水稻抽穗期基因Hd7m的定位及遺傳分析
2015-07-18 15:02:40曾蓮張華柳絮等
山東農業科學 2015年1期
關鍵詞:水稻
曾蓮 張華 柳絮等

摘要:以紅旗16為受體、明恢63為供體,經雜交和回交得到BC4F1。利用643對SSR分子標記對所構建的抽穗期基因早晚池和親本進行多態性篩選,得到4對多態性標記。將篩選出的雜合單株進行自交,獲得BC4F2,并從中篩選出抽穗期分離較明顯的群體進行田間表型調查和基因型檢測。發現目標基因與標記PSM391連鎖,位于第7染色體長臂末端,命名為Hd7m。在標記PSM391與第7染色體末端之間合成10對新SSR標記,其中多態性標記RM22156與目標基因相距4.1 cM。該結果為Hd7m基因的精細定位、基因克隆和分子標記輔助育種奠定了基礎。
關鍵詞:水稻;抽穗期;基因定位;SSR標記;遺傳分析
中圖分類號:S511:Q785 文獻標識號:A 文章編號:1001-4942(2015)01-0010-04
Abstract Using Hongqi 16 as the receptor and Minghui 63 as the donor, a BC4F1 segregation population was developed through cross between the two parents and backcross with Hongqi 16. The early heading gene pool, late heading gene pool, and the parents were analyzed with 643 pairs of SSR markers, and 4 pairs of polymorphic markers were obtained. BC4F2 populations derived from inbreeding of BC4F1 hybrid plants with obvious segregation phenotype were selected for field investigation and genotype detection. The results showed that the target gene, named Hd7m, was linked to SSR marker PSM391, which located at the end of chromosome 7. Based on the targeted interval, 10 pairs of new SSR markers were synthesized. Among which, the polymorphic marker RM22156 was apart from Hd7m of 4.1 cM. The results laid foundations for fine mapping, gene cloning and molecular marker assisted breeding of Hd7m.
Key words Rice; Heading date; Gene mapping; SSR marker; Genetic analysis
水稻是短日照植物,它的抽穗是通過光周期來調整的。抽穗期是水稻適應環境變化最基本的農藝性狀之一。水稻基因間互作及基因和環境間的互作使抽穗期的遺傳行為十分復雜。水稻復雜性狀遺傳學基礎的分類,不僅對水稻發展的基礎研究方面有很大的影響,對水稻育種也有實際的價值。隨著分子標記技術和基因克隆技術的應用,人們對水稻抽穗期遺傳機制的研究也逐漸深入,國內外定位了700多個抽穗期QTL(http//www.gramene.org/qtl/index.html),分布在水稻12條染色體上,其中第3、6、7染色體上較多,第10染色體上較少。存在于不同位點上的QTL被檢測到的概率取決于自身的效應大小,效應較大的QTL被檢測到的概率較大[1]。
日本水稻基因組計劃(Rice Genome Research Program, RGP)Yano團隊深入研究水稻的QTL,他們用秈粳交組合(Nipponbare/Kasalath)衍生的F2、回交重組自交系(Backcross recombinant inbred line,BIL)和高代回交群體定位了15個水稻抽穗期QTL:Hd1~Hd3a、Hd3b~Hd14[2]。其中,利用日本晴和Kasalath雜種F2代的186個植株和850多個分子標記對影響水稻抽穗期的QTL進行定位,發現2個主效QTLs(Hd1和Hd2)和3個微效QTLs(Hd3、Hd4和Hd5)。其中主效Hd1位于第6染色體中部,另一個基因Hd2位于第7染色體末端[3]。利用日本晴作輪回親本,Kasalath作供體,分別構建包含Hd1、Hd2和Hd3的近等基因系,并借助RFLP標記對這3個QTLs進行精細定位。其中,Hd1定位于水稻第6染色體RFLP標記R1679和P130之間,并與C235共分離[1]。在Hd1和Hd2座位上,Kasalath等位基因均促進抽穗,在Hd3、Hd4和Hd5座位上,Kasalath等位基因均推遲抽穗。分析表明,通過移除主效基因對表型的影響可以促進微效基因的表達。在BC1F5群體中定位了5個抽穗期QTLs,其中效應較大的2個,分別與Hd1和Hd2處于同一區間,另外3個QTLs(Hd7、Hd8和Hd11)的效應較小[4,5]。以Nipponbare為輪回親本,對Nipponbare/Kasalath組合進行多次回交和篩選,得到高世代回交群體,用上述遺傳背景較單一的回交群體,又檢測到兩個新的抽穗期QTLs:Hd6和Hd9,兩QTL的Kasalath等位基因都推遲水稻抽穗[6,7]。用上述高世代回交群體又定位了Hd4和Hd5,Hd4被定位在第7染色體上,在標記R46和C39最近的區域之間;Hd5被定位在第8染色體短臂C166和R902之間,兩個QTLs均為單一的孟德爾因子。Hd5與Hd2在調節抽穗期上表現為加性效應,表明Hd2與Hd5處在不同的光周期途徑中。光周期敏感因子Hd3作為與水稻抽穗期相關的QTL最初被定位于第6染色體上,為了驗證Hd3的基因型,在長日照和短日照兩種條件下分析BC4F2群體Hd3區段中20個重組單株的抽穗期,發現該區段包含兩個影響抽穗期的基因,分別命名為Hd3a和Hd3b[8]。后來,在Nippobare/Kasalath衍生的高世代群體中又鑒定到了Hd10、Hd12、Hd13和Hd14共4個影響水稻抽穗期的QTLs[9]。endprint
近年來,國內外在精細定位的基礎上克隆了大量與水稻抽穗期相關的基因,如Hd1、Hd2、Hd3a、Hd3b、RFT1、DTH8等,并研究它們對抽穗期調控的相互作用,為國內外學者們對水稻抽穗期的相關研究提供了重要的參考依據。本試驗所用的材料BC4F2為高世代回交重組自交群體,其抽穗期性狀穩定遺傳,受1對等位基因(Hd7m)控制。本研究為Hd7m的精細定位、分子標記輔助育種和基因克隆提供了重要的依據。
1 材料與方法
1.1 試驗材料
以紅旗16為受體親本,以明恢63為供體親本,雜交得F1,以紅旗16為回交親本得BC4F1并收取種子。將回交得到的種子自交得BC4F2,從中選出抽穗期分離較明顯的群體。
1.2 試驗方法
1.2.1 材料種植 試驗在山東省農業科學院生物技術研究中心飲馬泉農場進行,2013年4月20日播種、6月2日插秧,同年篩選到抽穗期分離明顯的群體,編號R185。選擇R185中基因型均表現為雜合的單株種子,于2014年4月28日播種、6月1日插秧,同年篩選出抽穗期分離明顯的群體,編號LL。
1.2.2 田間抽穗期調查 每3天進行一次田間調查,以單株的第一個穗子抽出1 cm作為抽穗日,以播種日至抽穗日的天數作為抽穗期,對親本、回交群體每個單株進行抽穗期記載。
1.2.3 水稻DNA的提取及SSR分子標記分析 用TPS法提取水稻DNA備用。用BSA法[10]分別構建抽穗期DNA早晚池。根據網上(http: // www.gramene.org)已發表的標記序列合成SSR標記。PCR擴增按照Panaud等(1996)的方法[11]進行部分修改后使用。配置20.5 μL反應體系,包括2 μL DNA模板、0.1 μL dNTP、2 μL 10×buffer、13.2 μL ddH2O、0.2 μL Taq酶、3 μL 10 μmol/L引物。PCR儀擴增程序為94℃預變性5 min;94℃ 1 min,55℃ 1 min,72℃ 1 min,35個循環;72℃延伸10 min。PCR產物經6%變性聚丙烯酰胺凝膠電泳及銀染顯色,記錄單株帶型結果,拍照保存。
1.2.4 用早熟單株定位Hd7m在染色體上的位置 結合田間表型對基因型進行分析。用篩選出的4對多態性標記對早熟的97個單株逐個進行基因型檢測,計算分離群體的重組率,C=(N1+N2/2)/N,其中N為早熟純合基因型的總株數,N1為晚熟純合基因型的總株數,N2為雜合基因型的總株數[12]。用MapDraw對目標基因進行連鎖分析。
2 結果與分析
2.1 分離群體的表型及遺傳分析
明恢63抽穗期為114天,紅旗16抽穗期為120天。選用次級分離群體中抽穗期分離較明顯的群體LL,其抽穗期在95~143天,呈雙峰分布(圖1),以107天為分界。卡方分析表明,LL早抽穗植株和晚抽穗植株數目的分離比符合1∶3 (χ2=0.72<χ20.05,1=3.84,表1)[2]。因此,認為分離群體的抽穗期受1對等位基因控制,且晚抽穗對早抽穗表現為完全顯性。
2.2 目標基因的定位
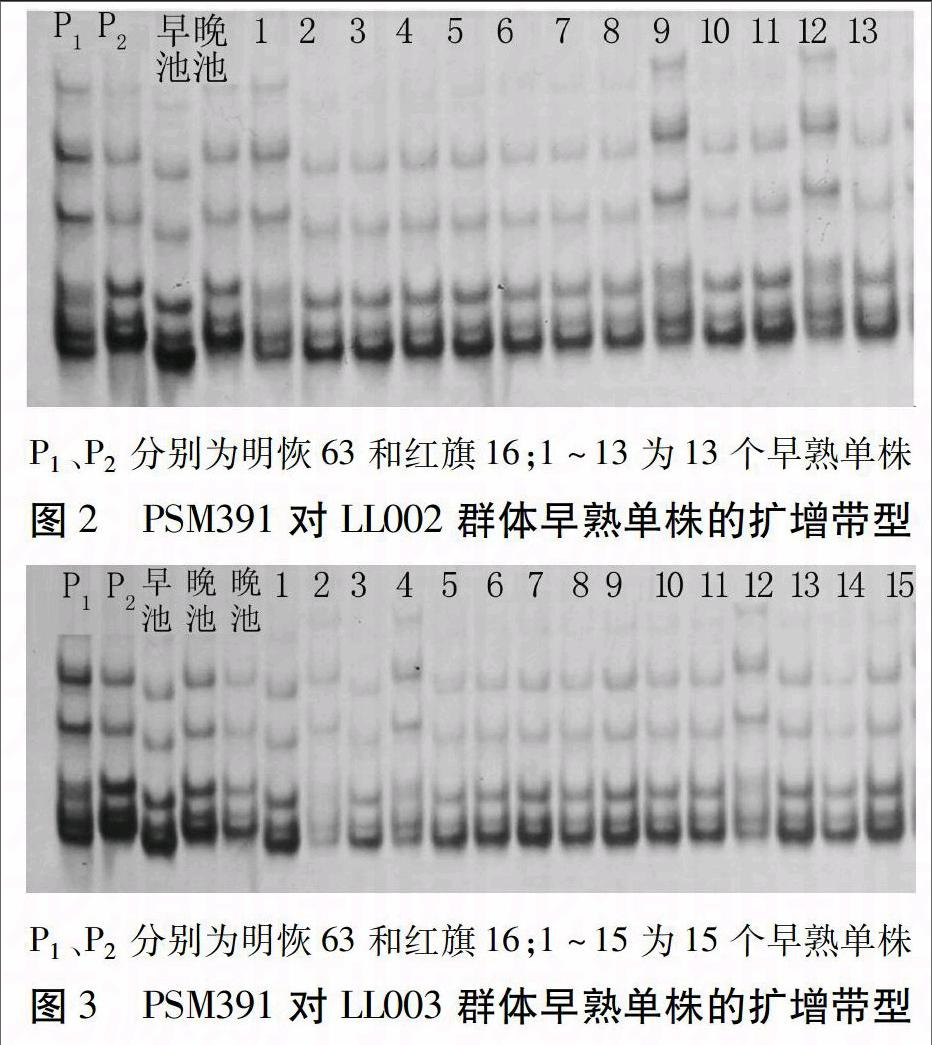
利用合成的643對SSR標記對親本及所構建的基因早晚池進行多態性篩選,得到多態性標記PSM353、RM505、RM18和PSM391。因LL群體中晚熟植株數量較大且抽穗期較分散,因此,選用早熟的97個單株作為定位群體,用4對標記對LL群體單株進行基因型驗證(以LL002和LL003群體為例,圖2、3)。經重組率計算,目標基因離4對標記的遺傳距離分別為38、34、26.8 cM和13.4 cM。根據物理圖譜和田間表型分析,目標基因位于PSM391與第7染色體末端之間,命名為Hd7m。
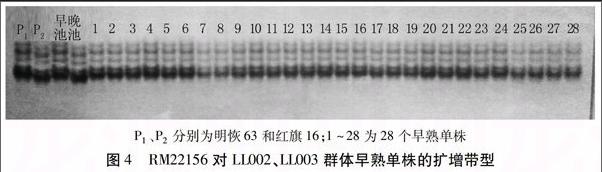
為了能將Hd7m基因限定在更小的范圍內,在標記PSM391與第7染色體末端區域間合成了10對新標記,對親本及基因早晚池進行多態性篩選,得到多態性標記RM22156。用標記RM22156對LL分離群體中97個早熟單株進行驗證(圖4)。通過重組率計算,Hd7m與標記RM22156相距4.1 cM(圖5)。
3 討論與結論
抽穗期是影響水稻品種栽培地區和栽培季節的重要農藝性狀,對水稻抽穗期基因進行精細定位,有助于抽穗期基因克隆和分子標記輔助育種。目前,國內外學者對不同的水稻品種進行了抽穗期QTL的定位與克隆的研究,通過構建不同QTL的近等基因系,對Hd1、Hd2、Hd3、Hd6和Hd9進行了精細定位分析,其中Hd2定位于第7染色體末端,距RFLP標記C586為0.5 cM[6,13,14],是OsPRR37的同源基因[15,16],OsPRR37的位置為29617430-29628600 bp[2]。本試驗定位的Hd7m與第7染色體標記RM22156相距4.1 cM,根據網站(http: //www.gramene.org)得到RM22156的位置為29146343-29146365 bp(C586的位置為29267487-29268073 bp)。Hd7m與OsPRR37和C586分別相距1.9 cM和0.5 cM。推測該基因可能與Hd2等位,然而,二者是否是同一基因,需要經精細定位或等位性分析才能得到證實。
根據本試驗結果,LL次級分離群體的抽穗期出現早晚極端表現型,早抽穗植株和晚抽穗植株數目的分離比符合1∶3,推測該次級分離群體抽穗期受1對等位基因控制,且控制早抽穗的等位基因來自供體明恢63,控制晚抽穗的等位基因來自受體紅旗16。在本試驗進程中,因種植地的面積限制、環境與基因間的互作和該性狀遺傳基礎的復雜性等各方面的影響,需要對已有試驗結果的驗證和進一步試驗的補充才能得到更精確的結果,為Hd7m的精細定位和克隆提供重要信息。
參 考 文 獻:endprint
[1]Yano M, Sasaki T. Genetic and molecular dissection of quantitative traits in rice [J]. Plant Mol. Biol., 1997, 35: 145-153.
[2]李廣賢. 基于單片段代換系的水稻抽穗期QTL分析[D]. 泰安:山東農業大學, 2014.
[3]Yano M, Harushima Y, Nagamura Y, et al. Identification of quantitative trait loci controlling heading date in rice using a high density linkage map [J]. Theor. Appl. Genet., 1997, 95: 1025-1032.
[4]Lin S Y, Sasaki T, Yano M. Mapping quantitative trait loci controlling seed dormancy and heading date in rice, Oryza sativa L, using backcross inbred lines [J]. Theor. Appl. Genet., 1998, 96: 997-1003.
[5]Yano M, Kojima S, Takahashi Y, et al. Genetic control of flowering time in rice, a short-day plant [J]. Plant Physiol., 2001, 127: 1425-1429.
[6]Yano M, Katayose Y, Ashikari M, et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopis flowering time gene CONSTANS [J]. Plant Cell, 2000, 12: 2473-2483.
[7]Lin H X, Ashikari M, Yamanouchi U, et al. Identification and characterization of a quantitative trait locus Hd9, controlling heading date in rice [J]. Breed Sci., 2002,52: 35-41.
[8]Monna L, Lin H X, Kojima S, et al. Genetic dissection of a genomic region for a quantitative trait locus, Hd3, into two loci, Hd3a and Hd3b, controlling heading date in rice [J]. Theor. Appl. Genet., 2002, 104: 772-778.
[9]Lin H X, Liang Z W, Sasaki T, et al. Fine mapping and characterization of quantitative trait loci Hd4 and Hd5 controlling heading date in rice [J]. Breed Sci., 2003, 53: 51-59.
[10]Michelmore R W, Paran I, Kesseli R V. Identification of markers linked to disease-resistance genes by bulked segregation analysis: a rapid method to detect markers in specific genomic regions by using segregating populations [J]. Proc. Natl. Acad. Sci. USA, 1991, 88: 9828-9832.
[11]Panaud O, Chen X, McCouch S R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.) [J]. Mol. Gen. Genet., 1996, 252: 597-607.
[12]Zhang Q, Shen B Z, Dai X K, et al. Using bulked extremes and recessive class to map genes for photoperiod-sensitive genic male sterility in rice [J]. Proc. Natl. Acad. Sci. USA, 1994, 91: 8675-8679.
[13]Kojima S, Takahashi Y, Kobayashi Y, et al. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short day conditions [J]. Plant Cell Physiol., 2002, 43(10): 1096-1105.
[14]龔曉平, 楊正林, 趙明芳,等. 一個水稻抽穗期主基因Hd(t)的遺傳分析及分子定位[J].作物學報,2007,33 (11): 1906-1909.
[15]Kazuki M, Kiyosumi H, Eri O T, et al. Cloning of quantitative trait genes from rice reveals conservation and divergence of photoperiod flowering pathways in Arabidopsis and rice [J]. Plant Sci., 2014, 5: 193.
[16]Koo B H, Yoo S C, Park J W, et al. Natural variation in OsPRR37 regulates heading date and contributes to rice cultivation at a wide range of latitudes [J]. Mol. Plant, 2013, 6(6): 1877-1888.endprint
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
