朱紅毛斑蛾生物學特性研究
2015-11-25 08:28:30劉俊延何秋隆鄭霞林
植物保護 2015年3期
劉俊延, 何秋隆, 魏 航, 楊 娟, 李 俊, 陸 溫, 鄭霞林
(廣西大學農學院, 南寧 530004)
?
朱紅毛斑蛾生物學特性研究
劉俊延, 何秋隆, 魏 航, 楊 娟, 李 俊, 陸 溫, 鄭霞林*
(廣西大學農學院, 南寧 530004)
朱紅毛斑蛾(PhaudaflammansWalker)(鱗翅目:斑蛾科)是榕樹上的一種重要食葉害蟲,通過室內觀察和野外調查,研究了朱紅毛斑蛾的形態特征、年生活史及生活習性。結果表明,朱紅毛斑蛾卵粒聚集排列成連續線狀;幼蟲共6齡;雌雄成蟲可通過觸角鞭節顏色、腹末有無毛須和腹部腹面體毛顏色加以區分。在廣西1年發生2~3代,以第2代預蛹期幼蟲及蛹和第3代低齡幼蟲越冬。成蟲羽化高峰為10:00-13:00;交配高峰期為13:00-18:00;產卵高峰期為11:00-15:00;幼蟲僅為害榕屬植物葉片,當食料缺乏時還會為害其枝條韌皮部;越冬幼蟲喜歡在位于地表突出的根系附近或雜草叢下化蛹越冬,極少數在土壤表層0~10 mm處做蛹室化蛹。自然條件下發現2種朱紅毛斑蛾的寄生性天敵。
朱紅毛斑蛾; 形態特征; 年生活史; 生活習性
榕樹是我國南方城市及東南亞諸國廣為栽培的主要綠化樹種之一。由于榕樹的大面積栽培,導致園林生態系統多樣性逐步降低,害蟲種類數量增加至80余種,涉及食葉、刺吸、蛀干及地下害蟲等[1-5],常造成葉片枯黃、缺刻、落葉、枯枝,甚至整株枯死,不僅失去了觀賞價值,還降低了其在生態環境中應發揮的吸塵、降溫、吸收CO2等功效,嚴重影響其景觀效果和生態效益。
朱紅毛斑蛾(PhaudaflammansWalker)(鱗翅目:斑蛾科)是榕樹上的一種重要食葉害蟲,近幾年其發生和為害呈逐步加重趨勢。2011年10月南寧電視臺報道了南寧市仙葫大道兩旁大量垂葉榕變成“禿子”的新聞,就是朱紅毛斑蛾為害的結果。目前,由于缺乏對其生物學特性的了解,阻礙了朱紅毛斑蛾種群管理工作的進程。鑒于此,筆者通過室內飼養和觀察并結合野外調查,對朱紅毛斑蛾形態特征、年生活史和生活習性進行了初步研究,以期為其種群管理提供參考。
1 材料與方法
1.1 供試蟲源
于2012年11月在廣西大學校園內(108.29°E,22.84°N)采集朱紅毛斑蛾幼蟲置于培養皿中(d=9 cm),在室溫下,采集新鮮的黃金榕(Ficusmicrocarpa‘Golden Leaves’)葉片單頭飼養,每天更換1次食料,飼養至成蟲作為供試蟲源。
1.2 形態特征的觀察
將30對成蟲置于放有盆栽黃金榕的網籠(長×寬×高=50 cm×50 cm×80 cm)中讓其自由交配和產卵。并將10%蜂蜜水浸泡的棉團置于培養皿內,供成蟲補充營養。成蟲在盆栽植株上產卵后,收集卵粒(n=357),置于體視鏡(GL-6545TI,桂林桂光儀器有限公司)下觀察其形態特征,利用體視鏡上的數碼相機(YM130,山東易創電子有限公司)進行拍照。幼蟲孵化后,轉移至培養皿中單頭飼養(仍用黃金榕葉片飼養)直至化蛹和羽化,采用相同的方法分別依次觀察各齡期幼蟲(n=213~275)、蛹(n=203)和成蟲(n=194)的形態特征并拍照。同時,利用Image Measure軟件(VMS3.5-MUC1121)測量卵的直徑、幼蟲的頭殼寬度及體長、蛹寬及成蟲翅展等。
1.3 年生活史的觀察
將50對未交配的雌雄成蟲置于倒扣在培養皿的塑料杯(V=125 mL,1對/杯)中讓其自由交配。待觀察到雌蟲將卵產在塑料杯內壁后,更換新塑料杯,并將附著有卵粒的塑料杯置于一圓形塑料盒(d=25 cm;h=15 cm)中,在室溫下待其自然孵化。待幼蟲孵化后,用毛筆將初孵幼蟲挑至培養皿中用黃金榕葉片單頭飼養,每天更換1次食料。于自然溫度下連續飼養,記錄每代每頭幼蟲的蛻皮和化蛹時間、成蟲羽化和死亡時間,確定其年發生世代數。
1.4 生活習性的觀察
將50對成蟲置于放有盆栽黃金榕的網籠(長×寬×高=50 cm×50 cm×80 cm)中(10對/籠罩)讓其自由交配和產卵,幼蟲孵化后繼續讓其為害黃金榕,觀察其產卵時間、產卵部位、幼蟲孵化時間、為害部位、為害特征、活動習性、化蛹場所、羽化時間和交配時間等生活習性。其中,產卵、幼蟲孵化、羽化和交配每隔1 h觀察1次。此外,于2013年5月至2013年10月在廣西大學校園內、市區綠化帶、獅山公園、金花茶公園、南湖公園和青秀山公園等地選取蟲口密度較大的榕樹喬木和灌木綠化帶,在其內設置固定調查樣地,隨機選取30株有蟲喬木和灌木,每月調查2次,記錄該蟲的為害特征、為害部位、生活習性、活動情況、寄主和天敵種類等。寄生性天敵根據文獻資料進行鑒定[6-7]。
2 結果與分析
2.1 形態特征
成蟲:雌蟲體長10~15 mm,翅展32~39 mm。觸角雙櫛齒狀,黑色,鞭節灰白色,支角突外裹淡橘紅色絨毛,圍角片灰白色;復眼發達,黑色;具黃色可伸卷長喙;翅橘紅色。前翅和后翅紅色,臀區有一黑色橢圓形斑;胸部背面及腹部兩側被有深紅色體毛,胸腹部的腹面體毛為黑色,節間膜為金黃色,偶有蜜紅色;足的轉節和腿節內側被有灰白絨毛。雄蟲體型較雌蟲略小,體長9~13 mm,翅展25~33 mm。觸角雙櫛齒狀,鞭節黑色,胸腹部腹面的體毛為灰白色。腹部末端露出1對黑色毛須,其余特征與雌蟲相同(圖1a)。
卵:卵圓形,直徑0.7~1.0 mm,表面覆有黃色膠質,卵粒聚集排列呈連續線狀,初產時淺黃色,孵化前呈深黃色(圖1b)。
幼蟲:幼蟲共6個齡期,各齡期幼蟲的體長及頭殼寬度見表1。幼蟲頭部蛻裂線白色,呈“倒Y”字形,額區呈“△”形,頂端淡黑色,中部土黃色,底部(額唇基區)白色,且中部和額唇基區交界處的左右兩邊各著生1個小突起,突起頂端白色,其上著生1根淡黃色剛毛。前胸背板呈長方形,黃褐色,具黑色斑點,中胸似“盾”狀,深栗色,后胸同為深栗色,與中胸等高,但較中胸更寬,是整個體區最寬的體節。腹部共7節,腹足4對,分別著生在第2、3、4和5腹節上,臀足1對,著生在第7腹節上。腹部整個背面紅棕色(圖1c)。
蛹:被蛹,長10~12 mm,寬2.8~4.8 mm,紡錘形,背面黑褐色,其余均呈淡黃色(化蛹初期),后期漸變紅(羽化前期),翅芽達第5腹節末端(圖1d)。蛹室外觀形同船狀,土黃色,尾端有羽化孔。整個蛹室呈長橢圓形,長15~19 mm,寬6~9 mm。
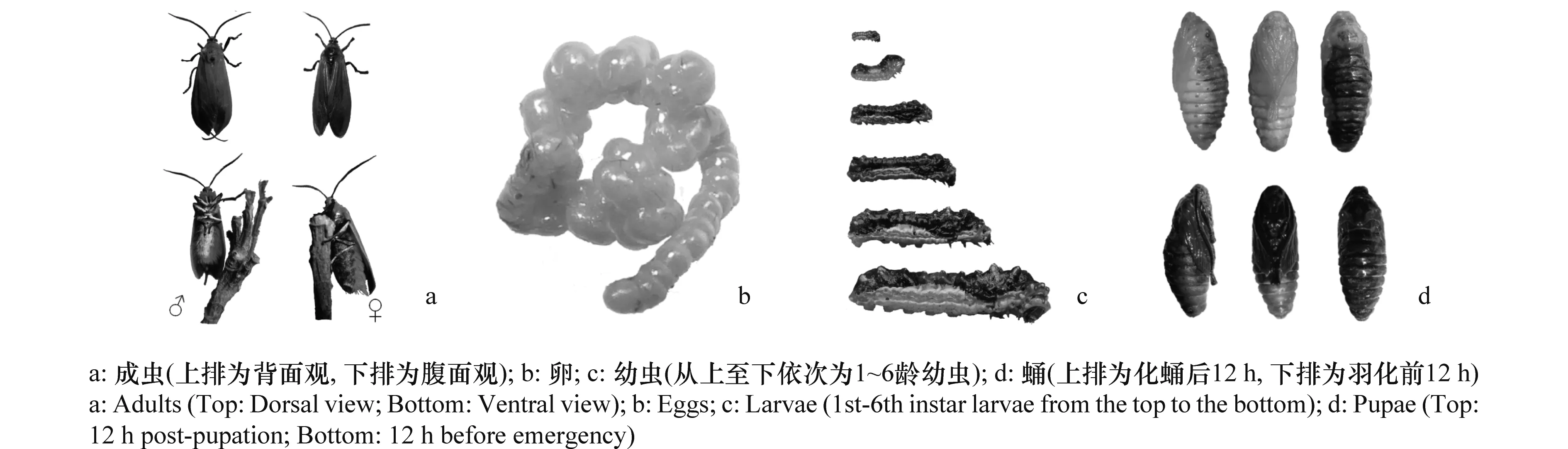
圖1 朱紅毛斑蛾成蟲、卵、幼蟲和蛹的形態特征Fig.1 Morphological characteristics of adults, eggs, larvae and pupae of Phauda flammans

齡期Instars體長/mmBodylength頭殼寬度/mmWidthofheadcapsule12.7±0.90.6±0.127.2±1.20.9±0.139.2±1.51.2±0.2416.8±2.21.6±0.2520.1±2.62.1±0.2623.0±3.93.3±0.3
1) 表中體長和頭寬的數據為平均值±標準誤。
Data are showed as mean±SE in the table.
2.2 年生活史
朱紅毛斑蛾在廣西南寧一年發生2~3代(表2),以部分第2代預蛹期幼蟲和蛹及第3代低齡幼蟲越冬。每年3月中旬到5月上旬為越冬代幼蟲的為害期,4月下旬到7月中旬為第1代幼蟲的為害期,7月下旬到11月中旬為第2代幼蟲的為害期,10月中旬到11月中旬為第3代幼蟲的為害期。11月下旬開始以預蛹期幼蟲或蛹越冬。翌年3月中下旬到5月幼蟲化蛹和越冬蛹陸續羽化。4月下旬成蟲開始產卵,第1代幼蟲開始孵化,5月中旬-6月下旬為第1代幼蟲發生盛期,6月下旬-7月下旬為第1代成蟲的羽化盛期,7月中下旬為產卵盛期;第2代幼蟲于8月上旬-10月中旬為盛發期,9月中旬-10月中旬為化蛹高峰期,9月下旬-10月下旬為羽化高峰期,10月為產卵高峰期,室內統計結果顯示,35.4%的第2代幼蟲進入預蛹期或蛹期后當年不再化蛹或羽化。第3代幼蟲孵化后以低齡幼蟲越冬,直至翌年4月上旬開始化蛹。第2代和第3代幼蟲存在世代重疊現象(表2)。
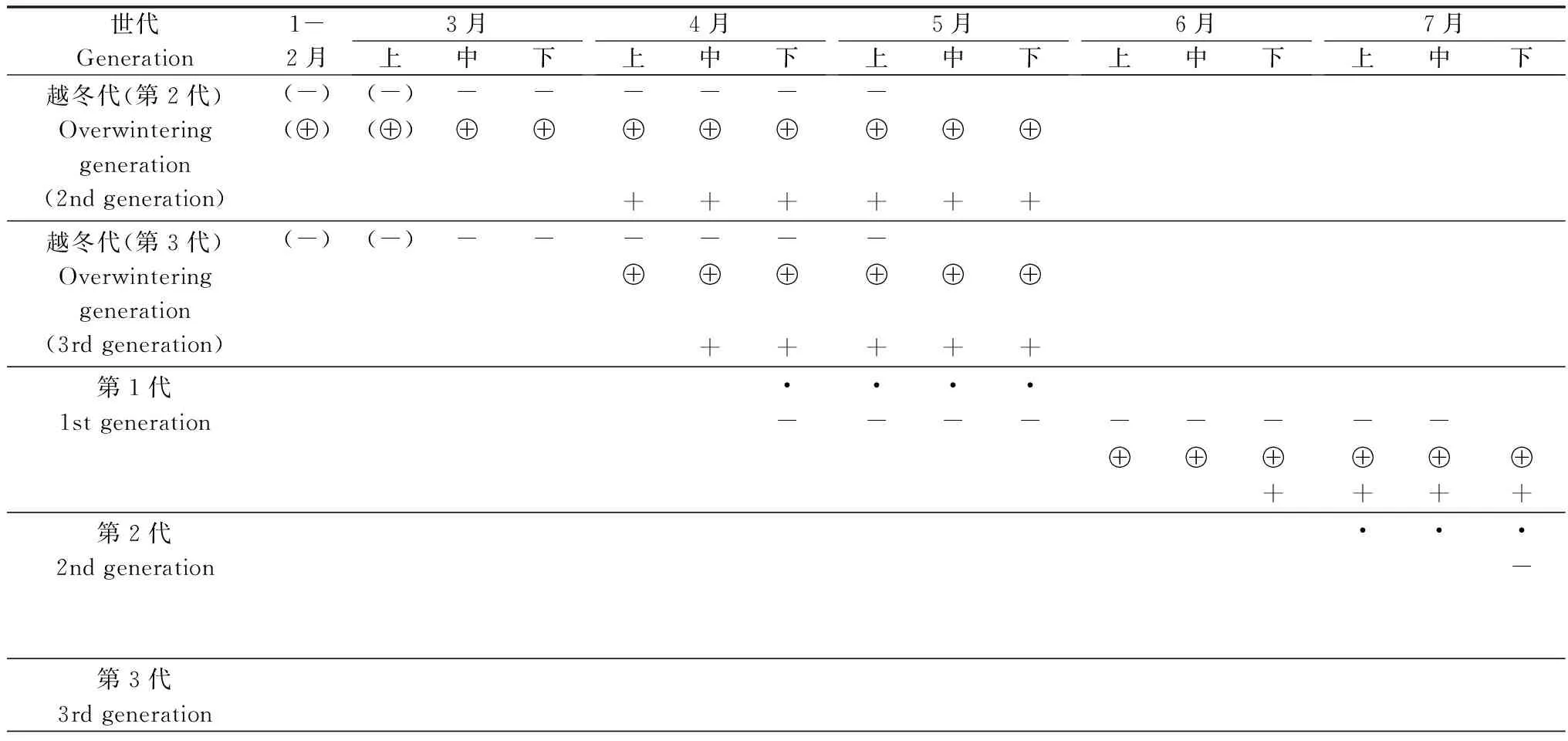
表2 朱紅毛斑蛾年生活史(廣西南寧,2012-2014)1)
續表2 Table 2(Continued)

世代Generation8月上中下9月上中下10月上中下11月上中下12月上中下越冬代(第2代)Overwinteringgeneration(2ndgeneration)越冬代(第3代)Overwinteringgeneration(3rdgeneration)第1代1stgeneration⊕+第2代2ndgeneration·------------(-)(-)(-)(-)⊕⊕⊕⊕⊕⊕⊕⊕⊕⊕(⊕)(⊕)(⊕)(⊕)+++++++++++第3代3rdgeneration·········----(-)(-)(-)(-)
1) ·卵;-幼蟲;(-)越冬幼蟲;⊕蛹;(⊕)越冬蛹;+成蟲。
·: Eggs;-: Larvae; (-): Overwintering larvae; ⊕: Pupae; (⊕): Overwintering pupae; +: Adults.
2.3 生活習性
朱紅毛斑蛾成蟲性比為1∶1.14(n=2 321,含室內羽化及野外調查的成蟲)。晝夜節律中,成蟲羽化高峰期為10:00-13:00(n=127),室內自然氣溫條件下羽化率為80.9%;交配高峰期為13:00-18:00(n=88),交配時長3~12 h;雌雄蟲交尾一次,未見多次交配現象,交尾時雌雄蛾腹末相對,呈“一”字型;產卵高峰期為11:00-15:00(n=44)。成蟲受驚擾時,腹部突然翹起或(同時)假死落地。卵多產于葉片的正面或榕樹枝條上,單雌產卵量為78.3±13.8粒,卵粒聚集呈不規則連續線狀,自然孵化率達95%以上,偶有未交配雌蟲產卵現象,但均不能孵化。幼蟲晝夜均可取食,可為害用作行道樹和綠籬的榕樹(圖2a~c),1~2齡幼蟲僅啃食葉肉(圖2d),留下白色膜狀上表皮,隨著蟲齡的增長,葉片被害狀呈缺刻或孔洞(圖2e)。嚴重為害時,整株葉片均被蠶食僅剩枝干,更有少數高齡幼蟲在無葉片時取食榕樹韌皮部(圖2f)。自然條件下,幼蟲行動緩慢,受驚嚇時頭部緊縮于前胸背板下并從口器中分泌黃褐色液體。老熟幼蟲化蛹前停止取食,起初分泌無色或紅褐色透明狀液體,常由樹干向下爬行,吐絲結繭化蛹,羽化孔開于尾部,化蛹部位位于地表突出的根系附近或雜草叢下,尤其喜歡在根系與地表形成的夾角處結繭化蛹,極少數在土壤表層0~10 mm處做蛹室化蛹;蛹期受溫度影響較大,5-9月蛹期11~20 d,10月至翌年4月蛹期21~38 d。野外調查發現,朱紅毛斑蛾可為害小葉榕(F.microcarpavar.pusillifolia)、黃金榕、垂葉榕(F.benjamina),但未見為害其他樹種,推測朱紅毛斑蛾為寡食性害蟲,但仍需進一步證實。此外,調查還發現了朱紅毛斑蛾的兩種寄生性天敵,一種為花胸姬蜂(GotraoctocinctusAshmead)(膜翅目:姬蜂科)(圖3a~b),另一種為盤絨繭蜂屬種類Cotesiasp.(膜翅目:繭蜂科)(圖3c~d),均寄生朱紅毛斑蛾幼蟲,其中花胸姬蜂為優勢種。
3 討論
朱紅毛斑蛾是我國南方榕樹重要的食葉害蟲之一,近幾年其發生呈逐步加重趨勢,嚴重影響市容市貌。20世紀80年代,董祖林和伍有聲報道該蟲在廣州1年發生2代[2],本文研究結果與其并不完全一致,是否與地理環境、寄主等因素相關仍需進一步研究。之后,未見有朱紅毛斑蛾相關研究的報道,由于缺乏對其生物學及生態學等特性的了解阻礙了其防控工作的開展。本文研究結果顯示,朱紅毛斑蛾在廣西南寧地區1年發生2~3代,5月中旬-6月下旬及8月上旬-10月中旬分別為第1和2代幼蟲發生盛期,為害最重。因此,在這兩個幼蟲高發期間做好防控工作是控制該蟲的關鍵。此外,越冬蛹多于地表結繭化蛹越冬,極易觀察,可結合平時園林工人的管理集中收集銷毀,以消滅其越冬蟲源,降低翌年的蟲口基數。目前,僅發現寄生朱紅毛斑蛾的兩種寄生蜂,是否存在其他天敵昆蟲以及這兩種寄生蜂的自然防控效果如何尚待進一步調查研究。
[1] 陳根富. 福州榕樹害蟲發生規律及生活習性的考察[J].福建師范大學學報(自然科學版),1989, 5(4):86-89.
[2] 董祖林,伍有聲. 榕樹斑蛾生活習性的初步觀察及防治[J].廣東園林,1982 (4):43-44.
[3] 羅實謙,羅娜. 小葉榕樹的主要害蟲鑒定及其危害特點[J].現代園藝,2011 (5):90-91.
[4] 張倫全,方志美. 小葉榕薊馬種類調查及防治試驗初報[J].四川林業科技,2002, 23(4):63-65.
[5] 鄭月瓊. 泉州市榕樹主要害蟲鑒定及其為害特點[J].福建農業科技,2010 (4):64-65.
[6] 何俊華. 浙江蜂類志[M].北京:科學出版社,2004: 515.
[7] 陳家驊,宋東寶. 中國小腹繭蜂(膜翅目:繭蜂科)[M].福州:福建科學技術出版社,2004: 143-174.
(責任編輯:楊明麗)
Biological characteristics ofPhaudaflammans(Lepidoptera: Zygaenidae)
Liu Junyan, He Qiulong, Wei Hang, Yang Juan, Li Jun, Lu Wen, Zheng Xialin
(College of Agriculture, Guangxi University, Nanning 530004, China)
Phaudaflammans(Lepidoptera: Zygaenidae) is one of the most serious defoliating pests ofFicus. Morphology, life history and habit were investigated based on the laboratory observation and field investigation. Results showed that eggs aggregated as the form of unbroken linearity, and larvae had six instars. Differences of male and female moths could be distinguished from the color of flagellum, with or without cercus at the end of the abdomen, and the color of hair on the segmental venter. Pupal color usually changed from buff to red. This species had 2-3 generations per year in Guangxi Province, China. Larval peak of the first and second generations occurred from mid May to late June and early August to mid October. The moth overwintered as the pre-pupae larvae and pupae of the second generation and young larvae of the third generation. Larvae of the second and third generations overlapped. Diurnal peaks of emergence, mating, and oviposition were 10: 00-13:00,13: 00-18:00, and 11:00-15:00, respectively. Moth could abruptly fall to the ground or abdomen lift when they were disturbed. Larvae could damageFicusleaves, but phloem was also attacked when they were lack of food. Most larvae preferred to pupate for overwintering near root exposed on the ground and tussock, only few individuals could overwinter at the 0-10 mm soil depth. Under natural conditions, two parasitic wasps were found. However, the control effect of these parasitic wasps needs to evaluate further.
Phaudaflammans; morphology; life history; life habit
2014-03-16
2014-05-24
廣西大學科研基金項目(XJZ130369)
S 436.8
A
10.3969/j.issn.0529-1542.2015.03.037
* 通信作者 E-mail:zheng-xia-lin@163.com
