哈氏弧菌谷胱甘肽還原酶基因克隆及原核表達(dá)
2015-10-27 01:25:23朱帆丁燏魯義善簡紀(jì)常吳灶和
生物技術(shù)通報 2015年6期
朱帆丁燏魯義善簡紀(jì)常吳灶和
(1.廣東海洋大學(xué)水產(chǎn)學(xué)院,湛江 524088;2.廣東省水產(chǎn)經(jīng)濟(jì)動物病原生物學(xué)及流行病學(xué)重點實驗室,湛江 524088;3.廣東省教育廳水產(chǎn)經(jīng)濟(jì)動物病害控制重點實驗室,湛江 524088;4.仲愷農(nóng)業(yè)工程學(xué)院,廣州 510225)
哈氏弧菌谷胱甘肽還原酶基因克隆及原核表達(dá)
朱帆1,2,3丁燏1,2,3魯義善1,2,3簡紀(jì)常1,2,3吳灶和2,3,4
(1.廣東海洋大學(xué)水產(chǎn)學(xué)院,湛江 524088;2.廣東省水產(chǎn)經(jīng)濟(jì)動物病原生物學(xué)及流行病學(xué)重點實驗室,湛江 524088;3.廣東省教育廳水產(chǎn)經(jīng)濟(jì)動物病害控制重點實驗室,湛江 524088;4.仲愷農(nóng)業(yè)工程學(xué)院,廣州 510225)
經(jīng)克隆哈氏弧菌谷胱甘肽還原酶(GR)基因,并構(gòu)建其原核表達(dá)載體,以獲得相應(yīng)的表達(dá)蛋白。將GR和pET-32a(+)通過BamH I和Xho I雙酶切后,體外用T4連接酶連接,構(gòu)建重組質(zhì)粒pET-GR;然后轉(zhuǎn)化至大腸桿菌BL21(DE3)中,利用異丙基-β-D-硫代半乳糖苷(IPTG)誘導(dǎo)表達(dá),應(yīng)用SDS-PAGE分析表達(dá)情況和表達(dá)條件。SDS-PAGE電泳獲得分子量約為68.9 kD融合蛋白條帶。在E.coli BL21(DE3)中重組質(zhì)粒pET-GR的表達(dá)條件為28℃,0.7 mmol/L的IPTG濃度誘導(dǎo)4 h表達(dá)量最高,且主要以包涵體形式表達(dá)。哈氏弧菌谷胱甘肽還原酶基因在大腸桿菌中獲得了高效表達(dá)。
哈氏弧菌;谷胱甘肽還原酶;基因克隆;原核表達(dá)
哈氏弧菌(Vibrio harveyi)是一種革蘭氏陰性、發(fā)光的海洋浮游細(xì)菌,廣泛分布于近岸溫暖的海水、海洋沉積物、海洋動物的體表等各種海洋場所[1],是近10多年才被認(rèn)識的水產(chǎn)養(yǎng)殖動物重要的條件致病菌。暴發(fā)性弧菌病是對蝦的一種嚴(yán)重疾病,曾引起多個國家對蝦養(yǎng)殖的大面積死亡,如墨西哥[2]、菲律賓[3]、澳大利亞[4]、中國[5]等,給對蝦養(yǎng)殖造成災(zāi)難性的經(jīng)濟(jì)損失[6]。該菌同時能夠感染大西洋棘白鯧(Chaetodipterus faber)[7]、大黃魚(Pseudosciaena crocea)[8]、斜帶石斑魚(Epinephelus coioides)[9]等多種養(yǎng)殖魚類。隨著海水養(yǎng)殖業(yè)的迅速發(fā)展,弧菌病的流行也越來越多。
目前,抗生素仍然是控制弧菌病的主要手段,然而由于抗生素的大量使用導(dǎo)致耐藥菌的出現(xiàn),使弧菌病的控制變得更加困難[10,11]。細(xì)菌耐藥的形成機(jī)制有目標(biāo)靶蛋白的修飾、主動藥物轉(zhuǎn)運(yùn)系統(tǒng)、細(xì)胞外膜通透性的改變等[12,13],有研究表明,細(xì)菌在抗生素的刺激下會產(chǎn)生氧化應(yīng)激,細(xì)胞內(nèi)產(chǎn)生過量的氧化物和自由基(Reactive oxygen species,ROS),導(dǎo)致細(xì)胞內(nèi)的氧化和抗氧化失去平衡,從而引起細(xì)胞損傷[14,15]。因此細(xì)菌細(xì)胞對活性氧和自由基的清除可能是耐藥性形成的一個原因。谷胱甘肽(Glutathione,GSH)是廣泛存在于動植物、微生物細(xì)胞內(nèi)重要的抗氧化劑,能夠有效清除活性氧自由基,維持細(xì)胞內(nèi)穩(wěn)定的氧化還原狀態(tài)[16,17]。GSH參與的大部分反應(yīng)都是還原態(tài)形式,還原型GSH失去電子形成氧化型谷胱甘肽(Glutathione oxidized,GSSG),谷胱甘肽還原酶(Glutathione reductase,GR)以還原型輔酶Ⅱ(NADPH)作為電子供體還原GSSG形成GSH[18]。而GSH的抗氧化能力主要取決于細(xì)胞內(nèi)GSH的濃度和GSH/GSSG的比率[19],因此GR是GSH抗氧化作用中的重要組成部分,與細(xì)菌的耐藥性形成也密切相關(guān)。
本實驗構(gòu)建含哈氏弧菌GR基因的大腸桿菌BL21(DE3)重組菌株,并對融合蛋白表達(dá)的IPTG誘導(dǎo)條件進(jìn)行優(yōu)化,旨為進(jìn)一步研究GR的功能特別是在細(xì)菌耐藥性中的作用奠定基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 菌株和質(zhì)粒 哈氏弧菌(Vibrio harveyi)ZJ0603,E.coli DH5α和E.coli BL21(DE3)均由本實驗室保存;克隆質(zhì)粒pMD18-T,內(nèi)切酶BamH I,Xho I均購自TaKaRa公司;原核表達(dá)質(zhì)粒pET-32a(+)由本實驗室保存。
1.1.2 主要試劑 細(xì)菌基因組DNA提取試劑盒,DNA凝膠回收純化試劑盒,購自Thermo公司;PCR試劑盒,T4連接酶,購自TaKaRa公司;PCR引物由上海生工合成;BamH I、Xho I均購自大連寶生物公司。
1.2 方法
1.2.1 GR基因克隆 使用細(xì)菌DNA提取試劑盒,提取哈氏弧菌基因組,根據(jù)GenBank中已發(fā)表的哈氏弧菌VIBHAR_00523的GR基因序列(登錄號5552818),利用Primer 5.0設(shè)計引物,grs 5'-CGGG ATCCATGGCGACTCATTTTGATTAT-3'(酶切位點BamH I)gra 5'-CCGCTCGAGCTAACCTGTCATGGT AACGAAC-3'(酶切位點Xho I)。以基因組DNA為模板擴(kuò)增GR的ORF,PCR程序為:94℃預(yù)變性4 min;94℃ 40 s,61℃ 40 s,72℃ 1 min,28個循環(huán);72℃,7min;4℃ forever。PCR結(jié)束后取產(chǎn)物進(jìn)行1.0%瓊脂糖凝膠電泳檢測,并回收PCR產(chǎn)物,-20℃保存。
1.2.2 PCR產(chǎn)物檢測 將回收的PCR產(chǎn)物16℃過夜連接到pMD18-T載體,命名為pMD18-gr,取10 μL連接產(chǎn)物轉(zhuǎn)化至E.coli DH5α感受態(tài)細(xì)胞[20];轉(zhuǎn)化產(chǎn)物涂布于含氨芐青霉素的LB瓊脂平板,37℃培養(yǎng)8-10 h。挑取單個轉(zhuǎn)化菌落,接種于1 mL含氨芐青霉素的LB液體培養(yǎng)基中,37℃ 200 r/min培養(yǎng)1-2 h,菌落PCR檢測陽性的單克隆送至上海生工測序部測序。
1.2.3 pET-gr目標(biāo)載體的構(gòu)建 測序結(jié)果比對正確后,提取重組質(zhì)粒pMD18-gr和質(zhì)粒pET-32a(+),經(jīng)過雙酶切后用T4 DNA連接酶將酶切后的目的片段和pET-32a(+)連接構(gòu)建重組質(zhì)粒,命名為pET-gr,再轉(zhuǎn)入受體菌E.coli BL21(DE3),PCR檢測陽性克隆并測序。
1.2.4 重組目標(biāo)蛋白的誘導(dǎo)表達(dá) 將含有重組質(zhì)粒pET-gr的大腸桿菌BL21按1∶50接種于含氨芐抗性的LB液體培養(yǎng)基中37℃ 200 r/min培養(yǎng)。當(dāng)OD600達(dá)到0.4-0.6時,加IPTG至終濃度為1 mmol/L,繼續(xù)培養(yǎng)4 h。取1 mL菌液于1.5 mL EP管中,10 000 r/min離心2 min,棄上清,菌體經(jīng)預(yù)處理后進(jìn)行SDS-PAGE分析。以同樣的方法誘導(dǎo)含空質(zhì)粒pET-32a(+)的Ecoli BL21(DE3)作對照;并從時間、溫度、IPTG濃度3個方面探索目標(biāo)蛋白表達(dá)的最佳條件。IPTG濃度:其他培養(yǎng)條件不變時,設(shè)置不同的IPTG濃度0.1、0.2、0.4、0.7和1.0 mmol/L誘導(dǎo),分別處理菌體進(jìn)行SDS-PAGE分析;時間:其他培養(yǎng)條件不變時,在誘導(dǎo)后的2、4、6、8和10 h分別取樣,分別處理菌體,進(jìn)行SDS-PAGE分析;溫度:其他培養(yǎng)條件不變時,在不同溫度28℃,37℃條件下誘導(dǎo)培養(yǎng),分別收集菌體后利用超聲波破碎,超聲程序為:超聲破碎6 s,間歇6 s,保護(hù)溫度設(shè)置為37℃,直至菌液澄清,然后離心收集上清和沉淀進(jìn)行SDS-PAGE分析。
2 結(jié)果
2.1 目的基因ORF克隆與分析
設(shè)計的GR引物,經(jīng)PCR擴(kuò)增分別得到1 000 bp左右的條帶(圖1-A),連接至pMD18-T vector后轉(zhuǎn)化入E.coli DH5α,菌落PCR擴(kuò)增的條帶大小與之相符(圖1-B)。
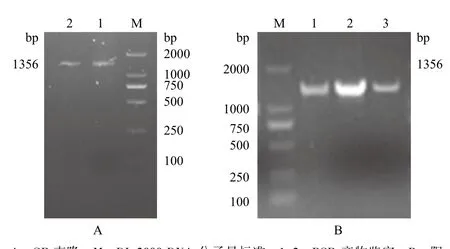
圖1 GR基因的克隆(A)及菌落PCR(B)
根據(jù)NCBI網(wǎng)站的ORF Finder軟件分析,GR基因序列包含一個1 356 bp的完整開放閱讀框(ORF),編碼451個氨基酸,分子量預(yù)測為48.9 kD,理論等電點為5.29,SignalP 3.0 Server分析顯示該基因沒有信號肽序列,TMHMM Server v. 2.0分析該基因不存在跨膜區(qū)。NetGlyc 1.0 Server 預(yù)測其沒有N-糖基化位點,NetPhos 2.0 Server預(yù)測該基因有3個絲氨酸磷酸化位點。8個酪氨酸磷酸化位點和4個蘇氨酸磷酸化位點,一個cAMP和cGMP依賴性的蛋白激酶磷酸化位點,2個蛋白激酶c磷酸化位點,6個酪蛋白激酶II磷酸化位點,14個N-十四酰化位點,一個CAAX box,11個微體C末端定位信號序列。氨基酸同源性分析序列分析顯示,哈氏弧菌GR基因與輪蟲弧菌(V.Rotiferianus),副溶血弧菌(V.parahaemolyticus)和溶藻弧菌(V.alginolyticus)都具有很高的同源性,其中與輪蟲弧菌的同源性達(dá)到99%。
2.2 原核表達(dá)
2.2.1 原核表達(dá)載體的構(gòu)建 克隆的GR基因連接到pET-32a(+)轉(zhuǎn)化至E.coli BL21(DE3)感受態(tài)細(xì)胞中,陽性菌落的PCR鑒定有約1 356 bp的亮帶(圖3-A),提取質(zhì)粒經(jīng)Xho I和BamH I雙酶切鑒定(圖3-B),出現(xiàn)的條帶大小與預(yù)期結(jié)果一致,測序并分析證明原核表達(dá)載體pET32-gr構(gòu)建成功。
2.2.2 重組質(zhì)粒的誘導(dǎo)表達(dá) 含重組質(zhì)粒pET-gr的大腸桿菌BL21經(jīng)IPTG誘導(dǎo),得到約68.9 kD的融合蛋白,其中預(yù)計GR分子量為48.9 kD,pET-32a(+)表達(dá)的融合標(biāo)簽約為20 kD;沒有誘導(dǎo)的含重組質(zhì)粒的E.coli BL21(DE3)和誘導(dǎo)的含pET-32a(+)空載體的E.coli BL21(DE3)作為對照,未發(fā)現(xiàn)目的蛋白表達(dá)(圖4),因此目的基因表達(dá)成功。
GR基因表達(dá)的條件優(yōu)化結(jié)果表明,GR在IPTG濃度為0-1.0 mmol/L范圍內(nèi),表達(dá)量先升高后降低,在0.7 mmol/L時表達(dá)量最大(圖5-A),誘導(dǎo)4 h時表達(dá)量最大(圖5-B),最適誘導(dǎo)溫度為28℃(圖5-C)。在最佳誘導(dǎo)條件下誘導(dǎo)表達(dá)后,收集重組菌經(jīng)超聲破碎細(xì)胞,SDS-PAGE分析表明,37℃和28℃誘導(dǎo)時沉淀和上清中都有明顯的目的蛋白條帶,但是28℃誘導(dǎo)時沉淀和上清的目的蛋白條帶均相對于37℃誘導(dǎo)時的亮,并且兩種溫度誘導(dǎo)下沉淀中的目的蛋白條比上清中的目的蛋白多(圖5-C),表明重組蛋白在E.coli BL21(DE3)中主要以包涵體形式存在。
3 討論
GSH作為細(xì)胞內(nèi)重要的抗氧化劑可以和許多化合物反應(yīng),參與細(xì)胞內(nèi)許多生理過程,以維持細(xì)胞內(nèi)硫醇的氧化還原狀態(tài)。GSH的抗氧化能力是通過為關(guān)鍵的抗氧化防御酶如抗壞血酸、過氧化物酶等提供還原當(dāng)量來實現(xiàn)的[21]。但在清除ROS的解毒過程中,形成大量的谷胱甘肽二硫化物(GSSG)即GSH的氧化形式,而GR則以NADPH作為輔酶,催化GSSG轉(zhuǎn)變成GSH[22],它屬于一種黃素蛋白還原酶。因此,GR通過保持較高的GSH/GSSG比率在維持細(xì)胞內(nèi)穩(wěn)定的氧化還原水平中具有重要作用。
目前,在許多生物中已克隆出編碼GR的基因組DNA或cDNAs,包括細(xì)菌[23]、植物[24]、和小鼠[25]等。并且通過酶活性測定和基因表達(dá)定量分析,對GR在植物的抗逆性和抗氧化,以及動物癌細(xì)胞的耐藥性方面的作用已經(jīng)進(jìn)行了較多的研究。研究顯示,GR在植物抗脅迫時參與氧化應(yīng)激和對刺激的耐受性具有重要作用[26],具有較低GR活性的植物對溫度、重金屬、鹽度等環(huán)境脅迫更加敏感,GR基因缺失菌株在環(huán)境脅迫因子刺激下存活率更低,重組高產(chǎn)GSH合成酶系統(tǒng)使大腸桿菌對重金屬的耐受性提高[27],因此GSH抗氧化系統(tǒng)在植物抗逆性、抗脅迫、動物腫瘤耐藥性治療、細(xì)胞抗氧化、抗衰老等方面具有廣闊的應(yīng)用前景,但是細(xì)菌耐藥性的形成與GSH抗氧化系統(tǒng)的相關(guān)性研究相對較少。

圖2 GR基因核苷酸序列及推導(dǎo)的氨基酸序列
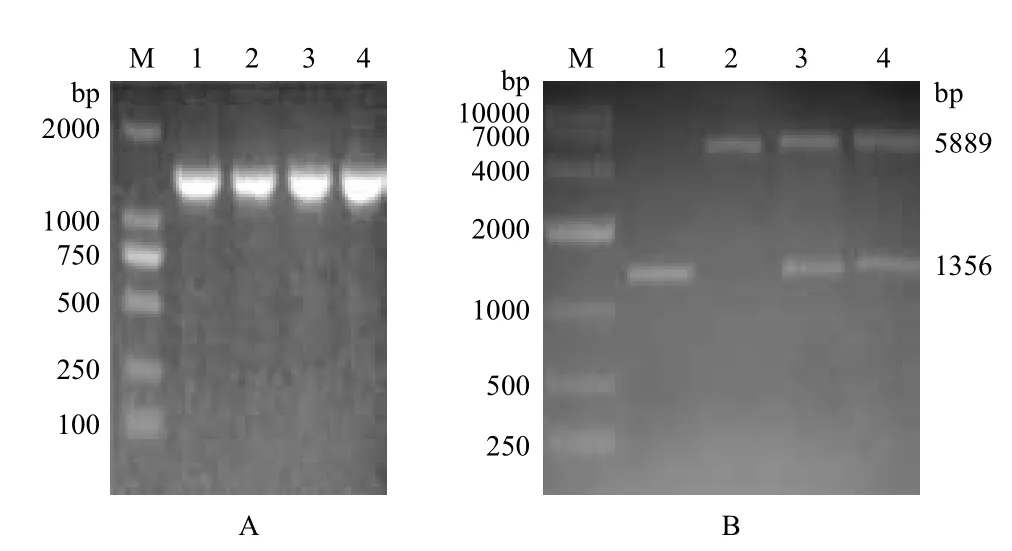
圖3 GR 原核表達(dá)載體構(gòu)建
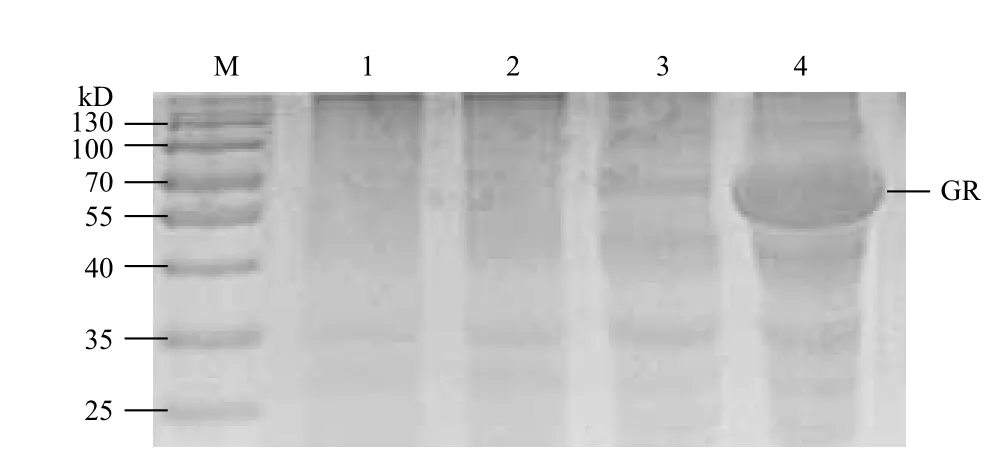
圖4 pET-gr原核表達(dá)分析
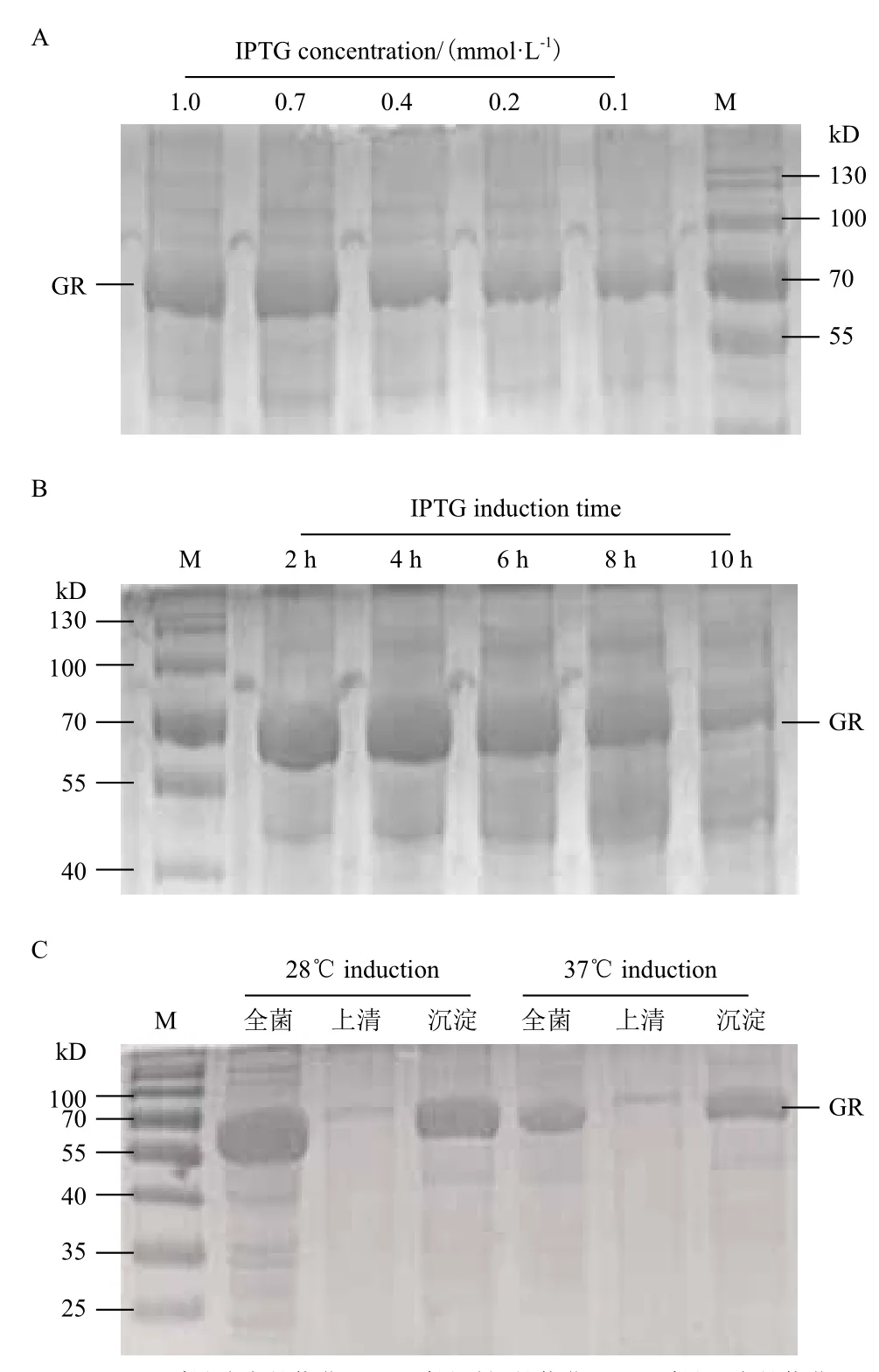
圖5 pET-gr誘導(dǎo)條件的優(yōu)化
抗生素作用于細(xì)菌,引起細(xì)菌細(xì)胞內(nèi)的氧化應(yīng)激時,過量氧化物和自由基消耗GSH,導(dǎo)致GSH/ GSSG比率降低,GSH合成酶體系的表達(dá)和GSSG的還原途徑在調(diào)控維持細(xì)胞的氧化還原平衡時的分子機(jī)制不是很清楚。本實驗克隆并分析了哈氏弧菌GR基因序列,其中活性位點和可能的功能位點可以幫助預(yù)測該基因的蛋白質(zhì)結(jié)構(gòu),從分子水平預(yù)測抗氧化系統(tǒng)與細(xì)菌耐藥性的關(guān)系,同時可以進(jìn)一步從構(gòu)建GR功能位點缺失的突變株來研究GR的功能和參與的代謝過程。
因為本實驗室制備的哈氏弧菌GSH抗氧化酶GST和GPx基因疫苗對斜帶石斑魚具有較好的保護(hù)作用,而且有研究在大腸桿菌中表達(dá)的哈氏弧菌GR[28]、SOD[29]純化蛋白對魚也有較好的保護(hù)作用,本實驗經(jīng)克隆病原菌哈氏弧菌GR基因構(gòu)建表達(dá)載體,并得到了該蛋白表達(dá)的最佳條件,28℃誘導(dǎo)時蛋白在上清中的表達(dá)量較多。探索重組活性蛋白的表達(dá)條件,獲得大量活性蛋白,可以從GR蛋白對細(xì)菌的藥物敏感程度的影響來探討GR與細(xì)菌耐藥性的關(guān)系,從而進(jìn)一步研究其在細(xì)菌耐藥性中的功能與作用;同時也可以用于弧菌疫苗的研究,以期從GR角度對弧菌病起到較好的防疫作用。
4 結(jié)論
本研究克隆了哈氏弧菌谷胱甘肽還原酶GR基因,得到1 356 bp堿基序列,將之與pET-32a(+)連接后,轉(zhuǎn)化入E.coli BL21(DE3)中構(gòu)建了重組菌株。表達(dá)條件優(yōu)化結(jié)果為,當(dāng)重組表達(dá)菌生長至OD600=0.4-0.6 時,經(jīng) 0.7 mmol/L 的 IPTG 在 28℃誘導(dǎo) 4 h,表達(dá)出較多的68.9 kD蛋白,為獲得高效表達(dá)的蛋白以檢測其生物學(xué)活性和以后實驗中的應(yīng)用奠定了必要的基礎(chǔ)。
[1] Han-Ching Wang K, Tseng CW, Lin HY, et al. RNAi knock-down of the Litopenaeus vannamei Toll gene(LvToll)significantly increases mortality and reduces bacterial clearance after challenge with Vibrio harveyi[J]. Developmental and Comparative Immunology, 2010,34:49-58.
[2] Vandenberghe J, Verdonck L, Robles Arozarena R, et al. Vibrios associated with Litopenaeus vannamei larvae, postlarvae, broodstock,and hatchery probionts[J]. Appl Environment Microbioltic, 1999,65(6):2592-2597.
[3] Lavilla-Pitogo CRM, Baticados CL, Cruz Lacierda ER, et al. Occurrence of luminous bacterial disease of Penaeus monodon larvae in the Philippines[J]. Aquaculture, 1990, 91(1):1-13.
[4] Pizzutto M, Hirst RG. Classification of isolates of Vibrio harveyivirulent to Penaeus monodon larvae by protein profile analysis and M13 DNA fingerprinting[J]. Diseases of Aquatic Organisms,1995, 21(1):61-68.
[5] Liu PC, Lee KK, Yii KC, et al. Isolation of Vibrio harveyi from diseased kuruma prawns Penaeus japonicus[J]. Current Microbiol, 1996,33(2):129-132.
[6] Huang HH, Liu XL, Xiang JH, Wang P. Selection of Vibrio harveyiresistant Litopenaeus vannameivia a three round challenge selection with a pathogenic strain of V. Harveyi[J]. Fish and Shellfish Immunology, 2013, 35:328-333.
[7] Alvare JD, Austin B, Alvarez AM, Reyes H . Vibrio harveyi:a pathogen of penaeid shrimps and fish in Venezuela[J]. Journal of Fish Diseases, 1998, 21(4):313-316.
[8] 祝璟琳, 王國良, 金珊. 養(yǎng)殖大黃魚病原弧菌多重PCR檢測技術(shù)的建立和應(yīng)用[J]. 中國水產(chǎn)科學(xué), 2009, 16(2):157-163.
[9] Talpur AD, Ikhwanuddin M. Azadirachta indica(neem)leaf dietary effects on the immunity response and disease resistance of Asian seabass, Lates calcarifer challenged with Vibrio harveyi[J]. Fish and Shellfish Immunology, 2013, 34:254-264.
[10] 俞慎, 王敏, 洪有為. 環(huán)境介質(zhì)中的抗生素及其微生物生態(tài)效應(yīng)[J]. 生態(tài)學(xué)報, 2011, 31(15):4437-4446.
[11] Ottaviania D, Bacchiocchi I, Masini L. Antimicrobial susceptibility of potentially pathogenic halophilic vibrios isolated from seafood[J]. International Journal of Antimicrobial Agents, 2001, 18:135-140.
[12] 朱家馨. 銅綠假單胞菌碳青霉烯類抗生素耐藥的分子機(jī)制研究[D]. 廣州:中山大學(xué), 2006.
[13] 吳崢嶸. 雙黃連粉針劑對多重耐藥大腸埃希菌耐藥性影響的機(jī)理研究[D]. 北京:北京中醫(yī)藥大學(xué), 2013.
[14] Albesa I, Becerra MC, Battán PC, Páez PL. Oxidative stress involved in the antibacterial action of different antibiotics[J]. Biochemical and Biophysical Research Communication, 2004,317:605-609.
[15] Aiassa V, Barnes AI, Albesa I. Resistance to ciprofloxacin by enhancement of antioxidant defenses in biofilm and planktonic Proteus mirabilis[J]. Biochemical and Biophysical Research Communications, 2010, 393:84-88.
[16] 樊閱評, 于建春, 余躍, 張琳. 谷胱甘肽的生理意義及其各測定方法比較、評價[J]. 中國臨床營養(yǎng)雜志, 2003, 11(2):136-139.
[17] Sies H. Glutathione and its role in cellular functions[J]. Free Radical Biology and Medicine, 1999, 27(9):916-921.
[18] 鄧治, 王翔, 李德軍. 巴西橡膠樹中谷胱甘肽及其合成代謝途徑關(guān)鍵酶活性的研究[J]. 熱帶農(nóng)業(yè)科學(xué), 2012, 32(12):50-53.
[19] 張亞妮. 谷胱甘肽與銅綠假單胞菌致病性及抗生素抗性的關(guān)系研究[D]. 西安:西北大學(xué), 2009.
[20] 王金慧. 南極冰藻GPx、GST和SAHH基因的克隆、定量分析及原核表達(dá)載體的構(gòu)建[D]. 湛江:廣東海洋大學(xué), 2011.
[21] Loprasert S, Whangsuk W, Sallabhan R, Mongkolsuk S. The unique glutathione reductase from Xanthomonas campestris:Gene expression and enzyme characterization[J]. Biochemical and Biophysical Research Communications, 2005, 331:1324-1330.
[22] Ding SH, Lei M, Lu QT, et al. Enhanced sensitivity and characterization of photosystem II in transgenic tobacco plants with decreased chloroplast glutathione reductase under chilling stress[J]. Biochimica et Biophysica Acta, 2012, 1817:1979-1991.
[23] 沈立新, 魏東芝, 趙哲峰, 等. 谷胱甘肽合成酶系的克隆、測序及表達(dá)[J]. 生物工程學(xué)報, 2001, 17(1):98-100.
[24] 劉瑩. 南極衣藻谷胱甘肽還原酶基因的克隆、表達(dá)和定量分析[D]. 湛江:廣東海洋大學(xué), 2010.
[25] 張菁. GSH對氧自由基介導(dǎo)的細(xì)胞凋亡保護(hù)機(jī)制研究[D].天津:天津醫(yī)科大學(xué), 2002.
[26] 張揚(yáng). 番茄S-亞硝基谷胱甘肽還原酶基因(S1GSNOR)沉默對番茄高溫抗性的影響[D]. 杭州:浙江大學(xué), 2013.
[27] 李平平, 柳生奎, 邱志奇, 安志剛. 谷胱甘肽合成酶基因StGCS-GS的表達(dá)提高大腸桿菌對重金屬的抗性[J]. 森林工程, 2011, 27(1):5-8.
[28] 陶然. 哈維一氏弧菌硫氧還蛋白還原酶在大腸桿菌中的表達(dá)及對大菱鲆免疫保護(hù)作用[D]. 青島:中國海洋大學(xué), 2011.
[29] 辛瑞曉, 易飛, 張蕊, 等. 哈維氏弧菌鐵超氧化物歧化酶基因表達(dá)及免疫作用研究[J]. 水生生物學(xué)報, 2012, 36(2):213-217.
(責(zé)任編輯 李楠)
Cloning and Prokaryotic Expression of Glutathione Reductase Gene in Vibrio harveyi
Zhu Fan1,2,3Ding Yu1,2,3Lu Yishan1,2,3Jian Jichang1,2,3Wu Zaohe2,3,4
(1. Fisheries College,Guangdong Ocean University,Zhanjiang 524088;2. Guangdong Provincial Key Laboratory of Pathogenic Biology and Epidemiology for Aquatic Economic Animals,Zhanjiang 524088;3. Key Laboratory of Diseases Controlling for Aquatic Economic Animals of Guangdong Higher Education Institutions,Zhanjiang 524088;4. Zhongkai University of Agriculture and Engineering,Guangzhou 510225)
The purpose of this study is to clone glutathione reductase(GR)gene from Vibrio harveyi, construct the prokaryotic expression vector of it, and obtain the corresponding expressed protein. Digested GR and pET-32a(+)were cut with the double enzymes BamH I and Xho I, then ligated with T4 ligase to construct the recombinant plasmid pET-GR. Then pET-GR was transformed into Escherichia coli BL21(DE3), which was induced by IPTG, and their expressions were analyzed by SDS-PAGE. An approximately 68.9 kD exogenous protein was observed on the SDS-PAGE. The optimal expression condition for the recombinant plasmid pET-GR was that the recombinant E. coli BL21(DE3)was induced for 4 h at 28℃ by 0. 7 mmol/L of IPTG and it expressed in E. coli as the inclusion bodies. The conclusion of the study is that GR gene from V. harveyi can efficiently express in E. coli.
Vibrio harveyi;glutathione reductase;gene cloning;prokaryotic expression
10.13560/j.cnki.biotech.bull.1985.2015.06.029
2014-10-22
廣東省教育廳高等學(xué)校高層次人才項目
朱帆,女,碩士研究生,研究方向:水產(chǎn)經(jīng)濟(jì)動物病害;E-mail:15622071756@163.com
丁燏,男,博士,教授,研究方向:水產(chǎn)經(jīng)濟(jì)動物免疫學(xué)及病害控制;E-mail:dingy@gdou.edu.cn
