Leaf adaptation of some Sharea species to sun and shade[J].New Phytologist,1992,121:587-596.
[17]Ashton P M S,Berlyn G P.A comparison of leaf physiology and anatomy of Quercus(Section Erythrobalanus-Fagaceae)species indifferent light environments[J].American Journal of Botany,1994,81(5):589-597.
[18]Crystalarens N.Response of leaf anatomy to light environments in the tree fem Cyahtea caracasana(Cyatheaceae)and its application to some ancient seed fem[J].Palaios,1997,12:84-94.
[19]Wolfgang B,Olle B,Susan S T.Light-induced spectral absorbance changes in relation to photosynthesis and the epoxidation state of xanthophyll cycle components in cotton leaves[J].Plant Physiology,1989,91:542-551.
[20]Tiffany A M,Mark H E,Casey W B,et al.Differences in chloroplast ultrastructure of Phaeocystis antarctica in low and high light[J].Marine Biology,2006,149:1281-1290.
[21]Wheeler W S,Fagerberg W R.Exposure to low levels photosynthetically active radiation induces rapid increases in palisade cell chloroplast volume and thylakoid surface area in sunflower(Helianthus annuus L.)[J].Protoplasma,2000,212:38-45.
[22]王菊鳳,李鵠嗚,廖飛勇,等.光照對盾葉薯蕷熒光光譜和葉綠體結構的影響[J].吉首大學學報,2006,27(1):80-85.
[23]陳章和,朱素琴,李韶山,等.UV-B 輻射對東南亞熱帶森林木本植物幼苗生長的影響[J].云南植物研究,2000(22):467-472.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
哲學評論(2021年2期)2021-08-22 01:53:34
當代陜西(2021年2期)2021-03-29 07:41:24
中華詩詞(2019年7期)2019-11-25 01:43:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
