軸突導向蛋白4D在腫瘤進展中的作用*
2015-11-23 02:34:02張磊陳穎王珂
中國腫瘤臨床 2015年3期
張磊 陳穎 王珂
·國家基金研究進展綜述·
軸突導向蛋白4D在腫瘤進展中的作用*
張磊 陳穎 王珂
軸突導向蛋白4D(SEMAPHORIN4D,SEMA4D)又稱CD100,是軸突導向蛋白Ⅳ亞族(SEMAsⅣ)的重要成員之一,其最初作為影響神經發育的軸突導向分子被發現。近來越來越多的研究表明,SEMA4D在免疫調節、血管發生及腫瘤生長轉移等方面也有重要的生物學功能,尤其作為新的促血管生成分子,SEMA4D的相關研究引起廣泛關注。本文旨在對SEMA4D的結構、受體特點、促血管生成及腫瘤侵襲轉移等方面的最新研究進展進行綜述。
SEMA4D 軸突導向 腫瘤 免疫調節 血管生成
Department of Gynecologic Oncology,Tianjin Medical University Cancer Institute and Hospital,National Clinical Research
Center for Cancer,Key Laboratory of Cancer Prevention and Therapy of Tianjin,Tianjin 300060,China
This work was supported by the National Natural Science Foundation of China(No.81302250)and Tianjin Health Bureau of Science and Technology Funds(No.2012KZ073)
在神經發育過程中,軸突導向分子引導軸突選擇正確生長方向,從而成功到達靶區,近年來關于軸突導向分子的研究進展迅速,SEMAPHORINs(SEMAs)家族是其中越來越受到關注的分子之一,該家族蛋白主要有2種受體:Plexins和Neuropilins[1]。但是,SEMAs家族除了最初作為軸突導向分子影響中樞神經系統的發育而被發現外,隨后越來越多的研究表明在多種細胞中都有SEMAs及其受體表達,從而引起一系列關于其功能及信號轉導通路的研究[2]。SEMAPHORIN4D(SEMA4D)又稱CD100,是SEMAPHORINs家族的重要成員之一,許多研究發現其在神經系統、免疫調節、血管發生、腫瘤侵襲轉移等方面均起著重要作用[3]。
1 SEMAPHORINs概述
SEMAs家族是一大類結構高度同源的分泌型或跨膜型蛋白分子,其蛋白結構中均包含一個SEMA結構域,其在神經系統發育過程中發揮重要作用。SEMAs分子可以調節神經軸突導向及介導細胞紡錘體的形成[4]。20多個SEMAs蛋白根據分子序列的相似度及結構的不同特點將其分為8個亞族,SEMAⅠ和SEMAⅡ亞族存在于無脊椎動物種屬中,SEMAⅢ到SEMAⅦ亞族表達于脊椎動物種屬,SEMAⅧ亞族則來自于非神經來源的病毒中[5](圖1)。既往研究表明在神經系統發育中,SEMAs蛋白主要參與軸突生長調控,軸突分裂、收縮,神經極化以及神經細胞遷移等生理過程,但近來越來越多的研究證實SEMAs分子在神經元細胞凋亡、轉化,免疫反應,細胞遷移,腫瘤發生發展等過程中均扮演重要角色[7-8]。
2 SEMA4D及其受體
2.1結構
SEMA4D是一個分子量為150 kD的跨膜蛋白分子,其結構包括一個氨基末端,一個SEMA結構域
(SEMA domain),一個免疫球蛋白域(Ig domain),一個富有賴氨酸的延長區,一個疏水的跨膜區和一個存在于細胞漿中的尾部結構。SEMA4D以同源二聚體形式分布在細胞表面,其細胞外的結構部分包括多個N-連接糖基化位點和17個高度保守的半胱氨酸殘基,用于亞基內二硫鍵的形成;胞內區有一個酪氨酸及多個絲氨酸磷酸化位點[9](圖2)[10]。經過蛋白酶水解作用,SEMA4D能夠以分泌型蛋白形式從細胞膜上釋放到局部微環境中。
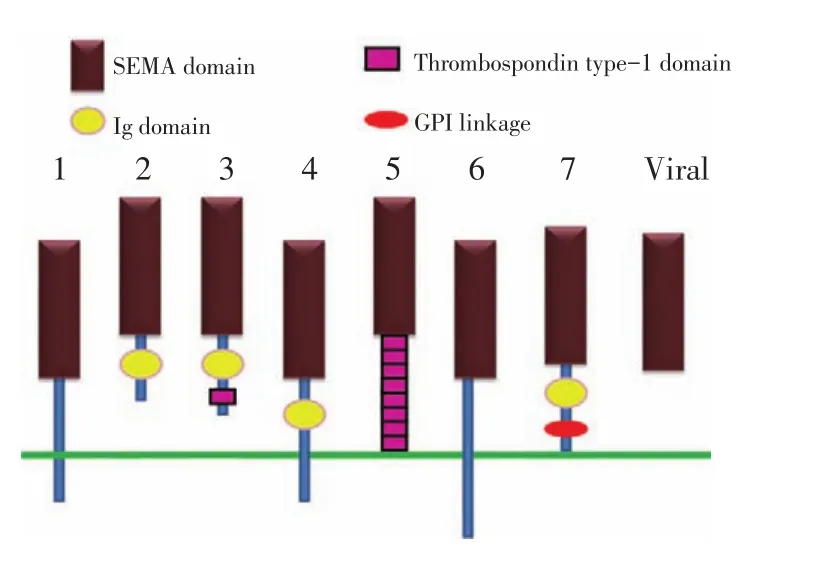
圖1 SEMAPHORINs(SEMAs)蛋白家族結構示意圖Figure 1Semaphorin(SEMA)family contains a large number of phylogenetically conserved,secreted,and transmembrane proteins.On the basis of structural features,the members were divided into eight classes,including a unique viral class.The members of SEMA family all share a common SEMA domain
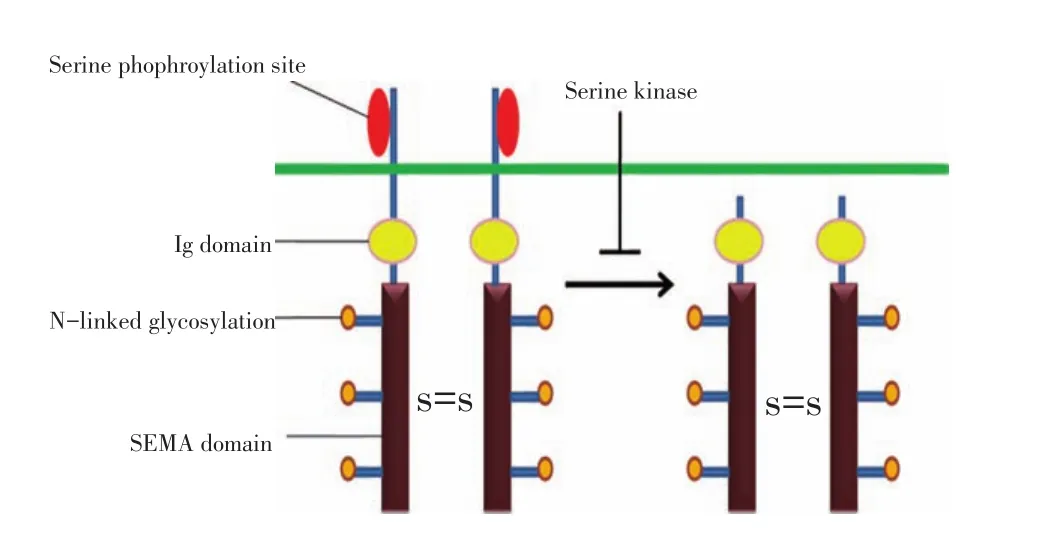
圖2 SEMPHORIN4D(SEMA4D)分子結構Figure 2The structure of semaphorin 4D(SEMA4D)consists of an amino-terminal signal sequence,a SEMA domain,Ig-like domain,a lysine-rich stretch,a transmembrane region,and a cytoplasmic region.Although SEMA4D is a transmembrane-type semaphorin,it is proteolytically cleaved into a soluble form
2.2分布
SEMA4D分布廣泛,在胚胎及成人組織中均有較高表達,其中包括非淋巴組織(心、腦、腎)和淋巴組織(脾、胸腺和淋巴結)[11]。在淋巴器官中,SEMA4D主要表達于靜息的T細胞表面,而在靜息的B細胞及抗原呈遞細胞(APC),如樹突細胞(DC)上則表達較低。但當B細胞或DC細胞被活化后,SEMA4D的表達明顯上調。在其他與宿主防御能力有關的細胞中,如單核巨噬細胞、中性粒細胞、內皮細胞和上皮細胞上也有SEMA4D的分布[12]。并且最近的研究表明多種實體腫瘤細胞上SEMA4D也有較高的表達,包括頭頸鱗狀細胞癌、乳腺癌、前列腺癌、結腸癌、軟組織肉瘤和卵巢癌[13-17]。
2.3受體
SEMA4D的受體主要包括:CD72和PlexinB1。CD72是一個分子量為45 kD的Ⅱ型跨膜蛋白,其屬于C型凝集素超家族,是SEMA4D在淋巴組織的主要受體。CD72主要表達于免疫細胞,如B細胞、APC及肥大細胞等。對于B細胞來說,CD72表達于從祖B細胞到成熟B細胞各階段,而分化到漿細胞階段CD72則不表達。CD72胞漿區含有2個免疫受體酪氨酸抑制序列(immunoreceptor tyrosine-based inhibitory mofifs,ITIMs),在B細胞發育分化及免疫應答中發揮負性調節作用。CD72與BCR發生交聯誘導ITIM酪氨酸磷酸化,并與蛋白質酪氨酸磷酸酶(SHP-1)結合,誘導酪氨酸去磷酸化及下游信號蛋白滅活,繼而促使靜止的B細胞向祖細胞發生分化,并且可以降低活化的B細胞增殖。而SEMA4D和競爭性CD72單克隆抗體(CD72-mAb)與CD72結合,促進CD72從B細胞受體復合物上解離,ITIM酪氨酸去磷酸化及CD72與SHP-1解離,從而消除CD72的抑制作用,使B細胞活化,并提高B細胞的聚集、生存能力及APC的活化與成熟[18]。
PlexinB1是跨膜蛋白Plexins蛋白家族的重要成員之一,是SEMA4D的高親和力受體,最初是從小鼠腎臟來源的細胞上發現的。PlexinB1細胞外部分的分子結構與Met分子有28%的相似度,包括一個SEMA結構域,3個PSI結構域,3個甘氨酸-脯氨酸(glycine-proline,G-P)富集模體,而胞漿部分含有2個GAP(GTPase活化蛋白)樣結構域,中間1個GTPase結合結構域,胞質尾部有一個盤狀同源區域(PSD-95,Discs large,zona occludens-1,PDZ)結構域結合位點。PlexinB1以異源二聚體的形式分布于內皮細胞胞膜上,包括α和β兩個亞基。β跨膜亞基包括較短的細胞外結構域和細胞漿結構域,而α亞基主要包括較長的細胞外結構域,α亞基和β亞基之間通過較為疏松的相互作用結合于細胞膜表面。經過蛋白水解作用,SEMA4D、PlexinB1和Met分子三者之間形成一個功能性的復合體存在于內皮細胞的表面,繼而發揮作用。相似地,Met分子在胞膜外區也存在1個SEMA結構域,1個PSI結構域,4個IgSF樣結構域。胞質區有1個酪氨酸激酶結構,當SEMA4D、PlexinB1及Met形成復合物后,酪氨酸激酶使Met和PlexinB1胞質區以及某些下游信號分子發生磷酸化,
從而參與血管形成、細胞遷移和侵襲等生物學過程。另外,Swiercz等[19]報道PlexinB1和酪氨酸激酶受體ErbB-2關系密切,PlexinB1與SEMA4D結合后誘導ErbB-2發揮內源性活性。在活化的T細胞上也發現了PlexinB1轉錄子,但其詳細的作用機制還有待深入研究。
3 SEMA4D促進血管生成
血管生成是惡性腫瘤侵襲轉移的重要環節,SEMA4D能夠通過介導內皮細胞遷移、管腔形成從而促進腫瘤血管發生。研究表明SEMA4D的促進腫瘤血管生成作用與其他一些重要的促血管發生的分子相類似,如血管內皮生長因子(vascular epithelial growth factor,VEGF)、HGF、bFGF等[20-21]。
SEMA4D的促血管生成作用是與其受體PlexinB1結合后啟動(繪制模式圖3)。PlexinB1廣泛高表達于內皮細胞胞膜表面,細胞膜上的SEMA4D分子通過水解酶作用之后釋放到微環境中,與內皮細胞膜上PlexinB1結合后促使一系列級聯式反應發生,從而介導血管生成,但其具體過程仍然存在爭議。Basile等[22]認為SEMA4D與其受體PlexinB1羧基端的PDZ結合域結合后,繼續結合RhoGEF和LARG分子,從而激活RhoA分子及整合素分子(Intergrins),繼而促進腫瘤血管發生,這一過程與細胞內酪氨酸激酶途徑的活化密切相關,尤其是Pyk2分子的參與在其中發揮了重要作用。但是,Conrotto等[20]卻認為SEM4D的促血管生成作用則是通過Met基因激活,繼而酪氨酸磷酸化的過程實現的。雖然上述兩種關于機制分析的觀點存在分歧,但兩人仍然一致認為最終都是導致內皮細胞骨架發生變化及細胞遷移而實現了腫瘤新生血管的發生。
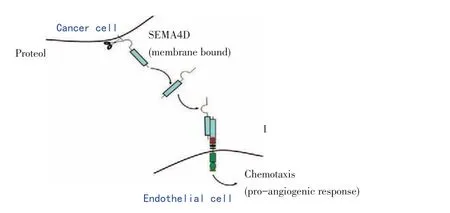
圖3 SEMA4D與受體PlexinB1結合促進血管生成模式圖Figure 3Model for semaphorin 4D(SEMA4D)/Plexin-B1 promoted angiogenesis.Membrane-bound SEMA4D was released by proteolytic cleavage.Subsequently,soluble SEMA4D bound Plexin-B1 on endothelial cells of the tumor stroma to initiate an angiogenic response
Basile等[23]在頭頸鱗癌的研究中發現:頭頸鱗癌細胞高表達SEMA4D,SEMA4D以分泌型蛋白的形式從癌細胞中釋放出來促進內皮細胞的遷移,從而促進腫瘤新生血管生成介導癌細胞發生遠處轉移,并且體內實驗證實在缺乏SEMA4D的腫瘤微環境中,癌細胞的生長或轉移明顯受到抑制。Sierra等[24]在裸鼠的乳腺癌成瘤模型中也觀察到了相似的結果:沉默腫瘤微環境中SEMA4D的表達后,腫瘤的微血管形成能力和轉移能力明顯降低。但與前者研究矛盾的是,Chen等[17]認為腫瘤微環境中分泌型的SEMA4D蛋白來自于腫瘤相關巨噬細胞(tumor-associated macrophage,TAM)而不是由腫瘤細胞分泌的,這種不一致結果的原因可能是由不同的腫瘤類型造成的。本研究組也證實:上皮性卵巢癌中SEMA4D高表達,與患者的臨床病理特征及預后密切相關,SEMA4D是上皮性卵巢癌患者無疾病進展生存期及總生存期的獨立預后因素。
4 SEMA4D調控侵襲轉移
侵襲生長是一個多步驟的復雜的形態發生過程,主要包括腫瘤細胞分離、細胞遷移、細胞種植、轉移灶形成、細胞增殖和細胞存活6個步驟。侵襲生長可見于正常的生理過程中,如胚胎發育和傷口愈合等。然而,侵襲生長是腫瘤進展過程中的重要環節[25]。研究表明,SEMA4D的重要受體之一PlexinB1在多種實體瘤中高表達,如乳腺癌、結腸癌、肝癌、胰腺癌和胃癌等。SEMA4D與PlexinB1結合后通過Met信號途徑促進了上述惡性腫瘤細胞的侵襲轉移[26]。但是,Swiercz等[19]研究表明SEMA4D與PlexinB1結合后是否促進癌細胞的侵襲生長取決于癌組織中PlexinB1、Met及ErbB-2的表達量的高低。ErbB-2是一個比較公認的在乳腺癌上提示患者預后較差的指標,SEMA4D與ErbB-2結合促進惡性腫瘤細胞侵襲生長,但與PlexinB1或Met結合后則抑制癌細胞的遷移。惡性腫瘤中,SEMA4D是促進癌細胞遷移還是抑制癌細胞遷移的效應取決于SEMA4D與PlexinB1、Met還是ErbB-2結合,復雜的下游的分子信號通路依賴于Rho分子的活化情況,但詳細的分子機制還需要進一步的研究[27]。
另外,研究表明SEMA4D與PlexinB1結合后促進神經軸突的生長。報道提到PlexinB1本身具有與R-Ras蛋白相類似的內在GAP活性。R-Ras失活抑制了細胞間粘附、細胞遷移及生長錐的外生長。與上述激活途徑相反,這種抑制作用降低了細胞的遷移。而PlexinB1的R-Ras GAP突變可以減少細胞粘附、遷移和侵襲的發生[28]。
5 SEMA4D在免疫系統的作用
外源性SEMA4D或可溶性重組SEMA4D蛋白均可促進B細胞的活化。體內實驗通過分析CD40、
CD80及IL-12的表達,發現SEMA4D促進CD40介導的抗原呈遞細胞(DC)細胞的成熟。并且在敲除SEMA4D表達的免疫缺陷小鼠的動物模型研究中觀察到:抑制SEMA4D的表達后,小鼠體內抗體的產生及T細胞誘發的特異性抗體的表達均降低。此外,無論是重組可溶性SEMA4D蛋白還是競爭性的SEMA4D單克隆抗體均可介導單核細胞釋放IL-6、IL-8等細胞因子促進炎癥發生[29]。Delaire等[30]報道無論對于新鮮分離的單核細胞還是單核細胞株,SEMA4D均可抑制自發性或MCP-3介導的細胞遷移。但SEMA4D的受體PlexinB1或CD72是否參與SEMA4D介導的免疫細胞的遷移尚不清楚。
在腫瘤相關巨噬細胞中,雖然缺乏SEMA4D的表達不能改變巨噬細胞本身的分化、激活以及細胞因子的產生,但是敲除腫瘤微環境中SEMA4D的表達后明顯可以下調腫瘤相關巨噬細胞的數量[24]。
6 SEMA4D與腫瘤患者預后及治療
SEMA4D在多種惡性實體瘤中表達明顯增高并與患者預后密切相關。在宮頸癌中,SEMA4D高表達提示患者總生存時間明顯縮短,這可能與SEMA4D通過自分泌的方式介導腫瘤血管發生從而促進癌細胞發生遠處轉移有關[31]。此外,在卵巢癌[17]及骨肉瘤[15]中的研究均發現,SEMA4D表達增高預示患者無疾病進展生存期及總生存期均較對照組縮短。SEMA4D的重要作用之一即誘導腫瘤新生血管形成。而新生血管形成是惡性腫瘤侵襲轉移的關鍵環節,SEMA4D作為重要的促血管生成因子,可以誘導內皮細胞的遷移和管腔形成,因此其高親和力受體PlexinB1可以成為抗腫瘤血管生成藥物的重要靶標[32]。丁曉潔等[33]已經證實:在裸鼠直腸癌移植瘤模型中,抑制SEMA4D的表達可以減弱癌細胞誘導內皮細胞遷移的能力,顯著減緩移植瘤生長速度并降低瘤體內血管密度,這也為SEMA4D的臨床抗血管生成治療進一步奠定了基礎。另外,SEMA4D的新型人源化單克隆抗體(mAb)VX15/2503顯示出良好的腫瘤治療應用前景。在2010年第6屆歐洲抗體大會上,Vaccinex公司報告VX15/2503可有效阻斷SEMA4D和plexinB1的相互作用,并且在小鼠CT26和EM6腫瘤模型中,已經觀察到VX15/2503可有效抑制腫瘤的生長和血管形成。2011年Vaccinex公司被FDA批準進行治療性VX15/2503的Ⅰ期臨床試驗,目的在于評估VX15/ 2503的安全性、耐受性、藥代動力學及藥效學。
7 展望
總的來說,SEMA4D在免疫、神經發育,促進腫瘤進展,尤其是腫瘤血管生成方面均扮演重要角色。在多種實體瘤組織中,SEMA4D高表達提示腫瘤細胞惡性程度較高,并且SEMA4D與受體PlexinB1結合后可以促進TAM遷移及增強TAM促進腫瘤微血管生成效應,但其作用的具體詳細機制仍需進一步探索。隨著SEMA4D研究的更加深入,SEMA4D作為靶點應用于惡性腫瘤的臨床診斷和治療必將有廣闊的前景,并且當抗VEGF治療效果不佳時,聯合或補充給予針對SEMA4D信號途徑多靶點抗血管生成治療也許能產生令人振奮的效果,而其在轉化醫學的研究和應用中也將扮演越來越重要的角色。
1Takegahara N,Kumanogoh A,Kikutani H.Semaphorins:a new class of immunoregulatory molecules[J].Philos Trans R Soc Lond B Biol Sci,2005,360(1461):1673-1680.
2Gu C,Giraudo E.The role of semaphorins and their receptors in vascular development and cancer[J].Exp Cell Res,2013,319(9): 1306-1316.
3Eissler N,Rolny C.The role of immune semaphorins in cancer progression[J].Exp Cell Res,2013,319(11):1635-1643.
4Gherardi E,Love CA,Esnouf RM,et al.The sema domain[J]. Curr Opin Struct Biol,2004,14(6):669-678.
5Koropouli E,Kolodkin AL.Semaphorins and the dynamic regulation of synapse assembly,refinement,and function[J].Curr Opin Neurobiol,2014,27:1-7.
6Kumanogoh A,Kikutani H.Biological functions and signaling of a transmembrane semaphorin,CD100/Sema4D[J].Cell Mol Life Sci,2004,61(3):292-300.
7Rehman M,Tamagnone L.Semaphorins in cancer:biological mechanisms and therapeutic approaches[J].Semin Cell Dev Biol,2013,24(3):179-189.
8Micucci C,Orciari S,Catalano A.Semaphorins and their receptors in stem and cancer cells[J].Curr Med Chem,2010,17(30):3462-3475.
9Chao KL,Tsai IW,Chen C,et al.Crystal structure of the Sema-PSI extracellular domain of human RON receptor tyrosine kinase[J].PLoS One,2012,7(7):e41912.
10 Suzuki K,Kumanogoh A,Kikutani H.CD100/Sema4D,a lymphocyte semaphorin involved in the regulation of humoral and cellular immune responses[J].Cytokine Growth Factor Rev,2003,14(1): 17-24.
11 Zhang Y,Liu B,Ma Y,et al.Sema 4D/CD100-plexin B is a multifunctional counter-receptor[J].Cell Mol Immunol,2013,10(2):97-98. 12 Nkyimbeng-Takwi E,Chapoval SP.Biology and function of neuroimmune semaphorins 4A and 4D[J].Immunol Res,2011,50(1): 10-21.
13 Ch'Ng ES,Kumanogoh A.Roles of Sema4D and Plexin-B1 in tumor progression[J].Mol Cancer,2010,9:251.
14 Campos M,DE Campos SG,Ribeiro GG,et al.Ki-67 and CD100 immunohistochemical expression is associated with local recurrence and poor prognosis in soft tissue sarcomas,respectively[J].Oncol Lett,2013,5(5):1527-1535.
15 Ch'Ng E,Tomita Y,Zhang B,et al.Prognostic significance of CD100 expression in soft tissue sarcoma[J].Cancer,2007,110(1): 164-172.
16 Chen Y,Zhang L,Lv R,et al.Overexpression of Semaphorin4D in-
dicates poor prognosis and prompts monocyte differentiation toward M2 macrophages in epithelial ovarian cancer[J].Asian Pac J Cancer Prev,2013,14(10):5883-5890.
17 Chen Y,Zhang L,Pan Y,et al.Over-expression of semaphorin4D,hypoxia-inducible factor-1alpha and vascular endothelial growth factor is related to poor prognosis in ovarian epithelial cancer[J].Int J Mol Sci,2012,13(10):13264-13274.
18 Wu HJ,Bondada S.CD72,a coreceptor with both positive and negative effects on B lymphocyte development and function[J].J Clin Immunol,2009,29(1):12-21.
19 Swiercz JM,Worzfeld T,Offermanns S.ErbB-2 and met reciprocally regulate cellular signaling via plexin-B1[J].J Biol Chem,2008,283(4):1893-1901.
20 Conrotto P,Valdembri D,Corso S,et al.Sema4D induces angiogenesis through Met recruitment by Plexin B1[J].Blood,2005,105(11): 4321-4329.
21 Zhou H,Yang YH,Binmadi NO,et al.The hypoxia-inducible factor-responsive proteins semaphorin 4D and vascular endothelial growth factor promote tumor growth and angiogenesis in oral squamous cell carcinoma[J].Exp Cell Res,2012,318(14):1685-1698.
22 Basile JR,Barac A,Zhu T,et al.Class IV semaphorins promote angiogenesis by stimulating Rho-initiated pathways through plexin-B[J].Cancer Res,2004,64(15):5212-5224.
23 Basile JR,Castilho RM,Williams VP,et al.Semaphorin 4D provides a link between axon guidance processes and tumor-induced angiogenesis[J].Proc Natl Acad Sci U S A,2006,103(24):9017-9022.
24 Sierra JR,Corso S,Caione L,et al.Tumor angiogenesis and progression are enhanced by Sema4D produced by tumor-associated macrophages[J].J Exp Med,2008,205(7):1673-1685.
25 Trusolino L,Comoglio PM.Scatter-factor and semaphorin receptors:cell signalling for invasive growth[J].Nat Rev Cancer,2002,2(4):289-300.
26 Soong J,Chen Y,Shustef EM,et al.Sema4D,the ligand for Plexin B1,suppresses c-Met activation and migration and promotes melanocyte survival and growth[J].J Invest Dermatol,2012,132(4): 1230-1238.
27 Swiercz JM,Kuner R,Offermanns S.Plexin-B1/RhoGEF-mediated RhoA activation involves the receptor tyrosine kinase ErbB-2[J].J Cell Biol,2004,165(6):869-880.
28 Wong OG,Nitkunan T,Oinuma I,et al.Plexin-B1 mutations in prostate cancer[J].Proc Natl Acad Sci U S A,2007,104(48): 19040-19045.
29 Suzuki K,Kumanogoh A,Kikutani H.Semaphorins and their receptors in immune cell interactions[J].Nat Immunol,2008,9(1):17-23.
30 Delaire S,Billard C,Tordjman R,et al.Biological activity of soluble CD100.II.Soluble CD100,similarly to H-SemaIII,inhibits immune cell migration[J].J Immunol,2001,166(7):4348-4354.
31 Liu H,Yang Y,Xiao J,et al.Semaphorin 4D expression is associated with a poor clinical outcome in cervical cancer patients[J].Microvasc Res,2014,93:1-8.
32 Malik MF,Ye L,Jiang WG.The Plexin-B family and its role in cancer progression[J].Histol Histopathol,2014,29(2):151-165.
33 Ding XJ,Li D,Huang XW,et al.Sema4D deficiency reduces colorectal carcinoma xenograft growth and vascularityin nude mice[J].Chinese Journal of Clinical Oncology,2014,41(14):885-889.[丁曉潔,李多,黃新偉,等.信號素4D影響直腸癌裸鼠移植瘤的血管新生[J].中國腫瘤臨床,2014,41(14):885-889.]
(2014-09-10收稿)
(2015-01-02修回)
(編輯:鄭莉)
Functions of semaphorin 4D in tumor progression
Lei ZHANG,Ying CHEN,Ke WANG
Ke WANG;E-mail:lychenying2004@126.com
Semaphorin 4D(SEMA4D),also known as CD100,is a protein that belongs to class IV semaphorin.Its physiologic role in the nervous system has been extensively explored.However,the roles of SEMA4D have extended beyond the traditionally studied territories.Collective data proved that SEMA4D has an important role in the regulation of immune system,angiogenesis,and tumorigenesis,among others.Specifically,SEMA4D enhanced the angiogenesis in malignant diseases.This review summarized the latest progression in the research on SEMA4D,including the structure,receptors,mechanism,and biological functions in tumor angiogenesis and progression.
SEMA4D,axon guidance,tumor,immunoregulation,angiogenesis

10.3969/j.issn.1000-8179.20141558
天津醫科大學腫瘤醫院婦瘤科,國家腫瘤臨床醫學研究中心,天津市腫瘤防治重點實驗室(天津市300060)
*本文課題受國家自然科學基金項目(編號:81302250)和天津市衛生局科技基金項目(編號:2012KZ073)資助
王珂lychenying2004@126.com
張磊專業方向為婦科腫瘤臨床診治。
E-mail:zhanglei008_008@126.com
