普通野生稻花器性狀及異交結實性分析
2015-08-06 20:21:34孫佩甫郎越裴新梧袁潛華
湖北農業科學 2015年10期
孫佩甫+郎越+裴新梧+袁潛華
摘要:以7個普通野生稻(Oryza rufipogon Griff.)和3個栽培稻(Oryza sativa L.)為材料,研究普通野生稻的花器性狀和異交結實率及其相關性。結果表明,普通野生稻的花藥長度和寬度、柱頭長度和寬度及花粉量均比栽培稻大,但其花粉粒大小、花粉活力、花粉可育度和套袋自交結實率均比栽培稻小。相關分析結果表明,普通野生稻的花藥長度與花藥寬度、花粉量與花藥寬度、柱頭長度與柱頭寬度、柱頭寬度與花柱夾角分別呈顯著正相關,花粉可育度與花粉活力呈極顯著正相關;普通野生稻與栽培稻的異交結實率與其花器性狀和套袋自交結實率之間無顯著相關性。
關鍵詞:普通野生稻(Oryza rufipogon Griff.);花器性狀;自交結實率;異交結實率
中圖分類號:S511 文獻標識碼:A 文章編號:0439-8114(2015)10-2317-06
DOI:10.14088/j.cnki.issn0439-8114.2015.10.004
普通野生稻(Oryza rufipogon Griff.)是目前中國三種野生稻中分布最廣的一種野生稻,一般分布在熱帶、亞熱帶等溫度相對較高、雨量充沛的地區[1],尤其以海南省的分布密度最大,在海南省絕大部分縣(市)如海口、文昌、萬寧、東方等地區均有廣泛分布。普通野生稻長期在自然條件下適應環境的變化,具有復雜的遺傳背景和豐富的遺傳多樣性,含有高產、優質、抗病蟲、耐逆境等優異基因, 是亞洲栽培稻(Oryza sativa L.)育種的重要基因源[2]。普通野生稻與栽培稻同屬AA型基因組,親緣關系較近,兩者雜交可以獲得種子,轉基因水稻可以通過花粉擴散向普通野生稻產生基因漂流[3]。外源基因逃逸到作物的野生近緣種中可直接或間接地影響這些野生種群的遺傳和生態適應性,進而影響該野生種群的生態和進化方向[4],因而一些研究者開展了水稻向野生稻的基因漂流研究。在田間自然狀況下,Song等[5],Chen等[6]和Wang等[7]對栽培稻或轉基因水稻向普通野生稻的基因漂流進行了評估。結果表明,向普通野生稻的最大基因飄流頻率分別為2.94%,1.21%~2.19%和11.24%~18.00%,數值介于向不育系和常規稻的基因漂流頻率之間[8]。從不同研究者的研究結論來看,漂流頻率有著較大差異,這種差異主要是因為研究者采用了不同的普通野生稻居群作為受體材料,而遺傳多樣性較高的普通野生稻是高度雜合的群體[2,7],其群體中存在部分不育株和半不育株,且野生稻的柱頭外露率高、容易接受外來花粉[9],因此在基因漂流研究中作為受體的野生稻居群不同。會產生不同的基因漂流研究結果。為了客觀比較和評價轉基因水稻向不同普通野生稻的基因漂流頻率,有必要研究同等試驗條件下、不同居群普通野生稻與栽培稻在人工雜交授粉條件下的異交結實率,比較不同普通野生稻居群與栽培稻的雜交親和性,進而評價基因漂流率差異及其原因。另一方面,普通野生稻的花器性狀是否通過影響異交結實率來影響基因漂流率是值得關注的問題,已有相關學者研究了雜交水稻不育系花器特征與異交結實率的關系,但未見對普通野生稻的花器性狀與異交結實率關系的相關報道。因此需要對普通野生稻的花器性狀、自交結實率及與栽培稻的異交結實性進行相關分析,這對于水稻育種中野生稻資源利用和轉基因水稻向普通野生稻的基因漂流研究均具有重要意義。
1 材料與方法
1.1 供試材料
試驗采用7個居群的普通野生稻和3個栽培稻品種(系),普通野生稻包括海南海口、文昌、東方H、東方B(相同縣市普通野生稻編號以村名首字母為代號,以區別同一地區兩種不同的野生稻居群)、萬寧和廣西桂平、來賓居群。栽培稻包括雜交水稻恢復系9311(揚稻6號)、3550和轉bar基因水稻Bar9311(簡稱B9)。其中9311為兩系雜交稻恢復系,生育期適中,花粉量中等,株型集散適中,莖稈粗壯,穗大粒重,生育后期光合優勢強[10]。3550的生育期中等,莖稈粗壯,株型好;抗稻瘟病,恢復力強;配合力好,穗粒數多,開花散粉時間長[11,12]。B9為抗除草劑的轉基因秈稻品種,中國科學院亞熱帶生態研究所肖國櫻教授用基因槍法將bar基因導入兩系雜交早稻恢復系D68,獲得抗除草劑草銨膦的水稻恢復系 Bar68-1、從9311×Bar68-1分離后代中,篩選出抗除草劑的穩定株系,該株系生育期長、株高較高、穗長、穗粒數多、產量潛力大[13],3種栽培稻是研究異交結實的良好材料。
1.2 方法
1.2.1 試驗設計 2013年,供試材料于海南大學農學院海甸試驗基地進行播種,為了使供試材料花期相遇時間較長,分別在7月13日、7月19日、7月24日和7月28日對3種栽培稻分期播種。每期每品種10行,每行10株,行距約為16.7 cm×20.0 cm。
1.2.2 柱頭和花藥形態性狀的測定 在普通野生稻和栽培稻的開花期,隨機選取參試材料的不同稻穗中上部的5個即將開花的穎花制片[14],在ZEISS Primo Star顯微鏡下配合AxioCam ERc 5s對柱頭和花藥進行拍照,用Image-Pro Plus軟件測量柱頭長度、柱頭寬度、花柱夾角、花藥寬度和長度。
1.2.3 花粉相關性狀的測定 于7:00~8:00隨機采集稻穗插入盛有適量水的錐形瓶中,選取穎花,將單個穎花中的全部花藥放入1.5 mL 離心管中,加入1.0 mL去離子水,10 000 r/min離心 2 min,再渦旋混勻,用2.5 μL移液槍吸取1.0 μL溶液滴在載玻片上[14,15],在顯微鏡下計數其中的花粉數量,根據稀釋倍數計算單個花藥的花粉量。在顯微鏡下結合測量軟件測定花粉粒直徑,即花粉粒大小。采用I2-KI染色法測定花粉可育度[16-19],采用氯化三苯基四氮唑(TTC)染色法測量花粉活力[20]。endprint
1.2.4 自交結實率和人工雜交結實率的測定 調查稻穗套袋自交結實率和人工異交結實率,使用透明、防雨、透氣的羊皮紙袋并固定羊皮紙袋于竹竿上,防止大風折斷穗莖[21]。測定人工異交結實率需要采用溫湯去雄并結合剪穎法并進行人工輔助授粉,當普通野生稻開花時,用高20 cm的保溫壺裝滿45 ℃的溫水,溫湯5 min[22],一次浸泡2~4穗普通野生稻的稻穗,使花粉致死,并剪去穎殼的1/3便于接受花粉。采集7:00~9:00的栽培稻植株花粉以捻穗法對野生稻進行離體授粉2~3 d[23]。每個居群的野生稻設置2穗空白對照,采用溫湯去雄并剪穎,但不進行授粉。于授粉后第25~30天收集種子,計算人工異交結實率。使用SPSS軟件統計測量數據并進行相關性分析。
2 結果與分析
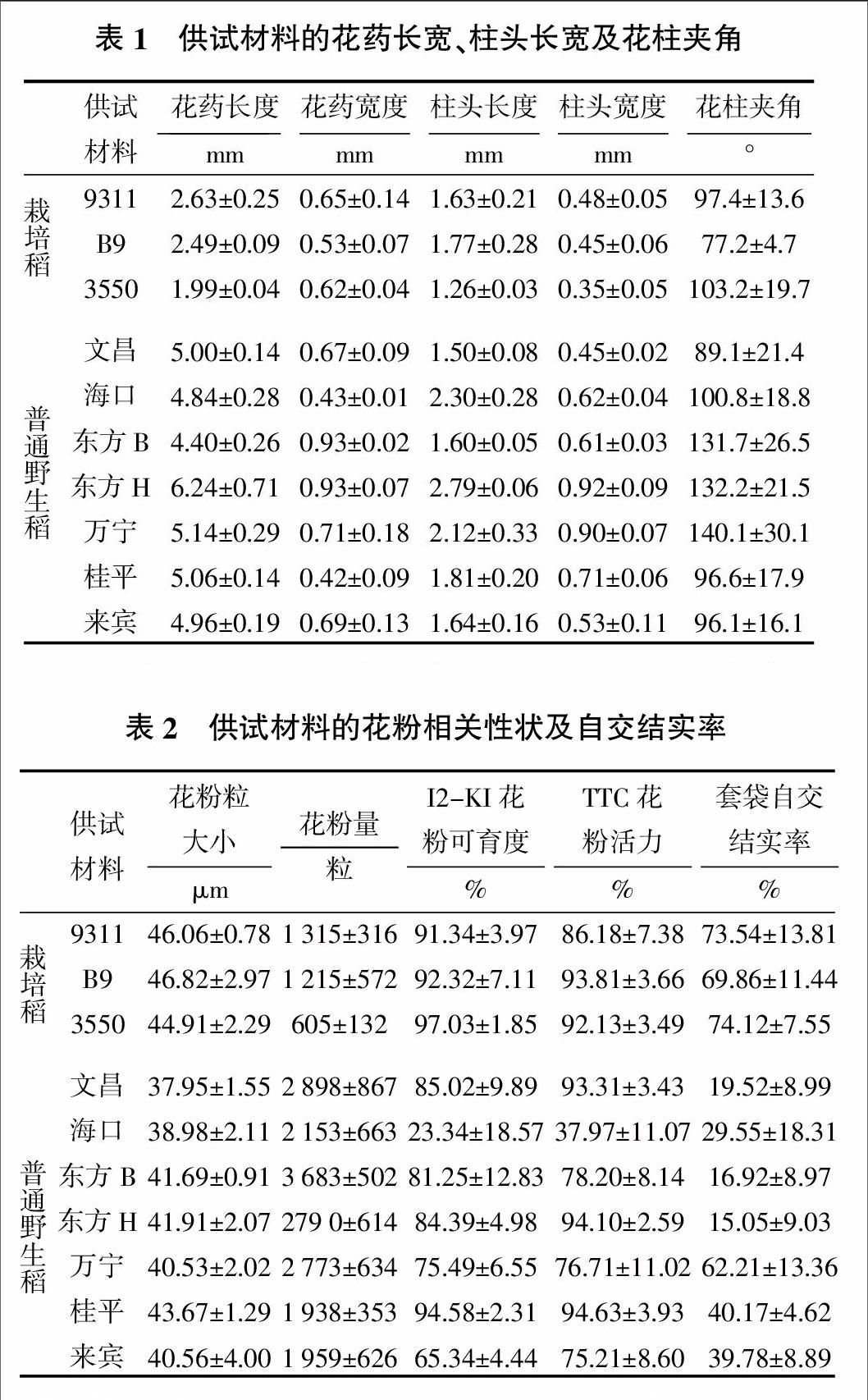
2.1 普通野生稻、栽培稻花藥和柱頭形態性狀比較
由表1可知,普通野生稻居群中花藥長度為4.40~6.24 mm,栽培稻的花藥長度為1.99 mm~2.63 mm,普通野生稻的花藥長度均值(5.09 mm)較栽培稻花藥長度均值(2.37 mm)長2.72 mm,所有供試的普通野生稻花藥長度均較栽培稻長。普通野生稻花藥寬度為0.42~0.93 mm,栽培稻的花藥寬度為0.53~0.65 mm,栽培稻的花藥寬度均值(0.60 mm)較普通野生稻的花藥寬度均值(0.68 mm)小0.08 mm。供試材料中普通野生稻的柱頭長度為1.50 ~2.79 mm,栽培稻的柱頭度長為1.26~1.77 mm,普通野生稻的柱頭長度均值(1.97 mm)較栽培稻的柱頭長的均值(1.55 mm)長0.42 mm。普通野生稻柱頭寬度為0.45~0.92 mm,栽培稻中柱頭寬度0.35~0.48 mm,普通野生稻的柱頭寬度均值為0.68 mm,除了文昌的柱頭寬與栽培稻的柱頭寬相近,其余的普通野生稻柱頭寬均較栽培稻的長。普通野生稻的花柱夾角均值為112.4°,較栽培稻的花柱夾角均值(92.6°)大19.8°。可見普通野生稻的花藥長和花藥寬、柱頭長和柱頭寬均比栽培稻長,花柱夾角的均值也略大于栽培稻。
2.2 普通野生稻和栽培稻的花粉相關性狀及套袋自交結實率的比較
由表2可知,供試材料中普通野生稻的花粉粒大小為37.95~43.67 μm,栽培稻的花粉粒大小為44.91~46.82 μm,栽培稻的花粉粒大小均值(45.93 μm)較普通野生稻的均值(40.76 μm)大5.17 μm(表2)。有相關研究表明栽培稻的花粉粒大小為36~56 μm,多數在42~50 μm的范圍內[24],本研究的栽培稻的花粉粒直徑在多數水稻花粉粒大小的范圍內,但普通野生稻的花粉粒直徑偏小。
普通野生稻居群中花粉數量為1 938~3 683粒,栽培稻的花粉量為605~1 315粒。楊弘遠[24]調查了4個栽培稻品種的一個花藥內約有1 000粒左右的花粉量,與本研究的栽培稻的花粉量基本一致,其中栽培稻3550的花粉量偏小,而普通野生稻的花粉量遠大于栽培稻的花粉量。栽培稻品種中花粉可育度在91.34%以上,花粉活力在86.18%以上,而普通野生稻的花粉可育度為23.34%~94.58%,花粉活力為37.97%~94.63%,兩項指標的均值均較栽培稻小。
普通野生稻居群的自交結實率為15.05%~62.21%,均值為31.89%,變化范圍比較大;栽培稻的自交結實率在69.86%~74.12%,均值為72.51%,且變化范圍較小,表明栽培稻品種套袋自交結實率穩定。栽培稻的自交結實率明顯較普通野生稻高,可能與栽培稻經過長期的人工農藝性狀選擇有關。另一方面,栽培稻的花粉粒大小較普通野生稻大,且花粉可育度和花粉活力均較普通野生稻大,花粉粒貯存的淀粉等營養物質多,因而其花粉粒萌發能力及花粉管伸長均具有優勢,從而更容易受精結實。
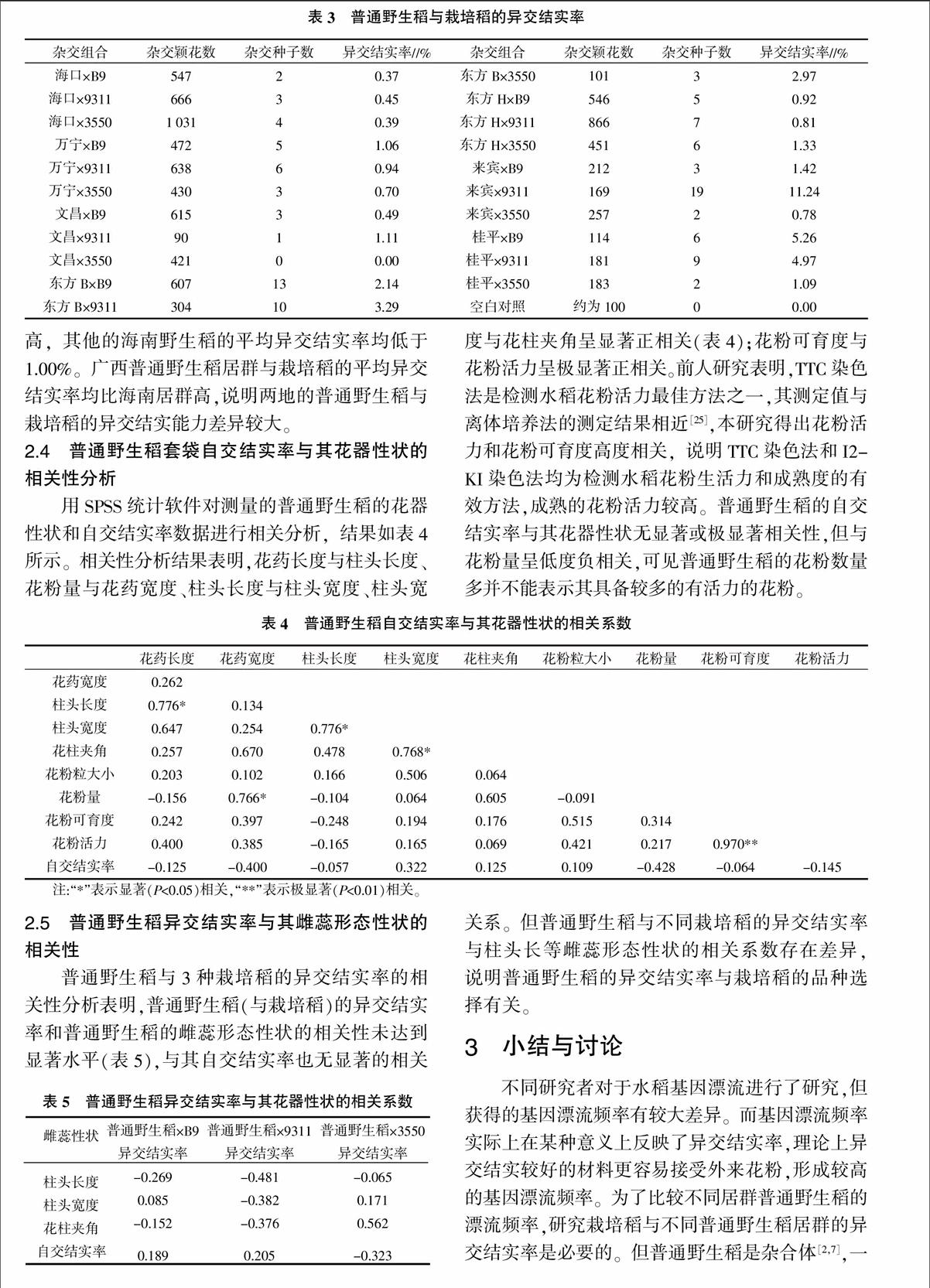
2.3 普通野生稻與栽培稻的異交結實性分析
以普通野生稻為母本、栽培稻為父本進行人工雜交。本試驗中對普通野生稻先進行人工去雄,再進行人工授粉,配置雜交組合,各組合的異交結實率見表3。由表3可知,空白對照所獲得的雜交種子均為0,說明人工雜交過程中去雄效果好,無自交結實現象的干擾。雜交組合中來賓×9311異交結實率最高,為11.24%,說明在這些組合中來賓與9311親和性最好,桂平×B9(5.26%)和桂平×9311(4.97%)也較高,而海口與三種栽培稻的異交結實率很低,文昌×3550的異交結實率為0,雜交親和性最差。普通野生稻與栽培稻的異交結實率不同,說明栽培稻與不同普通野生稻的親和性不同。同時不同栽培稻與同一種普通野生稻的異交結實率也有差異。
根據表3數據可以求得所有普通野生稻與栽培稻雜交的平均異交結實率。B9、9311和3550的平均異交結實率分別為1.67%、3.26%和1.04%,以9311平均異交結實率最大,最小為3550。普通野生稻與所有的栽培稻的平均異交結實率大小依次為來賓(4.48%)>桂平(3.77%)>東方B(2.80%)>東方H(1.02%)>萬寧(0.90%)>文昌(0.53%)>海口(0.40%)。廣西來賓和桂平的平均異交結實率相對最高,海口最低。海南野生稻中以東方B、東方H較高,其他的海南野生稻的平均異交結實率均低于1.00%。廣西普通野生稻居群與栽培稻的平均異交結實率均比海南居群高,說明兩地的普通野生稻與栽培稻的異交結實能力差異較大。
2.4 普通野生稻套袋自交結實率與其花器性狀的相關性分析
用SPSS統計軟件對測量的普通野生稻的花器性狀和自交結實率數據進行相關分析,結果如表4所示。相關性分析結果表明,花藥長度與柱頭長度、花粉量與花藥寬度、柱頭長度與柱頭寬度、柱頭寬度與花柱夾角呈顯著正相關(表4);花粉可育度與花粉活力呈極顯著正相關。前人研究表明,TTC染色法是檢測水稻花粉活力最佳方法之一,其測定值與離體培養法的測定結果相近[25],本研究得出花粉活力和花粉可育度高度相關,說明TTC染色法和I2-KI染色法均為檢測水稻花粉生活力和成熟度的有效方法,成熟的花粉活力較高。普通野生稻的自交結實率與其花器性狀無顯著或極顯著相關性,但與花粉量呈低度負相關,可見普通野生稻的花粉數量多并不能表示其具備較多的有活力的花粉。endprint
2.5 普通野生稻異交結實率與其雌蕊形態性狀的相關性
普通野生稻與3種栽培稻的異交結實率的相關性分析表明,普通野生稻(與栽培稻)的異交結實率和普通野生稻的雌蕊形態性狀的相關性未達到顯著水平(表5),與其自交結實率也無顯著的相關關系。但普通野生稻與不同栽培稻的異交結實率與柱頭長等雌蕊形態性狀的相關系數存在差異,說明普通野生稻的異交結實率與栽培稻的品種選擇有關。
3 小結與討論
不同研究者對于水稻基因漂流進行了研究,但獲得的基因漂流頻率有較大差異。而基因漂流頻率實際上在某種意義上反映了異交結實率,理論上異交結實較好的材料更容易接受外來花粉,形成較高的基因漂流頻率。為了比較不同居群普通野生稻的漂流頻率,研究栽培稻與不同普通野生稻居群的異交結實率是必要的。但普通野生稻是雜合體[2,7],一些群體中存在部分不育和半不育株,為了防止自身花粉影響到異交結實,需要人工去除自身可育花粉,對普通野生稻先采用溫湯等方法去雄,再行人工授粉雜交,以獲得真實的異交結實率。
在田間自然狀況下,栽培稻或轉基因水稻向普通野生稻的最大基因飄流頻率為1.21%~18.00%, 該數值介于向不育系和常規稻的基因漂流頻率之間[5-8,26]。本試驗通過栽培稻與不同普通野生稻的人工雜交,研究普通野生稻的異交結實率,其范圍為0%~11.24%,平均值為1.99%。但需要指出的是,盡管基因飄流頻率與試驗的異交結實率屬性相似,但計算方法有較大差異。在轉基因水稻基因漂流研究過程中,其試驗設計一般是在試驗中心設一個轉基因稻樣方, 周圍種植非轉基因稻或其親緣種,于水稻成熟期在不同方向的不同距離上取一定面積的樣方或一定數量的植株或種子, 分析植株或成熟種子轉基因存在的頻率[27],即作為轉基因傳粉的頻率,以評價外源轉基因通過授粉雜交途徑的擴散過程。這種方法會因外來花粉(或自身花粉,如野生稻)競爭大小造成結實率高低不同,從而影響基因漂流頻率的計算值。但如果參試的受體為雜交水稻的不育系時,由于不育系自身無可育花粉或者無花粉,其結實率高低受外來花粉多少的影響較大,而當周圍無常規水稻競爭花粉、只有轉基因供體花粉的情況下,授粉全部來自于轉基因花粉源,結實的子粒全部或絕大部分為轉基因種子,只有極少數來自于不純的不育系雜株,這樣估算的基因漂流頻率往往偏高。對于雜合群體的普通野生稻,群體中不育株或半不育株的數量將影響到競爭花粉的數量,進而影響基因漂流頻率。而異交結實率反映的是受體異交結實的能力,在自然條件下其接受的花粉既可來自普通野生稻自身,也可來自外來花粉。本試驗中人工異交結實率遠低于其套袋自交結實率,除了與雜交親和性有關外,可能還與人工去雄對普通野生稻雌蕊器官造成傷害進而影響授粉有關。
普通野生稻自交結實和異交結實的研究中,大田試驗中下雨天氣往往使套袋潮濕甚至霉變,影響受(授)精結實,造成空粒或雜交稻穗的損失,而且普通野生稻植株較高且莖稈細小易折使人工輔助授粉困難,使得人工去雄雜交過程非常艱難,樣本數量難以達到栽培稻之間進行人工雜交獲得種子的數量級。
影響異交結實的花器性狀很多, 柱頭外露率是其中非常重要的因素。在雜交水稻制種中一般圍繞提高母本柱頭外露率、 父母本開花時相遇率等調節措施來提高制種產量, 并改進授粉技術。龍俐華等[28]以三系不育系為材料, 研究結果表明, 雜交水稻制種中柱頭外露率與異交結實率呈極顯著相關(r=0 .966 7)。但不同居群的普通野生稻的柱頭外露率均較高,差異較小,大多高于95%[29],因此本研究不考慮柱頭外露率與其他花器性狀及自交、異交結實率的關系。
水稻的柱頭長寬、花藥長寬與柱頭夾角等花器形態性狀與異交結實率的相關研究表明,粳稻三系不育系的異交結實率與其花柱長度呈極顯著正相關,而花藥長與柱頭長、柱頭長與柱頭寬無顯著相關性[30];恢復系柱頭長寬與花柱夾角無顯著相關,而花粉量與花藥長度呈極顯著線性正相關(r=0.911 2),與花藥寬度的正相關系數雖然也很高,但未達到顯著水平(r=0.679 7)[14]。本試驗對普通野生稻雌蕊花器性狀和自交結實率相關性分析結果表明,普通野生稻的花藥長與柱頭長、柱頭長與柱頭寬、柱頭寬與花柱夾角、花粉量與花藥寬分別呈顯著正相關,花粉可育度和花粉活力呈極顯著正相關。異交結實率與自交結實率和雌蕊花器形態性狀的相關分析表明,普通野生稻的雌蕊形態性狀對其(與栽培稻的)異交結實率和自交結實率無顯著影響,這與張忠旭[14]和全東興等[30]在雜交水稻上的研究結果有差異。
柱頭活力關系到花粉飄落到柱頭之后能否萌發、花粉管能否生長,所以柱頭活力直接關系到受精的成敗。普通野生稻的柱頭活力也應影響其異交結實進而影響基因漂流率。在植物界中,柱頭活力持續時間的長短因植物種類而異,水稻一般能維持6~7 d,以開花當天相關活力最強,以后逐漸下降。在雜交水稻制種實踐中,不育系柱頭活力強有利于異交結實和制種產量的提高[31,32]。研究柱頭活力一般針對不育系材料, 對不育系的穎花進行處理, 通過人工授粉的結實率高低來衡量開花后不同天數的外露柱頭活力。但對于野生稻, 由于自身含有可育花粉,通過異交結實率判定柱頭活力或會導致誤差, 若用溫湯去雄等方法也會在一定程度上傷害柱頭等雌蕊器官, 影響柱頭活力研究的準確性。因此對野生稻柱頭活力的評價方法尚有待于進一步研究。
參考文獻:
[1] 范樹國,張再君,劉 林,等.中國野生稻的種類、地理分布及其特征特性綜述[J].武漢植物學研究,2000,18(5):417-425.
[2] 楊慶文,黃 娟.中國普通野生稻遺傳多樣性研究進展[J].作物學報,2013,39(4):580-588.
[3] JIA S R, YUAN Q H, PEI X W, et al. Rice transgene flow: its patterns,model and risk management[J]. Plant Biotechnology Journal, 2014, 12(9): 1262-1273.endprint
[4] 盧寶榮.全球轉基因逃逸及其生態后果的最新研究動態[J].生物多樣性,2003,11(2):177-178.
[5] SONG Z P,LU B R,CHEN J K, et al.Gene flow from cultivated rice to the wild species Oryza rufipogon under experimental field conditions[J].New Phytologist, 2003,157 (24):657-665.
[6] CHEN L J, LI D S,SONG Z P, et al .Gene flow from cultivated rice (Oryza sativa) to its weedy and wild relatives [J]. Annals of Botany, 2004, 93(1):67-73.
[7] WANG F, YUAN Q H, SHI L, et al. A large scale field study of transgene flow from cultivated rice (Oryza sativa) to common wild rice(O. rufipogon) and barnyard grass (Echinochloa crusgalli)[J]. Plant Biotechnology Journal,2006,4(6):667-676.
[8] 袁潛華,裴新梧,何美丹.轉基因水稻向普通野生稻的基因漂流研究[R].國家科技報告服務系統.2013.(館藏號:306--2013-035209).(http://www.nstrs.cn/xiangxiBG.aspx?id=35209).
[9] 賈士榮,袁潛華,王 豐,等.轉基因水稻基因飄流研究十年回顧[J].中國農業科學,2014,47(1):1-10.
[10] 戴正元, 趙步洪, 劉曉靜,等. 優質高產多抗中秈新品種揚稻6號[J].江蘇農業科學,1997(4):13-14.
[11] 彭惠普,李維明,伍應運,等.廣譜恢復系3550及其系列雜交稻的選育和應用Ⅰ.恢復系3550的選育及其特征特性[J].雜交水稻,1993(6):1-3.
[12] 彭惠普,李維明,伍應運,等.廣譜恢復系3550及其系列雜交稻的選育和應用Ⅱ.3550系列雜交稻組合的試種及推廣應用[J]. 雜交水稻,1994(1):9-11.
[13] 何美丹,徐立新,袁潛華,等.秈、粳型轉基因水稻花粉源基因飄流的比較研究[J].南方農業學報,2012,43(5):559-565.
[14] 張忠旭.北方雜交粳稻異交結實機制及高產制種技術研究[D].沈陽:沈陽農業大學,2009.
[15] 郭軍洋,陳龍正,曹清河,等.黃瓜現采花粉生活力最佳染色方法的篩選[J].廣東農業科學,2004,6(1):48-49.
[16] ZHANG Y, LIYF, ZHANG J, et al. Characterization and mapping of a new male sterility mutant of anther advanced dehiscence (t) in rice[J]. Journal of Genetics and Enomics,2008,35(3):177-182.
[17] 李自超,王象坤,庫爾班.I-KI溶液鑒別水稻花粉育性可行性(簡報)[J].北京農業大學學報,1993(4):109-110.
[18] 袁隆平.雜交水稻簡明教程[M].長沙:湖南科學技術出版社,1985.
[19] 盧興桂,袁潛華,姚克敏,等.光溫敏雄性不育水稻的生態適應性研究[M].北京:氣象出版社,2001.
[20] KOGA Y,AKIHAMA T, FUJIMAKI H. Study on the longevity of pollen grains of rice, Oryza sativa L.:Ⅰ.Morphological change of pollen grains after shedding[J]. Cytologia,1971,36(1):104-110.
[21] 姚克敏,袁潛華.水稻不育系育性觀察指標套袋自交結實率和花粉可育度的比較分析[J].雜交水稻,1998(4):25-27.
[22] 徐國生,譚文兵.水稻雜交剪穎溫湯去雄方法的研究[J].湖南農業科學,1983(3):17-18.
[23] 郭嗣斌,林興華.小粒野生稻與栽培稻遠緣雜交探討[J].廣西農業科學,2010,41(2):95-99.
[24] 楊弘遠.水稻生殖生物學[M].杭州:浙江大學出版社,2005.
[25] 劉壽東,史佩劍,江曉東,等.轉基因水稻B2花粉活力的溫度模型[J].中國水稻科學,2011,25(2):219-222.
[26] 尹昭坤,喬 健,袁潛華.轉基因水稻向普通野生稻及其他近緣植物的基因漂流研究進展[J].熱帶生物學報,2011,2(4):373-377.
[27] 程 林,韓 飛,袁潛華.轉基因稻向栽培稻及其稻屬植物的基因漂流研究進展[J].貴州科學,2007,25(4):42-46.
[28] 龍俐華,舒 琨.提高秈型雜交水稻制種異交結實率的研究[J].湖南農業大學學報,2000,26(3):167-170.
[29] 余昌花.轉bar基因水稻向普通野生稻的基因漂流及與開花習性的關系[D].海口:海南大學,2011.
[30] 全東興,周廣春,孟維韌.粳稻三系不育系花器特征與異交結實率的關系分析[J].雜交水稻,2010(S1):487-491.
[31] 米佳佳, 劉常金,王汝華,等. 七個水稻粳型不育系的柱頭活力[J].湖北農業科學,2014,53(9):2003-2005.
[32] 賀曉鵬,楊書化,田亞雯,等.5個新育成粳型光溫敏核不育系開花及異交性能的研究[J].中國農學通報,2004,20(4):104-107.endprint
