轉基因小鼠特異性免疫效應淋巴細胞影響乙型肝炎病毒復制的體外研究
2015-11-22 07:44:34宋紅麗鄭衛萍楊洋劉濤吳本娟付楠楠張友成沈中陽
實用器官移植電子雜志 2015年4期
宋紅麗,鄭衛萍,楊洋,劉濤,吳本娟,付楠楠,張友成,沈中陽
(1.天津市第一中心醫院器官移植中心,天津市器官移植重點實驗室,天津 300192;2.天津市兒童醫院,天津 300074)
目前肝移植術后預防乙型肝炎(乙肝)復發的常用方案為小劑量乙肝免疫球蛋白(HBIG)聯合核苷類似物,并取得了良好的效果[1]。但是長期應用HBIG和核苷類似物藥物費用很高,對患者和社會來說負擔沉重;同時作為生物制品的HBIG日益短缺,使部分患者不能堅持有效預防乙肝復發的方案,而且長期使用核苷類似物容易導致病毒耐藥[2]。術后乙肝疫苗的應用,使患者看到了停用抗病毒藥物及HBIG的可能性,但是目前由于移植術后免疫抑制劑的使用,使乙肝疫苗治療有效性較差[3-4]。如果能夠尋找出經濟有效的新方法,從而使乙肝患者移植術后徹底停用抗病毒藥物,將在該領域取得突破,獲得重大的經濟和社會效應。
肝移植是各類終末期肝臟疾病的主要治療手段,在中國,其中70%~80%為乙型肝炎病毒(HBV)導致[5],患者通常在等待肝移植手術時機的同時接受抗病毒治療以降低體內HBV DNA的復制水平[6],同時術前HBV DNA的水平也是術后HBV復發的重要預測指標。因此,提高患者主動免疫力、降低等待移植患者體內HBV DNA水平,已成為預防術后乙肝復發的重要目標。樹突狀細胞(DC)是機體內能激活初始T細胞主要的抗原呈遞細胞(APC),其強大的抗原呈遞能力是連接機體固有免疫和適應性免疫的橋梁[7],因此,DC在各類病毒感染、腫瘤免疫中扮演著重要角色[8]。DC作為體內功能最強大的APC,可以通過對抗原的攝取、加工處理呈遞給T細胞啟動相應性免疫應答,從而增強患者體內特異性殺傷HBV的能力,提高患者的主動免疫力。T細胞亞群的調節紊亂,也是HBV在體內持續復制的主要原因[9]。本研究主要通過觀察轉HBV基因小鼠DC刺激自體淋巴細胞對體外HBV復制,尤其對cccDNA的作用,來評價HBV感染后特異性免疫功能的變化及其與HBV復制水平的關系。
1 材料與方法
1.1 主要儀器及試劑:FACS Calibur流式細胞儀(美國BD公司);酶標儀(美國Biotek公司);DC生成培養基DXF(德國PromoCell公司);淋巴細胞分離液(天津灝洋生物制品科技有限責任公司);胎牛血清(FBS,奧地利PAA公司);干擾素(IFN)-γ-APC、白細胞介素(IL)-4-藻紅蛋白(PE)單抗(美國BD Pharmingen公司);CD80-APC/CD86-FITC/CD11C-FITC MHC-Ⅱ-PE/CD83-PE(美國BD公司);乙肝核心抗原/乙肝表面抗原(HBcAg/HBsAg)(英國 Peprotech公司);IL-10、IL-2和IFN-γ的ELISA試劑盒(中國Biovalue公司)。
1.2 研究對象:HBV全基因轉基因C57 BL/6J小鼠[10]由上海南方模式生物科技發展有限公司提供;HepG 2.2.15細胞株由北京軍區302醫院贈送。
1.3 實驗分組:A組為淋巴細胞(LC)+未成熟DC組;B組為HepG 2.2.15 +未成熟DC + LC組;C組為LC +負載HBV相關抗原刺激的成熟DC組(成熟DC組);D組為HepG 2.2.15 +成熟DC + LC組;E組為HepG 2.2.15組。
1.4 實驗方法
1.4.1 淋巴細胞的獲取:無菌制備C57BL/6J小鼠脾和胸腺的細胞懸液,將淋巴細胞懸液移到離心管中的細胞分離液(Percoll)淋巴細胞分離液上面,使其具有明顯分層。以2 000 r/min離心20分鐘,吸取中間白膜層,再以1 000 r/min離心8分鐘,棄去上清。最后用含0.05%吐溫-20的pH 7.4的磷酸鹽緩沖液(PBS)洗滌,計數后加入培養液轉移到培養板中共培養。
1.4.2 提取HBV轉基因鼠的DC進行培養及形態學觀察:取6~8周(體重18~22 g)的HBV全基因轉基因C57BL/6小鼠,斷頸椎處死,按以下步驟進行操作:取股骨及脛骨骨髓,PBS沖洗,離心棄上清,收集骨髓細胞;加入5 ml已預熱37℃的紅細胞裂解液,除去紅細胞;加入5倍體積RPMI 1640培養液稀釋,離心(1 500 r/min,5分鐘);用含100 ml/L 胎牛血清(FCS)的RPMI 1640的完全培養液重懸,調整細胞密度為1×109/L,加入到6孔板中,每孔2 ml;37℃、50 ml/L CO2培養箱中培養。加入新鮮RPMI 1640完全培養液,同時添加rGM-CSF(終濃度為 10 ng/ml)和 rIL-4(1 ng/ml)于骨髓細胞中;第2天去除未貼壁的細胞;第4天半量換液,并補充細胞因子rGM-CSF和rIL-4(濃度同前);第6天成為不成熟的DC,倒置顯微鏡下觀察細胞形態和數量變化。
1.4.3 成熟的DC獲取:將培養至第8天的DC分別用HBV相關蛋白HBsAg(19.6 μg/ml)和HBcAg(10 μg/ml)誘導,第10天細胞即轉為帶有HBV的抗原成熟的DC。通過流式細胞儀檢測DC表面標志物CD80、CD86、CD83等的表達。
1.4.4 免疫效應細胞(IEC)的制備及檢測:成熟的DC刺激自體淋巴細胞(DC︰淋巴細胞=1︰10),經過10余天的培養及活化,制備成IE細胞(CD8+為主)并進行不斷的分裂與增殖。應用流式細胞儀測定淋巴細胞表面標志物。
1.4.5 ELISA法檢測上清中的IL-10/IFN-γ/IL-2的分泌:按照試劑盒說明書步驟來操作,顯色后在450 nm波長下檢測吸光度(A)值。
1.4.6 IEC與HepG 2.2.15細胞進行混合培養,并檢測免疫細胞對HepG 2.2.15細胞中cccDNA及HBV DNA水平的影響:免疫效應細胞與HepG 2.2.15細胞混合于6孔板,效靶比為1︰10,在37℃、50 ml/L CO2培養箱中共培養24、48和72小時,每組設3個空白孔。收集混合細胞上清及細胞,進行cccDNA及HBV DNA水平的檢測。
1.4.7 流式細胞儀檢測DC表面分子:用流式細胞儀對DC表面標志物CD80/CD83/CD86、MHCⅡ/CD11C進行檢測,按試劑盒說明書進行操作。
1.4.8 生物化學方法檢測上清液的肝功能指標。
1.4.9 實時熒光定量PCR方法檢測血清及細胞內HBV DNA:使用ABI 7500實時定量PCR儀(Applied Biosystems),操作按PCR試劑盒說明書進行(上海Kehua公司)。
1.4.10 HBV共價閉合環狀DNA(cccDNA)檢測:采用實時定量PCR方法檢測細胞內的cccDNA,具體方法見參考文獻[11]。
1.5 統計學處理:采用SPSS 16.0軟件進行統計分析。計量資料以均數±標準差(±s)表示,組間比較采用單因素方差分析,P<0.05為差異有統計學意義。采用GraphPad Prism5軟件做圖。
2 結 果
2.1 檢測HBV全基因轉基因C57BL/6J小鼠血清的HBsAg、HBeAg、HBV DNA均陽性,而肝細胞內的cccDNA陰性。肝功能、腎功能等正常。
2.2 成功獲取轉HBV基因鼠骨髓中的DC細胞
2.2.1 DC形態學變化:第1天,鏡下觀察可見貼壁的圓形單核細胞,細胞體積小,數量多,無突觸;第3天,細胞體積逐漸變大,由規則的圓形向長條狀進展;第6天,細胞體積明顯增大,細胞表面伸出短的突觸(圖1a)。按第6天DC細胞是否負載HBsAg和HBcAg抗原肽分為負載抗原成熟組和未負載抗原未成熟組,第8天負載抗原成熟組的細胞大量懸浮,小而圓的單核細胞逐漸消失,細胞表面都伸出兩個或多個突觸,細胞體積大,突觸明顯,未分化的單核細胞較少(圖1b)。圖1c、1d顯示出由未成熟到成熟的DC表型的典型變化。
2.2.2 DC表面標志的變化(表1):與培養6天未成熟DC相比,負載抗原成熟的DC表面標志物CD11c/MHC-Ⅱ、CD83、CD80、CD86的表達在成熟DC中均明顯增高(均P<0.05),說明在體外環境HBV相關抗原可以刺激體外DC的成熟。
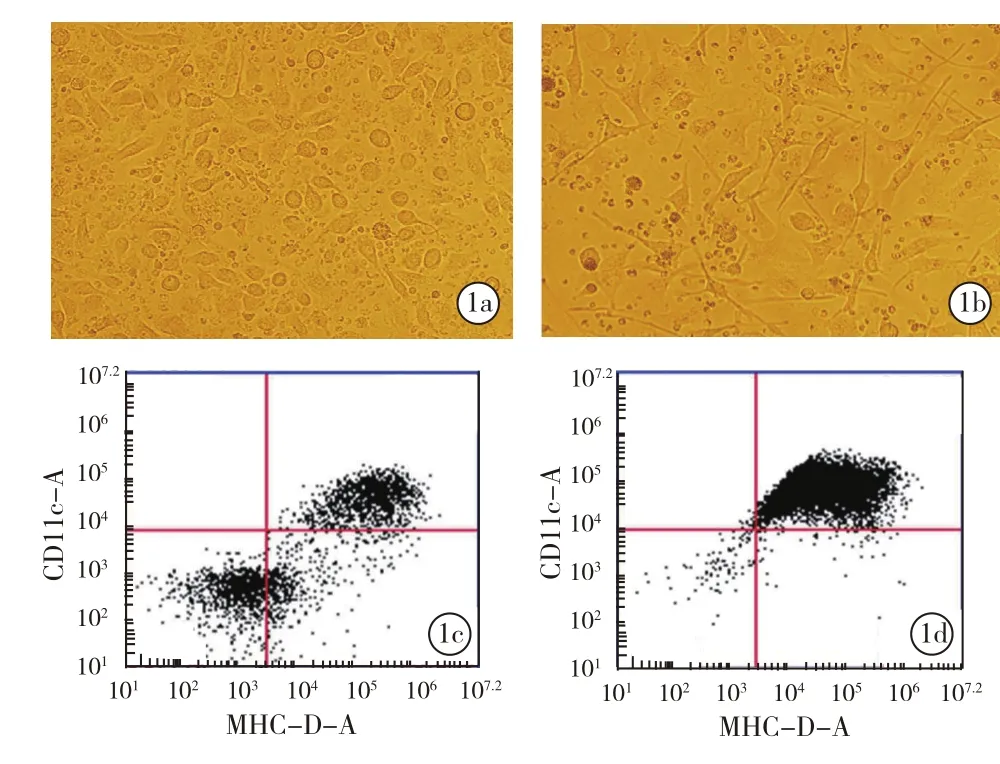
圖1 未成熟與成熟DC形態比較(a、b均為低倍放大)
表1 未成熟DC與成熟DC表型變化比較(±s)
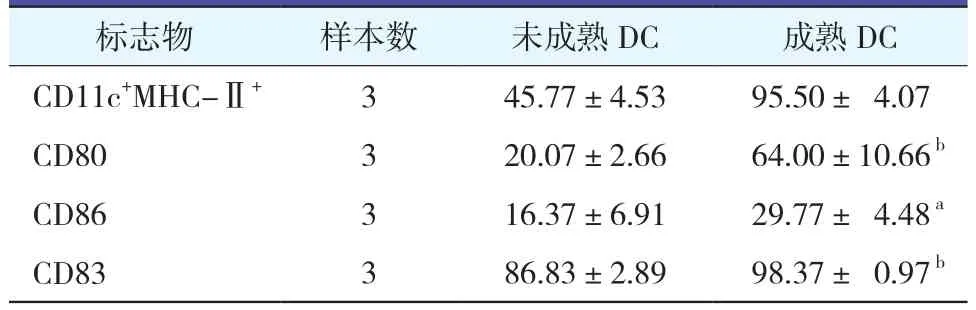
表1 未成熟DC與成熟DC表型變化比較(±s)
注:與未成熟DC組比較,aP<0.05,bP<0.01
標志物 樣本數 未成熟DC 成熟DC CD11c+MHC-Ⅱ+ 3 45.77±4.53 95.50± 4.07 CD80 3 20.07±2.66 64.00±10.66 b CD86 3 16.37±6.91 29.77± 4.48 a CD83 3 86.83±2.89 98.37± 0.97 b
2.3 特異性效應性淋巴細胞對HepG 2.2.15細胞的肝功能及上清HBV DNA的影響:
2.3.1 D組與B組細胞上清液HBV DNA變化 (表2;圖2):D組在24、48和72小時細胞上清HBV DNA均較E組明顯下降(均P<0.01),B組細胞上清HBV DNA水平也較E組有所下降,但差異無統計學意義(均P>0.05)。說明特異性的成熟DC刺激的淋巴細胞能有效地抑制體外HBV的釋放,但與B組相比,D組作用更顯著。
表2 各組不同時間點細胞上清液HBV DNA變化(±s)
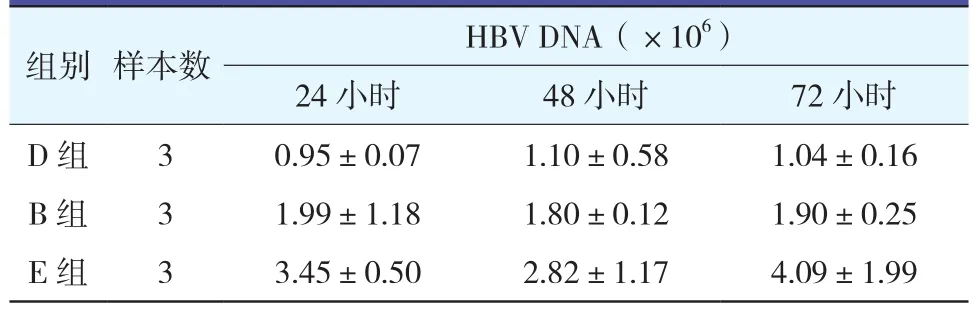
表2 各組不同時間點細胞上清液HBV DNA變化(±s)
組別 樣本數 HBV DNA(×106)24小時 48小時 72小時D 組 3 0.95±0.07 1.10±0.58 1.04±0.16 B 組 3 1.99±1.18 1.80±0.12 1.90±0.25 E 組 3 3.45±0.50 2.82±1.17 4.09±1.99
2.3.2 各組間肝臟酶學的變化(圖2):在48和72小時B組和D組肝細胞丙氨酸轉氨酶(ALT)水平有下降趨勢,與E組相比差異有統計學意義。天冬氨酸轉氨酶(AST)水平雖然有上升趨勢,但差異沒有統計學意義。說明免疫細胞與異種肝細胞之間混合培養,并不能導致細胞間的排斥反應發生,可能有保護肝細胞膜的作用。
2.4 特異性免疫效應淋巴細胞對HepG 2.2.15細胞內HBV DNA和cccDNA的影響(表3):成熟DC與LC作用(C組)24、48和72小時后,HepG 2.2.15細胞內的HBV DNA和cccDNA表達均明顯下降,與E組相比差異有統計學意義(均P<0.05)。未成熟DC與LC作用(A組)24、48和72小時后的HepG 2.2.15細胞內的HBV DNA和cccDNA表達亦均有下降,但與E組相比差異無統計學意義(P>0.05)。結果提示由HBV相關蛋白刺激的轉HBV基因鼠的DC刺激自體淋巴細胞后對體外HepG 2.2.15細胞中的HBV有顯著的抑制作用,對細胞內HBV cccDNA的復制及HBV DNA的分泌均有抑制作用。而未成熟DC組抑制HBV復制作用較成熟DC組作用弱。B組與D組在24、48和72小時對HBV DNA和cccDNA均有抑制作用,D組的作用更明顯,與E組比較有統計學意義(P<0.05)。而B組與E組比較無統計學意義(P>0.05)。
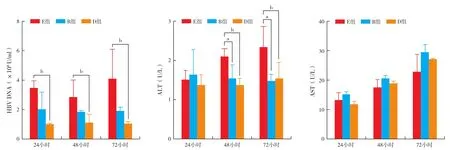
圖2 各組對HepG 2.2.15細胞上清的肝功能與HBV DNA的影響
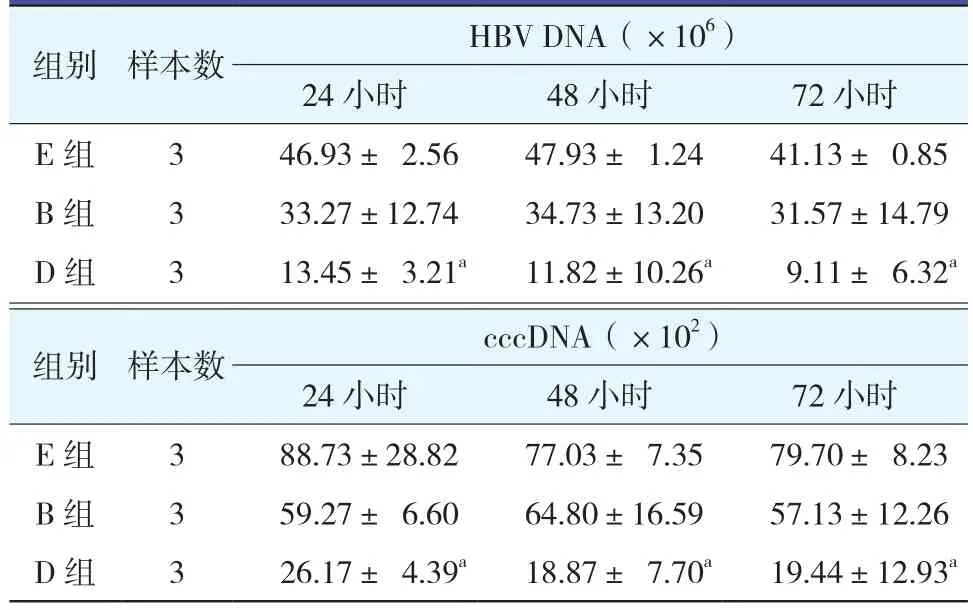
表3 各組不同時間點對HepG 2.2.15細胞內HBV DNA和cccDNA的影響
2.5 各組淋巴細胞亞群的變化(圖3):將成熟DC或不成熟DC與淋巴細胞共刺激后再與HepG 2.2.15細胞共培養,觀察CD3+CD4+、CD3+CD8+及CD3+CD4+/ CD3+CD8+比值的變化,結果提示,隨著DC由不成熟到成熟,CD3+CD4+及CD4+/ CD8+比值先下降后升高;而CD3+CD8+比例變化不明顯。說明成熟DC刺激的淋巴細胞在24小時以CD3+CD8+作用為主發揮免疫作用,48小時以后則以CD3+CD4+陽性細胞作用為主。
對CD3+CD4+細胞的影響:B、C、D組在24、48和72小時分別與A組相比差異均有統計學意義(均P<0.01),A組與B組、C組與D組相比差異也均有統計學意義(均P<0.01),表現為24小時先下降,48小時以后升高,而CD3+CD8+比例各個時間點均無明顯變化。
CD4+/ CD8+比值:C組較A組CD4+/ CD8+比值明顯降低;D組較C組CD4+/ CD8+比值在24小時明顯降低,48小時和72小時上升,差異有統計學意義(P<0.01);D組較B組CD4+/ CD8+比值在24小時明顯下降(P<0.01),48 小時和72小時明顯升高,48小時差異有統計學意義(P<0.05),而72小時差異無統計學意義(P>0.05)。
2.6 IEC與HepG 2.2.15細胞共培養后上清液的細胞因子水平變化(圖4;表4):A組與B組、C組與D組細胞上清液的IL-2和IFN-γ分泌水平在24、48和72小時均為上升趨勢,且差異均有統計學意義(均P<0.05);各個時間點D組與B組水平均下降,差異均有統計學意義(均P<0.05),且不隨時間而變化;C組與A組比較24小時時IFN-γ分泌量無明顯差異,而IL-2分泌量差異有統計學意義,其他時間點均有意義,但不隨時間變化而變化;C組與A組比較24和72小時IFN-γ無明顯差異。
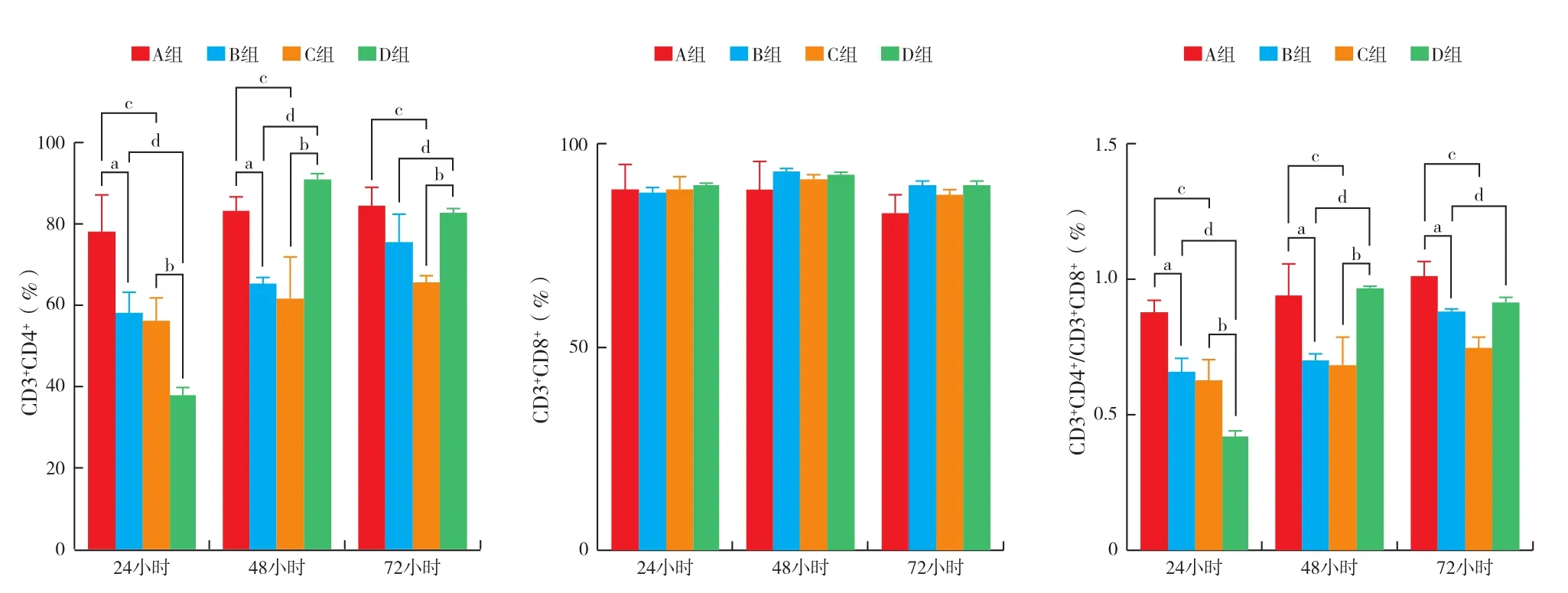
圖3 各組C57BL/6小鼠淋巴細胞亞群的變化
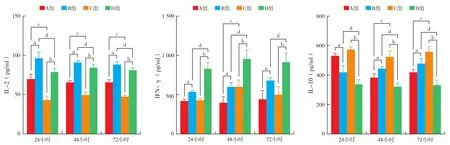
圖4 各組細胞上清液的IL-2、IFN-γ和IL-10分泌量的變化
IL-10的變化卻相反。A組與B組24小時呈下降趨勢,48小時和72小時時均呈上升趨勢(均P<0.05);C組與D組24、48和72小時均呈下降趨勢(均P<0.05);C組與A組24小時IL-10分泌量無明顯差異,D組與B組則有顯著差異(P<0.01)。但不隨時間變化而變化。
提示DC刺激淋巴細胞對體外HepG 2.2.15細胞作用后都能產生較高的IL-2、IFN-γ和較低的IL-10。但成熟DC較未成熟DC的作用更明顯。
表4 各組不同時間點、HepG 2.2.15細胞上清液細胞因子水平變化比較(±s)
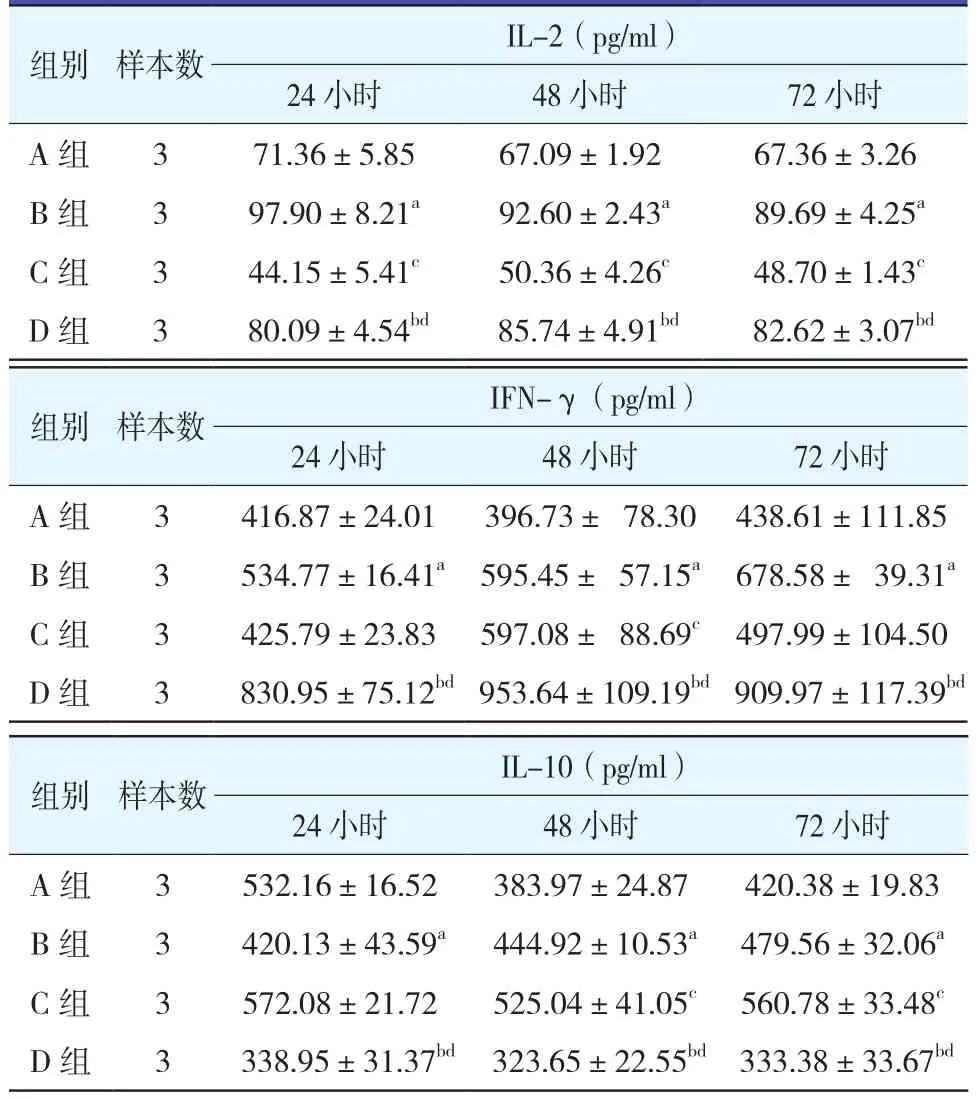
表4 各組不同時間點、HepG 2.2.15細胞上清液細胞因子水平變化比較(±s)
注:B組與A組比較,aP<0.05;D組與C組比較,bP<0.05;C組與A組比較,cP<0.05;D組與B組比較,dP<0.05
組別 樣本數 IL-2(pg/ml)24小時 48小時 72小時A 組 3 71.36±5.85 67.09±1.92 67.36±3.26 B 組 3 97.90±8.21a 92.60±2.43a 89.69±4.25a C 組 3 44.15±5.41c 50.36±4.26c 48.70±1.43c D組 3 80.09±4.54bd 85.74±4.91bd 82.62±3.07bd組別 樣本數 IFN-γ(pg/ml)24小時 48小時 72小時A 組 3 416.87±24.01 396.73± 78.30 438.61±111.85 B 組 3 534.77±16.41a 595.45± 57.15a 678.58± 39.31a C 組 3 425.79±23.83 597.08± 88.69c 497.99±104.50 D 組 3 830.95±75.12bd 953.64±109.19bd 909.97±117.39bd組別 樣本數 IL-10(pg/ml)24小時 48小時 72小時A 組 3 532.16±16.52 383.97±24.87 420.38±19.83 B 組 3 420.13±43.59a 444.92±10.53a 479.56±32.06a C 組 3 572.08±21.72 525.04±41.05c 560.78±33.48c D 組 3 338.95±31.37bd 323.65±22.55bd 333.38±33.67bd
3 討 論
多數學者認為,發生慢性病毒性肝炎及病毒持續感染的機制除了取決于病毒本身的生物學特性外,更重要的是與宿主自身的免疫功能狀態有關。利用自身的免疫細胞培養識別標靶特異抗原的淋巴細胞,以特定方法促使功能性淋巴細胞增殖,制成高純度、特異識別以及有殺傷標靶細胞(例如腫瘤、病毒感染的細胞等)功能的IEC。IEC的特點在于除了含有多種免疫活化所需的功能性細胞外,還可以促使免疫反應更加完整、活躍,達到高效清除病原的目的。有研究表明,提取患者自身DC,在體外經過HBcAg刺激后回輸到體內能夠促進T細胞免疫,從而抑制病毒復制和改善肝功能[12]。而Weiwei等[13]發現,通過HBcAg活化的DC能夠產生針對于HBV特異性的CD8+細胞反應。
本研究提取轉HBV基因C57BL/6小鼠DC及淋巴細胞,通過不同的HBV蛋白刺激DC成熟,并通過成熟的DC與淋巴細胞共培養得到IEC,在體外HepG 2.2.15細胞(含有完整的HBV基因)共培養,結果顯示了細胞間并沒產生異種細胞間的排斥反應,肝細胞的ALT有變化,但與對照組比較差異沒有統計學意義,而ALT有下降趨勢,故免疫細胞療法不會造成肝細胞的嚴重損傷[14],可能有修復肝細胞膜的作用。提示我們采用的體外處理方法對HBV的研究是可行的。其次我們發現,HBV相關抗原刺激的成熟DC與淋巴細胞共刺激組較未成熟DC與淋巴細胞組HepG 2.2.15細胞上清HBV DNA水平明顯下降,同時細胞內的HBV DNA和cccDNA水平也明顯下降。說明該種成熟DC作用后的淋巴細胞具有明顯的抑制HBV cccDNA的合成以及分泌的作用。
HBV cccDNA在HBV復制過程中起關鍵作用,是HBV復制的突出標志,很難被清除,監測其水平被認為是目前評價抗HBV療效或臨床治愈的“金標準”。目前認為,機體通過免疫清除HBV cccDNA的途徑主要為:① 通過細胞溶解機制,清除感染的肝細胞,HBV被巨噬細胞吞噬或與抗-HBs抗體結合成為免疫復合物后被巨噬細胞吞噬并從尿中排出,破壞的肝細胞由新生肝細胞替代;② 通過Th1反應介導的依賴細胞因子的非細胞溶解機制,如IFN-γ、TNF-α和IL-2等細胞因子介導非細胞裂解機制清除細胞核內的HBV cccDNA或阻斷合成新的HBV cccDNA,即通過細胞因子的作用“治療”感染[15-16],從而清除HBV。目前在移植患者中發現可以通過供者針對HBV的過繼免疫而實現HBV的清除[17]。
為了解釋其原因及機制,我們檢測了致敏淋巴細胞的亞群變化,發現特異性IEC在作用HBV的早期主要以CD8+細胞為主,CD4+細胞水平明顯降低,CD4+/ CD8+比值顯著降低,48小時以后CD4+細胞明顯升高,表明IEC的抗HBV的早期作用主要是CD8+細胞發揮的,隨后以CD4+T細胞作用為主;隨著時間的延長,CD4+/CD8+比值上升,而細胞的HBV DNA水平明顯下降,可能與在抗HBV過程中CD4+和CD8+細胞直接參與有關[18-19]。HBV DNA水平降低,CD4+/ CD8+比值卻明顯升高,說明IEC的亞群改變首先是CD4+/ CD8+比值的降低,而后在HBV作用后升高,并且與異種細胞間的排異反應無關。有研究表明,IEC的CD8+淋巴細胞是決定HBV清除的關鍵細胞亞群,細胞溶解或非溶解機制(IFN-γ和TNF-α等)是CD8+T細胞清除HBV的主要方式,而CD4+淋巴細胞主要通過分泌Thl細胞因子來抑制HBV復制[20-21],提示此種情況下加強急性CD4+T細胞應答的治療和疫苗的策略是有意義的。而DC是目前已知的唯一能激活Th0細胞向Thl細胞或Th2細胞分化的功能最強的抗原呈遞細胞[22]。為了進一步解釋其機制,我們對各組進行了細胞因子的檢測。
Th1和Th2兩個細胞亞群相互拮抗,Thl/Th2動態平衡是機體處于正常免疫狀態的保證[23]。Thl主要分泌IFN-γ、IL-2、IL-3等細胞因子,這些細胞因子誘發細胞免疫反應,增強微生物對宿主的感染,尤其是病毒與細胞內感染的免疫性和防御功能,達到清除病毒的目的;Th2主要分泌IL-10、IL-4、IL-5等細胞因子,這些細胞因子可刺激B細胞增生,產生相應抗體,誘發體液免疫反應[24],并與感染的進展、慢性化有關。Thl/Th2的失衡應答可能是HBV感染慢性化的主要機制,這種失衡又體現在細胞因子失調方面[25]。IFN-γ、IL-2能正向調節免疫應答,清除侵入體內的病毒抗原。IL-10是一種重要的抗炎細胞因子,也可抑制Th1成熟和產生相應細胞因子,故IFN-γ、IL-2、IL-10可大致代表 Th1 / Th2的功能[26-27]。我們檢測了IFN-γ、IL-2和IL-10,結果顯示,HBV DNA高表達時,IFN-γ和IL-2水平下降,而IL-10均升高,HBV DNA低表達時,IFN-γ和IL-2水平上升,而IL-10水平下降。由于IFN-γ分泌增加可使CD4+T細胞向Th1分化,抑制Th2形成,從而激活HBV特異的CTL克隆增殖,殺傷HBV[28]。IL-10含量升高,且伴隨病毒載量增高而增加,顯示HBV致病與免疫應答中Th2介導的體液免疫活性偏高有關系。IL-10水平升高抑制了Th1群細胞的功能,使Th1分泌IFN-γ和IL-2的量減少,細胞免疫效應降低,機體清除病毒能力減弱,導致病毒持續存在[29]。有研究報道HBV感染者血清IL-10濃度明顯高于正常對照組,HBV高載量組感染者血清IL-10濃度明顯高于正常對照組和低載量組[30],說明細胞因子與肝細胞中HBV DNA的消長有一定關系,特別是與肝細胞中HBV cccDNA、HBV DNA的關系需要進一步研究,以探討cccDNA的清除與機體免疫的關系。
以往人們對血清及腹腔積液中細胞因子與HBV DNA的研究較多,但對細胞因子與肝細胞中HBV DNA及HBV cccDNA的關系卻研究較少。本研究結果證實,IEC對體外HBV DNA及HBV cccDNA均有明顯的抑制作用,這一結果也與其他報道[31-33]一致。通過對轉HBV基因小鼠的DC處理的淋巴細胞亞群檢測,可以證明體外IEC中HBV特異性的CD4+和CD8+細胞對HBV清除和疾病轉歸有重要影響,對體外HBV復制及分泌均有抑制作用。肝移植術后HBV再感染的病原來源于病肝釋放到循環中的病毒顆粒,或者淋巴系統及淋巴細胞等肝外組織中存在的病毒[34]。而移植術后,隨著大量免疫抑制劑的應用,肝移植受體術后外周血來源的DC呈遞乙肝表面抗原的能力低下,針對HBV的抗原特異性T細胞應答低下,此種功能缺陷是導致受體針對HBV的主動免疫反應性低的重要原因[35]。特異修飾的DC刺激淋巴細胞可以在很大程度上減少HBV的復制,不會造成肝功能的損傷。這種療法可能在不久的將來用于預防和治療移植術后乙肝復發患者。
綜上所述得出以下結論:① IEC與異種肝細胞體外混合培養,細胞間不產生排斥反應;② 轉HBV基因鼠DC誘導的IEC對體外HBV復制有明顯的抑制作用,為免疫細胞療法在臨床中的應用提供理論依據;③ Th1/Th2平衡可能在控制體外HBV復制及分泌過程中處于一個動態的平衡;④ 細胞因子在抗HBV中發揮重要作用,可能也是保護肝細胞損傷的重要因素。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
海峽科技與產業(2016年3期)2016-05-17 04:32:12
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
