中華絨螯蟹性腺特異表達蛋白EsSOX21b-like的原核表達與抗體制備
2015-10-27 01:25:24楊國翠崔崢邱高峰
生物技術通報 2015年6期
關鍵詞:檢測
楊國翠 崔崢 邱高峰
(上海海洋大學 省部共建水產種質資源發掘與利用教育部重點實驗室,上海 201306)
中華絨螯蟹性腺特異表達蛋白EsSOX21b-like的原核表達與抗體制備
楊國翠 崔崢 邱高峰
(上海海洋大學 省部共建水產種質資源發掘與利用教育部重點實驗室,上海 201306)
Sox(SRY-related HMG-box)基因編碼一類重要的轉錄因子,其產物都具有一個HMG基序保守區。之前我們在中華絨螯蟹分離鑒定了一個只在性腺中特異表達轉錄本EsSox21b-like,為了驗證EsSOX21b-like蛋白是否也在中華絨螯蟹性腺中特異表達,本研究原核表達了EsSOX21b-like蛋白,即將EsSox21b-like基因序列克隆到pGEX-2T表達載體,構建重組表達質粒pGEX-2T-EsSox21b-like,轉化大腸桿菌BL21,IPTG誘導融合表達,經SDS-PAGE分析表明,融合蛋白主要以包涵體形式存在,分子量約為54 kD。利用Ni柱親和純化融合蛋白后免疫家兔,制備了多克隆抗體,Western blotting檢測表明該抗體能特異地識別性腺組織中EsSOX21b-like蛋白,而在其他組織未檢測到其表達,目的蛋白分子量約為28 kD,在精巢組織中的表達量比在卵巢中高,該結果暗示EsSOX21b-like可能在性腺發育過程中起重要調控作用。
中華絨螯蟹;EsSOX21b-like蛋白;原核表達;抗體制備
Sox(SRY-related HMG-box)基因是一類SRY(sex-determining region of Y chromosome)相關基因,編碼一系列轉錄因子,參與性別決定與分化、骨組織發育、血細胞生成、神經系統發育、晶狀體發育等多種早期胚胎發育過程[1]。SRY基因是哺乳動物雄性性別決定基因,對睪丸的發育有著決定性的影響,它首先在人類Y染色體上發現,是Sox基因家族的第一個成員[2]。其產物含有一個HMG盒[3],能夠與DNA特異性結合,調控其轉錄。之后研究發現了許多與SRY相關的Sox基因,Sox3、Sox5、Sox6、Sox8、Sox9、Sox17等都是參與性別分化的重要基因。Sox9基因雄鼠胚胎精巢中表達,具有促使精巢支持細胞分化的作用[4;5]。在小鼠中Sox9基因的缺失導致XY向XX的性反轉現象[6]。Sox3是與SRY同源性最高的基因[7],但是并不直接參與睪丸的分化和發育,Graves[7]認為在雌性中,Sox3表達,抑制了Sox9的表達;在雄性中SRY抑制Sox3,Sox9得以表達,從而決定睪丸的形成。Sox8也是哺乳動物睪丸決定途徑相關的基因,能夠促進Sox9在睪丸形成中的作用。但是,Sox8基因缺陷的小鼠睪丸發育正常[8]。另外,Sox5、Sox6在小鼠的減數分裂后的生殖細胞中表達,尤其是在精細胞階段[9];Sox17則在減數分裂前的精母細胞中表達[10]。
水產動物中也發現存在Sox基因,參與性別決定與分化過程。虹鱒中Sox24作為轉錄因子在卵子發生過程起作用[11]。黃鱔Sox17在精巢、卵巢和兩性腺中表達,特別是在卵巢、兩性腺和精巢片層及生精細胞中含量更高,可以推測Sox17對黃鱔自然性反轉過程中的性腺發育有重要作用[12]。鱸魚在150 d性別分化開始時Sox17在雌雄性腺中均表達,在250 d能觀察到明顯的性別差異時,雌魚Sox17表達量明顯高于雄魚,并且其RNA水平與芳香化酶相關,可能參與卵巢發育[13]。另外鱸魚中還發現存在Sox19與卵巢分化相關[14]。但是在甲殼類動物中Sox基因是否參與性別決定與分化過程目前尚不清楚。
本研究組之前在中華絨螯蟹(Eriocheir sinensis)克隆得到只在性腺中特異表達轉錄本EsSox21blike[15],并在RNA水平分析了Sox基因的組織表達,結果顯示EsSox21b-like基因在精巢、卵巢中均有表達,推測EsSox21b-like可能參與性腺的發育。為驗證EsSox21b-like基因在蛋白水平的表達及作用,本研究組構建了EsSOX21b-like蛋白原核表達質粒,并在大腸桿菌中高效表達重組蛋白,純化重組蛋白,制備EsSOX21b-like多克隆抗體,并對抗體進行免疫學鑒定,旨為進一步研究中華絨螯蟹性別決定與分化機制提供強有力的工具。
1 材料與方法
1.1 材料
中華絨螯蟹購自當地水產市場。大腸桿菌(Escherichia coli)DH5α、BL21(DE3)感受態細胞、原核表達載體pGEX-2T為本實驗室保存,Trizol購自Invitrogen。First strand cDNA synthesis kit,Ex-Taq DNA聚合酶和T4 DNA連接酶購自TaKaRa。BamHⅠ和EcoRⅠ購自Promega。PCR純化產物試劑盒、低分子蛋白Marker、預染蛋白Marker、SDSPAGE各組分購自上海天根生化科技公司。His-select Spin Columns純化試劑盒、His標簽抗體購自Sigma。辣根過氧化物標記的山羊抗鼠、山羊抗兔IgG購自艾比瑪特。DAB顯色劑購自武漢博士德生物工程有限公司。
1.2 方法
1.2.1 原核表達載體的構建 使用Trizol Reagent提取中華絨螯蟹性腺總RNA,經DNaseⅠ處理后,按First strand cDNA synthesis kit說明書進行反轉錄,獲得的cDNA作為模板,擴增EsSox21b-like基因。根據本實驗室已克隆到的中華絨螯蟹EsSox-like基因全長cDNA序列,利用Primer 5軟件設計特異引物(表1),擴增EsSox-like編碼區序列。同時,該對引物的5'端分別加入BamHⅠ和EcoRⅠ酶切位點,另外,在下游引物緊鄰終止密碼子的3'端引入編碼6個組氨酸核酸序列。反應條件為:94℃預變性3 min,94℃ 30 s、58℃ 30 s、72℃ 1.5 min,共30個循環;最后72℃延伸10 min。PCR產物用柱純化回收與pGEX-2T分別進行BamHⅠ和EcoRⅠ雙酶切(37℃,2 h),酶切產物純化,16℃連接過夜。連接產物轉化大腸桿菌DH5α,菌落PCR篩選陽性克隆,擴大培養陽性克隆提取質粒(天根質粒抽提試劑盒),經BamHⅠ和EcoRⅠ雙酶切鑒定和測序(上海美吉生物),確定插入片段序列正確性。
1.2.2 工程菌的誘導表達 重組質粒轉化大腸桿菌BL21(DE3)。挑選陽性菌落接種于液體培養基中,37℃,振蕩培養過夜。次日按1∶100的比例在含有Amp+(終濃度為60 μg/mL)的液體培養基中擴大培養至OD595介于0.4-0.6之間,加IPTG誘導表達。離心收集菌體(5 000 r/min,5 min 4℃),用1×PBS重懸菌體,冰浴下超聲波破碎(超聲時間4 s,間歇時間4 s,保護溫度24℃,總時間90次)。12 000 r/min 4℃離心10 min,沉淀用含8 mol/L尿素的1×PBS重懸,分別取等量的上清和重懸液SDS-PAGE(5%濃縮膠,12%分離膠)電泳,考馬斯亮藍R250染色過夜,充分脫色后觀察并分析結果。
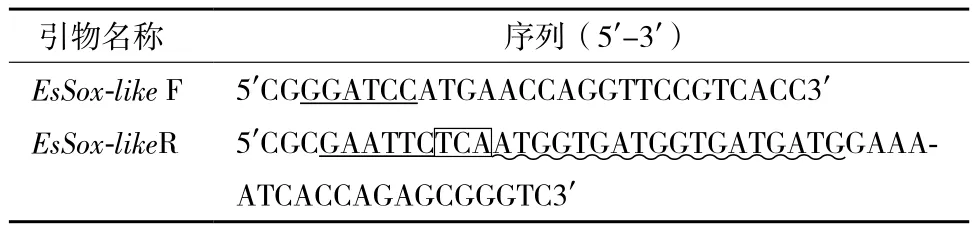
表1 構建EsSox21b-like原核表達載體使用的PCR特異引物
1.2.3 Western blotting檢測融合蛋白 重組蛋白經SDS-PAGE分析后,使用半干轉移膜將重組蛋白轉移到硝酸纖維NC膜上。含10%小牛血清的TTBS室溫封閉1 h,1∶1 000稀釋的His標簽單抗為一抗4℃孵育過夜,TTBS洗滌3次,1∶3 000稀釋的HRP標記山羊抗兔的IgG為二抗,室溫孵育2 h,TTBS緩沖液洗滌3次,去除非特異性結合,用DAB顯色液(DAB 15 mg、甲醇 5 mL、30%雙氧水15 μL)顯色。
1.2.4 EsSOX21b-like多克隆抗體的制備及檢測 用His-select Spin Columns純化試劑盒純化的重組蛋白經弗氏佐劑乳化后免疫2只成年健康的大白兔。初次免疫后,每隔2周加強免疫1次,第5次免疫后8 d處死兔子。取全血分離血清,抗原親和純化血清。1.2.5 組織蛋白的提取以及抗血清的檢測 分別取50 mg精巢、卵巢、心臟、肝胰腺、肌肉、胸神經節和鰓組織與2 mL離心管中,加入1 mL組織蛋白抽提液(博士德),于冰上充分勻漿后4℃ 12 000 r/min離心10 min,取上清。SDS-PAGE電泳,轉膜,適度稀釋的EsSOX21b-like多克隆抗體為一抗,HRP標記的山羊抗兔IgG為二抗進行Western檢測。
2 結果
2.1 原核表達載體的構建
以中華絨螯蟹性腺cDNA為模板,PCR擴增,瓊脂糖凝膠電泳得到一條約800 bp的條帶,與目的片段大小基本一致。將其克隆至原核表達載體pGEX-2T中,得到重組質粒。BamHⅠ和EcoRⅠ雙酶切重組質粒,瓊脂糖凝膠電泳(圖1)顯示在800 bp處有條帶。同時測序結果也顯示,克隆得到的序列與EsSox21b-like ORF一致,并且在其終止密碼子前成功的引入了編碼6個組氨酸的核酸密碼子序列。將重組質粒命名為pGEX-2T-EsSox21b-like。
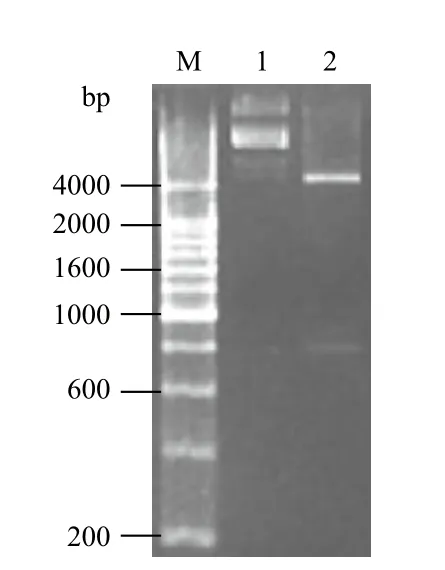
圖1 重組質粒pGEX-EsSox21b-like雙酶切圖
2.2 重組蛋白的表達及檢測
重組質粒pGEX-2T-EsSox21b-like轉化大腸桿菌BL21(DE3)。IPTG誘導,超聲破碎,離心分離沉淀與上清,SDS-PAGE電泳分析。與未經IPTG誘導的工程菌相比,誘導組的沉淀中有一條明顯加粗的條帶(圖2,箭頭),分子量約54 kD,這與EsSox21b-like基因編碼的蛋白分子量(28 kD)和pGEX-2T上攜帶的GST標簽蛋白(圖2,箭頭)分子量(26.3 kD)總和相當。在分離的上清中則未檢出相應的條帶。
為了證實重組蛋白的表達,使用His標簽抗體做為一抗進行Western blotting分析,免疫檢測發現在54 kD處有特異性條帶(圖3)。說明經過誘導后能成功的獲得融合蛋白,且該融合蛋白主要以包涵體的形式存在。

圖2 pGEX-2T-EsSox21b-like融合蛋白表達產物的SDSPAGE檢測
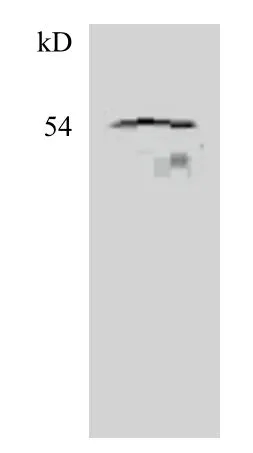
圖3 His抗體檢測EsSOX21b-like重組蛋白的Western blotting分析
2.3 EsSOX21b-like抗體的制備與免疫鑒定
純化后的重組蛋白分5次注射免疫2只家兔,制備EsSO21b-like多克隆抗體。第5次免疫后采血,抗血清用包被有重組EsSOX21b-like蛋白的酶標板進行間接ELISA反應,每孔用100 μL濃度為1 μg/mL的抗原包被,用酶標儀讀取450 nm下的光吸收值,吸光度(OD值),結果(表2)顯示2只兔在第5次免疫后,其血清分別在1∶8 000和1∶16 000的稀釋度下,OD值大于1.000,說明制備的多抗效價高。
使用制備的EsSOX21b-like多抗檢測EsSOX21blike蛋白的組織分布情況。Western blotting結果(圖4)顯示中華絨螯蟹精巢、卵巢的蛋白提取物中能夠檢測到特異性條帶,且該特異性條帶所對應的蛋白分子量與軟件預測的EsSOX21b-like蛋白分子量(28 kD)大小相等。而心臟、肝胰腺、肌肉、胸神經節和鰓組織總蛋白中則不能檢測到特異性條帶。該結果說明制備的EsSOX21b-like多克隆抗體能有效的與目的蛋白結合,EsSOX21b-like蛋白在性腺組織中特異表達,并且在精巢組織中表達量高于卵巢。

圖4 EsSOX21b-like 抗體檢測不同組織總蛋白

表2 ELISA法檢測抗體效價
3 討論
本研究通過IPTG誘導、超聲波破碎工程菌,在沉淀中檢測到融合表達的目的蛋白,并且用Hisselect Spin Columns純化獲得的融合蛋白能夠有效地免疫家兔獲得效價高、特異性強的EsSOX21b-like蛋白的多克隆抗體。相反,在離心后的上清液中沒有檢測到可溶性的融合蛋白,說明表達的蛋白以包涵體形式存在。與具有活性的可溶性蛋白相比包涵體蛋白具有相同的一級結構,不影響抗體制備,只是由于肽鏈無法正確折疊形成二、三級結構而不具有生物活性[16]。包涵體蛋白經過變性、洗滌、復性、純化等一系列處理過程最終可以轉變為活性蛋白[17]。之前許多研究表明,原核表達也可以獲得有活性的可溶蛋白[18-21],其中IPTG的誘導至關重要,可以通過減少IPTG的誘導量和誘導時間控制目的蛋白表達量,避免包涵體的形成;溫度對蛋白的可溶性也有很大的影響,如范丙友[21]通過降低溫度等成功獲得了可溶性的香豆酸:輔酶A連接酶。除此之外,以大腸桿菌為宿主細胞進行原核表達時,還可以在培養基中加入一些能促進重組蛋白質可溶性表達的生長添加劑,如高濃度的多醇類、蔗糖或非代謝糖(阻止分泌到周質的蛋白質聚集反應)、乙醇(誘導熱休克蛋白的表達)、低分子質量的巰基或二硫化合物(影響細胞周質的還原態,從而影響二硫鍵的形成)和NaC1等來增加融合蛋白的溶解度[22]。后續我們擬采用以上各種實驗方法進行直接誘導表達以其獲得具有活性的目的蛋白,為研究EsSOX21b-like 蛋白生物學功能奠定必不可少的物質基礎。
Sox21基因是SoxB基因亞族中的重要一員,在脊椎動物與無脊椎動物中均有發現。在小鼠,Sox21在未成熟神經系統腦室區表達,而在成熟腦內其表達則限制在腦室下區,進一步的研究證實Sox21在小鼠神經系統發育過程能抑制神經元分化[23]。另外,Sox21在小鼠的耳蝸中也有表達,且Sox21缺陷型小鼠表現出輕度聽力障礙[24]。在斑馬魚,Sox21存在Sox21a與Sox21b兩種亞型,Sox21a在腦、皮膚、腸及卵巢中表達,而Sox21b除了影響性腺的正常發育外對晶狀體的正常發育也會起到一定的調控作用[25,26]。Sox21b基因在蜂王卵巢管中的滋養細胞及其卵子中也有較強表達,在蜜蜂卵巢發育及成熟過程中發揮其功能[27]。蜜蜂在分類系統上與中華絨螯蟹同屬于節肢動物門,其親緣關系更加緊密,故彼此相同的基因在表達特征與功能上應該相近或相同。之前我們的研究結果表明,中華絨螯蟹EsSox21blike基因的mRNA僅局限在精巢和卵巢表達,暗示在精子的發生及卵細胞的發育過程中發揮作用。本研究在蛋白水平進一步證明EsSOX21b-like在性腺中特異表達,且在精巢組織中表達量明顯高于卵巢,推測該蛋白可能是精巢發育必不可少的因子。
4 結論
本研究以中華絨螯蟹性腺為模板,設計特異性引物,擴增EsSox21b-like的開放閱讀框,克隆至pGEX-2T表達載體,在大腸桿菌中表達EsSOX21blike重組蛋白。SDS-PAGE結果顯示,EsSOX21b-like重組蛋白主要以包涵體形式存在。大量表達重組蛋白,親和層析純化后免疫家兔,得到高效價的多克隆抗體。Western blotting檢測中華絨螯蟹不同組織EsSOX21b-like蛋白表達情況,結果顯示EsSOX21blike蛋白在性腺中特異表達,尤其在精巢中表達量高,而在其他組織中不表達,推測EsSOX21b-like在性腺發育中具有重要的調控作用。
[1] 常重杰, 杜啟艷, 邵紅偉. Sox 基因家族研究的新進展[J]. 遺傳, 2002, 24(4):470-476.
[2] Berta P, Hawkins JB, Sinclair AH, et al. Genetic evidence equating SRY and the testis-determining factor[J]. Nature, 1990, 348(6300):448-450.
[3] Bowles J, Schepers G, Koopman P. Phylogeny of the SOX family of developmental transcription factors based on sequence and structural indicators[J]. Developmental Biology, 2000, 227(2):239-255.
[4] Da Silva SM, Hacker A, Harley V, et al. Sox9 expression during gonadal development implies a conserved role for the gene in testis differentiation in mammals and birds[J]. Nature Genetics, 1996,14(1):62-68.
[5] Fr?jdman K, Harley VR, Pelliniemi LJ. Sox9 protein in rat Sertoli cells is age and stage dependent[J]. Histochemistry and Cell Biology, 2000, 113(1):31-36.
[6] Chaboissier MC, Kobayashi A, Vidal VI, et al. Functional analysis of Sox8 and Sox9 during sex determination in the mouse[J]. Development, 2004, 131(9):1891-1901.
[7] Foster JW, Graves J. An SRY-related sequence on the marsupial X chromosome:implications for the evolution of the mammalian testis-determining gene[J]. Proceedings of the National Academy of Sciences, 1994, 91(5):1927-1931.
[8] Kiefer JC. Back to basics:Sox genes[J]. Developmental Dynamics, 2007, 236(8):2356-2366.
[9]Newton J, Cohen-Barak O, Hagiwara N, et al. Mutations in the human orthologue of the mouse underwhite gene underlie a new form of oculocutaneous albinism, OCA4[J]. The American Journal of Human Genetics, 2001, 69(5):981-988.
[10]Sekiya I, Koopman P, Tsuji K, et al. Dexamethasone enhances SOX9 expression in chondrocytes[J]. Journal of endocrinology,2001, 169(3):573-579.
[11]Kanda H, Kojima M, Miyamoto N, et al. Rainbow trout Sox24, a novel member of the Sox family, is a transcriptional regulator duringoogenesis[J]. Gene, 1998, 211(2):251-257.
[12]Wang R, Cheng H, Xia L, et al. Molecular cloning and expression of Sox17 in gonads during sex reversal in the rice field eel, a teleost fish with a characteristic of natural sex transformation[J]. Biochemical and Biophysical Research Communications, 2003, 303(2):452-457.
[13] Navarro-Martín L, Galay-Burgos M, Sweeney G, et al. Different sox17 transcripts during sex differentiation in sea bass, Dicentrarchus labrax[J]. Molecular and Cellular Endocrinology, 2009,299(2):240-251.
[14] Navarro-Martín L, Galay-Burgos M, Piferrer F, et al. Characterisation and expression during sex differentiation of Sox19 from the sea bass Dicentrarchus labrax[J]. Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology, 2012, 163(3):316-323.
[15] 亓海燕, 邱高峰. 一個在中華絨螯蟹性腺中特異表達的 Sox 基因 HMG 盒區的克隆與鑒定[J]. 自然科學進展, 2009, 19(3):279-284.
[16] Hoffmann F, Heuvel JVD, Zidek N, et al. Minimizing inclusion body formation during recombinant protein production in Escherichia coli at bench and pilot plant scale[J]. Enzyme and Microbial Technology, 2004, 34(3):235-241.
[17] 萬雪, 王磊, 寧官保. 包涵體及其復性研究概況[J]. 畜牧獸醫科技信息, 2005, (2):13-15.
[18] 邵恩斯, 林莉, 關雄. 蘇云金桿菌 Cry2Ab 可溶蛋白的原核表達及多克隆抗體的制備[J]. 農業生物技術學報, 2013, 21(1):106-111.
[19]劉忠淵, 張富春, 王蕓, 等. 赤翅甲抗凍蛋白基因的原核表達及蛋白生物活性檢測[J]. 昆蟲學報, 2005, 48(2):179-183.
[20]孫健偉, 韓洪彥, 徐文琳, 等. 大鼠腦紅蛋白(NGB)可溶性原核表達和單克隆抗體制備及鑒定[J]. 中國生物化學與分子生物學報, 2003, 19(6):751-756.
[21]范丙友, 胡詩宇, 陸海, 等. 毛白楊 4-香豆酸:輔酶 A 連接酶可溶性原核表達及活性檢測[J]. 北京林業大學學報, 2006,28(2):1-8.
[22]艾恒, 莫書榮, 魯波. 包涵體研究進展[J]. 中國醫學文摘(內科學), 2006, 2:006.
[23]Ohba H, Chiyoda T, Endo E, et al. Sox21 is a repressor of neuronal differentiation and is antagonized by YB-1[J]. Neuroscience Letters, 2004, 358(3):157-160.
[24]Hosoya M, Fujioka M, Matsuda S, et al. Expression and function of Sox21 during mouse cochlea development[J]. Neurochemical Research, 2011, 36(7):1261-1269.
[25]Lan X, Wen L, Li K, et al. Comparative analysis of duplicated sox21 genes in zebrafish[J]. Development, Growth & Differentiation,2011, 53(3):347-356.
[26]Pauls S, Smith SF, Elgar G. Lens development depends on a pair of highly conserved Sox21 regulatory elements[J]. Developmental Biology, 2012, 365(1):310-318.
[27]Wilson MJ, Dearden PK. Evolution of the insect Sox genes[J]. BMC Evolutionary Biology, 2008, 8(1):120.
(責任編輯 李楠)
Prokaryotic Expression,Antibody Preparation of Gonad-specific Protein EsSOX21b-like of the Chinese Mitten Crab Eriocheir sinensis
Yang Guocui Cui Zheng Qiu Gaofeng
(Key Laboratory of Aquatic Genetic Resources and Utilization Certificated by Ministry of Agriculture and Shanghai Ocean University,Shanghai 201306)
The Sox(SRY-related HMG-box)is a large family of genes which encode transcription factors with high-mobility-group DNA binding domain related to the SRY(sex determining region Y). In previous studies we isolated and identified EsSox21b-like transcripts exclusively expressed only in gonads of the Chinese mitten crab(Eriocheir sinensis), this study is to verify whether EsSOX21b-like protein is also specifically expressed in the gonad. Gene EsSox21b-like was cloned to expression vector pGEX-2T, the recombinant prokaryotic expression plasmid pGEX-2T-EsSox21b-like was constructed, then transferred into host Escherichia coli BL21, and induced by IPTG. SDS-PAGE analysis revealed that the fusion protein was expressed as inclusion body with the molecular weight of 54 kD. The recombinant proteins were subsequently purified by Ni+affinity chromatography. Then the purified protein was used to immunize rabbits for preparation of the EsSOX21b-like antibody. Western blotting detection showed the antibody specifically recognized the EsSOX21b-like protein in the gonad, with molecular weight of 28 kD,whereas no expression was detected in other tissues. The expression level of the target protein was much higher in testis than that in ovary. This result implied that the EsSOX21b-like protein could have a regulatory role in the gonad development.
Chinese mitten crab(Eriocheir sinensis);EsSOX21b-like protein;prokaryotic expression;antibody preparation
10.13560/j.cnki.biotech.bull.1985.2015.06.030
2014-09-16
國家自然科學基金項目(31272655),國家科技支撐計劃項目(2012BAD26B04),上海市科委重點基礎科研基金項目(11JC140 4600),上海高校水產學一流學科建設項目(B5005120001)
楊國翠,女,碩士研究生,研究方向:中華絨螯蟹生殖與發育;E-mail:554025117@qq.com
邱高峰,男,教授,博士生導師,研究方向:蝦蟹分子遺傳與繁殖;E-mail:gfqiu@shou.edu.cn
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
