小干擾RNA沉默脂肪酸合酶基因?qū)θ烁渭?xì)胞株HepG2脂代謝的影響
2015-12-16 03:38:31張秀云馮艷萍熊東葉文靜朱粒仁范阿強(qiáng)陳建業(yè)宋永硯
中國(guó)循環(huán)雜志 2015年7期
關(guān)鍵詞:水平
張秀云,馮艷萍,熊東,葉文靜,朱粒仁,范阿強(qiáng),陳建業(yè),宋永硯
小干擾RNA沉默脂肪酸合酶基因?qū)θ烁渭?xì)胞株HepG2脂代謝的影響
張秀云,馮艷萍,熊東,葉文靜,朱粒仁,范阿強(qiáng),陳建業(yè),宋永硯
目的:研究基因干擾技術(shù)沉默脂肪酸合酶(FAS)基因后對(duì)人肝細(xì)胞株HepG2細(xì)胞內(nèi)外脂質(zhì)含量及脂代謝相關(guān)基因表達(dá)的影響。
方法:合成3對(duì)針對(duì)FAS mRNA不同位點(diǎn)序列的小干擾RNA(siRNA),分別轉(zhuǎn)染至人肝細(xì)胞株HepG2,實(shí)時(shí)熒光定量聚合酶鏈反應(yīng)(PCR)檢測(cè)空白對(duì)照組(HepG2細(xì)胞不經(jīng)過(guò)任何處理)、陰性對(duì)照組(轉(zhuǎn)染沒(méi)有任何基因干擾作用的陰性siRNA)、FAS-siRNA-1轉(zhuǎn)染組、FAS-siRNA-2轉(zhuǎn)染組和FAS-siRNA-3轉(zhuǎn)染組的FAS mRNA的表達(dá)。蛋白免疫印跡(Western blot)法檢測(cè)蛋白質(zhì)的表達(dá)水平,篩選出沉默效果最佳的一對(duì)siRNA。將最有效siRNA轉(zhuǎn)染至HepG2細(xì)胞,48 h后測(cè)定細(xì)胞內(nèi)外甘油三酯及總膽固醇含量以及脂代謝相關(guān)基因的mRNA表達(dá)水平。
結(jié)果:25 nmol/L siRNA和3 μl/孔(24孔板)轉(zhuǎn)染試劑的轉(zhuǎn)染效率最高。FAS-siRNA-3對(duì)HepG2的FAS基因的沉默效果最好,F(xiàn)AS-siRNA-3轉(zhuǎn)染組的FAS在mRNA水平和蛋白水平分別降低為空白對(duì)照組的(52.33±3.07)%和(51.57±3.14)%。FAS-siRNA-3沉默F(xiàn)AS基因后HepG2的細(xì)胞內(nèi)和細(xì)胞外培養(yǎng)液中甘油三酯含量顯著低于空白對(duì)照組,細(xì)胞外總膽固醇含量顯著低于空白對(duì)照組。與空白對(duì)照組比較,F(xiàn)AS-siRNA-3轉(zhuǎn)染組HepG2的肝脂酶基因mRNA表達(dá)水平顯著升高(P<0.0001),微粒體甘油三酯轉(zhuǎn)運(yùn)蛋白基因mRNA表達(dá)水平顯著降低(P<0.05),差異均有統(tǒng)計(jì)學(xué)意義。
結(jié)論:FAS基因沉默可顯著降低HepG2細(xì)胞內(nèi)甘油三酯、細(xì)胞外培養(yǎng)液甘油三酯和總膽固醇的含量,需進(jìn)一步的體內(nèi)外實(shí)驗(yàn)加以驗(yàn)證。
小干擾RNA ; 脂肪酸合酶 ;人肝細(xì)胞株HepG2
Methods: A total of 3 pairs of small interfering RNA (siRNA) targeting different sequences of FAS mRNA were synthesized as FAS-siRNA-1, FAS-siRNA-2 and FAS-siRNA-3, meanwhile, 2 controls were established as Blank control group, in which HepG2 cells were not treated, and Negative control group, in which HepG2 cells were transfected by noneffective siRNA. The mRNA, and protein expression levels of FAS in HepG2 cells were examined by real-time fluorescence quantitative RCR and Western blot analysis to screen the most effective pair of siRNA for FAS gene silencing; and that specific siRNA was transtected to HepG2 cells for 48 hours to detect the intra-/extra-cellular TG, TC levels and the mRNA expression related to lipid metabolism in HepG2 cells.
Results: The screening experiment indicated that FAS-siRNA-3 was most effective for FAS gene silencing. Compared with Blank control group, the mRNA and protein expressions in FAS-siRNA-3 transfected HepG2 cells (Transfected group)
decreased to (52.33 ± 3.07) % and (51.57 ± 3.14) % respectively. Compared with Blank control group, Transfected group had the reduced intra-/extra-cellular TG levels and reduced extracellular TC level; while increased mRNA expression of hepatic lipase, P<0.0001 and decreased mRNA expression of TG transfer protein in HepG2 microsome, P<0.05.
Conclusion: FAS gene silencing could significantly decrease the intra-/extra- cellular TG level and extracellular TC level in HepG2 cells, those findings need to be confirmed by further in vivo and in vitro studies.
(Chinese Circulation Journal, 2015,30:670.)
高甘油三酯血癥(HTG)是動(dòng)脈粥樣硬化的獨(dú)立危險(xiǎn)因子,與心血管病(CVD)直接相關(guān)[1-6]。血漿甘油三酯主要來(lái)源于肝臟的從頭合成(de novo synthesis)。在肝甘油三酯從頭合成過(guò)程中,脂肪酸合酶(FAS)是起關(guān)鍵作用的多功能酶,其表達(dá)水平及活性決定了甘油三酯的合成速率以及血漿甘油三酯的水平[7]。重酒石酸煙堿誘導(dǎo)使大鼠肝FAS表達(dá)增強(qiáng)后,肝臟及血漿中甘油三酯水平顯著升高[8]。FAS抑制劑可以有效降低大鼠血漿甘油三酯水平以及逆轉(zhuǎn)非酒精性脂肪肝病[9,10]。本研究運(yùn)用RNA干擾技術(shù),特異性沉默HepG2細(xì)胞FAS表達(dá),在細(xì)胞及分子水平上觀(guān)察FAS沉默后肝細(xì)胞內(nèi)外脂質(zhì)含量以及脂代謝相關(guān)基因表達(dá)的改變,為相關(guān)體內(nèi)研究的開(kāi)展提供實(shí)驗(yàn)基礎(chǔ)。
1 材料與方法
主要實(shí)驗(yàn)材料:試驗(yàn)時(shí)間:2014-07至2014-10,本研究涉及主要儀器和試劑包括:人肝細(xì)胞株HepG2從武漢博士德購(gòu)買(mǎi);細(xì)胞培養(yǎng)箱(美國(guó)Thermo);實(shí)時(shí)熒光定量聚合酶鏈反應(yīng)(PCR)儀(美國(guó)ABI 7900HT);轉(zhuǎn)染試劑TransIT-siQUEST(美國(guó)Mirus);細(xì)胞總RNA提取試劑盒(北京天根);熒光定量試劑盒(日本TaKaRa);細(xì)胞總蛋白提取及定量試劑盒(上海碧云天);FAS兔抗人一抗(美國(guó)GeneTex);蛋白免疫印跡(Western blot)試劑盒(上海碧云天);無(wú)血清培養(yǎng)液Opti-MEM(美國(guó)Gibco);細(xì)胞內(nèi)甘油三酯/總膽固醇測(cè)定試劑盒(北京普利萊);血清甘油三酯/總膽固醇測(cè)定試劑盒(浙江東甌)。小干擾RNA(siRNA)由上海吉碼制藥技術(shù)有限公司設(shè)計(jì)合成,3對(duì)siRNA及陰性對(duì)照序列分別為:FAS-siRNA-1:(280)GGACCUGUCUAGGUUUGAUTT; FAS-siRNA-2:(3072)CCCAGGCUGAAGUUUACAATT; FAS-siRNA-3:(7377)GGUCCUUCUACUACAAGCUTT; 5’-UUCUCCGAACGUGUCACGUTT-3’。
細(xì)胞培養(yǎng):HepG2細(xì)胞采用Iscove's改良的Eagle培養(yǎng)基(IMEM)培養(yǎng),含10%胎牛血清、2 000 μmol/L L-谷氨酰胺及雙抗(100 U/ml青霉素、100 μg/ml鏈霉素)。細(xì)胞置37℃,5% CO2培養(yǎng)箱中培養(yǎng),生長(zhǎng)至對(duì)數(shù)生長(zhǎng)期的細(xì)胞用于實(shí)驗(yàn)。
轉(zhuǎn)染條件的優(yōu)化:HepG2細(xì)胞接種于24孔板培養(yǎng),細(xì)胞融合度長(zhǎng)至50%~70%時(shí)進(jìn)行轉(zhuǎn)染。將轉(zhuǎn)染試劑(1.5 μl/孔或3 μl/孔)和熒光標(biāo)記的陰性對(duì)照組siRNA(25、50或75 nmol/L)與無(wú)血清培養(yǎng)液(Opti-MEM)混合、靜置形成轉(zhuǎn)染復(fù)合物。轉(zhuǎn)染復(fù)合物的配制過(guò)程為:吸取50μl Opti-MEM培養(yǎng)液,加入轉(zhuǎn)染試劑并輕輕混勻。然后加入陰性對(duì)照組儲(chǔ)存液,移液槍反復(fù)輕輕吹打混勻。室溫下靜置20 min形成轉(zhuǎn)染復(fù)合物。將轉(zhuǎn)染復(fù)合物“十”字形滴加到細(xì)胞培養(yǎng)孔中,輕輕晃動(dòng)培養(yǎng)板使之混合均勻,放回培養(yǎng)箱繼續(xù)培養(yǎng),6 h后熒光顯微鏡拍照計(jì)數(shù)。根據(jù)熒光細(xì)胞數(shù)占總細(xì)胞數(shù)的比例確定最佳轉(zhuǎn)染條件。
最有效siRNA的篩選:以轉(zhuǎn)染條件優(yōu)化中得出的最佳轉(zhuǎn)染條件將陰性對(duì)照組及3對(duì)siRNA分別轉(zhuǎn)染進(jìn)入HepG2細(xì)胞。轉(zhuǎn)染后48 h,實(shí)時(shí)熒光定量PCR分別檢測(cè)空白對(duì)照組(HepG2細(xì)胞不經(jīng)過(guò)任何處理)、陰性對(duì)照組(轉(zhuǎn)染沒(méi)有任何基因干擾作用的陰性siRNA)、FAS-siRNA-1轉(zhuǎn)染組、FAS-siRNA-2轉(zhuǎn)染組和FAS-siRNA-3轉(zhuǎn)染組的FAS mRNA的表達(dá)。FAS上游引物為5’-AAGGGCCTAGAGGAGCGTGT-3’,下游引物為5’-TGGTACTTGGCCTTGGGTGT’,產(chǎn)物大小為149bp;內(nèi)參照GAPDH上游引物為5’-CTTTGGTATCGTGGAAGGACTC-3’,下游引物為5’- GTAGAGGCAGGGATGATGTTCT-3’,產(chǎn)物大小為132bp。定量反應(yīng)體系:上、下游引物各0.5 μl,cDNA 1.0 μl,SYBR Premix Ex Taq 10 μl,dH2O 8.0 μl,總體積20 μl。反應(yīng)條件:95℃預(yù)變性15s,95℃ 5s,60℃ 30s,45次循環(huán)。溶解曲線(xiàn)95℃ 5s,55℃ 60s,55℃~95℃30s。根據(jù)2-ΔΔCt法計(jì)算樣本的mRNA含量。
Western blot法檢測(cè)細(xì)胞中FAS的表達(dá)量:嚴(yán)格按照試劑盒說(shuō)明書(shū)提取細(xì)胞總蛋白并進(jìn)行定量,蛋白變性后行十二烷基磺酸鈉(SDS)—聚丙烯酰胺凝膠電泳,轉(zhuǎn)膜、封閉后,各組先后與兔抗人FAS一抗以及辣根過(guò)氧化物酶HRP標(biāo)記的二抗孵育,洗膜后化學(xué)發(fā)光法成像。采用Image lab 3.0分析軟件對(duì)條帶灰度值進(jìn)行測(cè)定并分析。
FAS-siRNA-3處理對(duì)HepG2細(xì)胞內(nèi)外脂質(zhì)含量的影響:以最佳轉(zhuǎn)染條件和最有效siRNA(FAS-siRNA-3)轉(zhuǎn)染HepG2細(xì)胞。轉(zhuǎn)染后48 h,同時(shí)收集細(xì)胞和上清液。細(xì)胞內(nèi)甘油三酯和總膽固醇含量測(cè)定參照北京普利萊基因技術(shù)有限公司測(cè)定試劑盒說(shuō)明書(shū)進(jìn)行:細(xì)胞加入裂解液充分吹打裂解,12 000×g、4℃離心10 min。取上清,分成兩份,一份真空冷凍抽干后采用細(xì)胞甘油三酯和總膽固醇測(cè)定試劑盒測(cè)定甘油三酯和總膽固醇含量;另一份采用二喹啉甲酸(BCA)法測(cè)定蛋白質(zhì)濃度(參照上海碧云天生物技術(shù)有限公司的蛋白質(zhì)濃度測(cè)定試劑盒說(shuō)明書(shū)進(jìn)行),結(jié)果以每g蛋白質(zhì)對(duì)應(yīng)的細(xì)胞內(nèi)甘油三酯或總膽固醇含量(mmol/g)表示。細(xì)胞培養(yǎng)上清液甘油三酯和總膽固醇含量測(cè)定:取1 ml上清液真空冷凍抽干,采用血清甘油三酯和總膽固醇測(cè)定試劑盒測(cè)定490 nm處的吸光度,根據(jù)標(biāo)準(zhǔn)液的吸光度值求出樣品溶液的甘油三酯及總膽固醇含量。每個(gè)處理重復(fù)3次,均值用于統(tǒng)計(jì)分析。
FAS-siRNA-3處理對(duì)HepG2細(xì)胞脂代謝相關(guān)基因表達(dá)的影響:以最佳轉(zhuǎn)染條件和最有效siRNA(FAS-siRNA-3)轉(zhuǎn)染HepG2細(xì)胞。轉(zhuǎn)染后48 h,裂解細(xì)胞提取總RNA,實(shí)時(shí)熒光定量PCR檢測(cè)肝脂酶基因(HL)、3-羥基-3-甲基戊二酸單酰輔酶A還原酶基因(HMGR)、固醇調(diào)節(jié)元件結(jié)合蛋白-1基因(SREBP1)、微粒體甘油三酯轉(zhuǎn)運(yùn)蛋白基因(MTP)、載脂蛋白B100基因(APOB100)的mRNA表達(dá)水平。每個(gè)處理重復(fù)3次,均值用于統(tǒng)計(jì)分析。

2 結(jié)果
轉(zhuǎn)染條件的優(yōu)化(圖1):轉(zhuǎn)染后6 h,根據(jù)熒光細(xì)胞數(shù)占總細(xì)胞數(shù)的比例計(jì)算,1.5 μl/孔(24孔板)轉(zhuǎn)染試劑×25 nmol/L siRNA、1.5 μl/孔轉(zhuǎn)染試劑×50 nmol/L siRNA、1.5μl /孔轉(zhuǎn)染試劑×75 nmol/L siRNA、3 μl/孔轉(zhuǎn)染試劑×25nmol/L siRNA、3 μl/孔轉(zhuǎn)染試劑×50 nmol/L siRNA和3 μl/孔轉(zhuǎn)染試劑×75 nmol/L siRNA的轉(zhuǎn)染率分別為43.0%、35.5%、49.0%、65.0%、32.5.0%和50.0%,因此確定3 μl/孔轉(zhuǎn)染試劑×25 nmol/L siRNA的轉(zhuǎn)染效率最高。

圖1 25 nmol/L熒光標(biāo)記的小干擾RNA轉(zhuǎn)染HepG2細(xì)胞6 h后的熒光照片(×200)
最有效siRNA的篩選:實(shí)時(shí)熒光定量PCR檢測(cè)發(fā)現(xiàn)FAS-siRNA-1和FAS-siRNA-3對(duì)FAS基因的沉默效果較好。與空白對(duì)照組(100±7.89)%比較,陰性對(duì)照組的FAS mRNA水平為(96.79±20.26)%。FAS-siRNA-1轉(zhuǎn)染組、FAS-siRNA-2轉(zhuǎn)染組和FAS-siRNA-3轉(zhuǎn)染組的mRNA水平分別降低為空白對(duì)照組的(52.57±2.36)%、(61.42±5.23)%和(52.33±3.07)%,與空白對(duì)照組和陰性對(duì)照組比較差異均有統(tǒng)計(jì)學(xué)意義(P<0.0001)。通過(guò)Western blot法檢測(cè)細(xì)胞內(nèi)FAS的表達(dá)量,與空白對(duì)照組比較,F(xiàn)AS-siRNA-1轉(zhuǎn)染組、FAS-siRNA-2轉(zhuǎn)染組和FAS-siRNA-3轉(zhuǎn)染組的FAS含量分別降低為空白對(duì)照組的(64.34±7.85)%、(69.74±11.61)%和(51.57±3.14)%,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05)(圖2)。根據(jù)實(shí)時(shí)熒光定量PCR和Western blot檢測(cè)結(jié)果確定FAS-siRNA-3對(duì)FAS基因沉默效果最好,為最佳干擾siRNA,用于后續(xù)實(shí)驗(yàn)。

圖2 蛋白免疫印跡法檢測(cè)不同處理下脂肪酸合酶的蛋白質(zhì)表達(dá)水平
FAS-siRNA-3處理對(duì)HepG2 48 h后細(xì)胞內(nèi)外脂質(zhì)含量的影響(表1): FAS-siRNA-3轉(zhuǎn)染組細(xì)胞內(nèi)甘油三酯含量顯著低于空白及陰性對(duì)照組(P<0.05),細(xì)胞外培養(yǎng)液中甘油三酯和總膽固醇含量顯著低于空白及陰性對(duì)照組(P<0.05),差異均有統(tǒng)計(jì)學(xué)意義。細(xì)胞內(nèi)總膽固醇含量在各組之間差異無(wú)統(tǒng)計(jì)學(xué)意義。
表1 FAS-siRNA-3處理HepG2細(xì)胞48h后細(xì)胞內(nèi)外甘油三酯和總膽固醇的相對(duì)百分含量
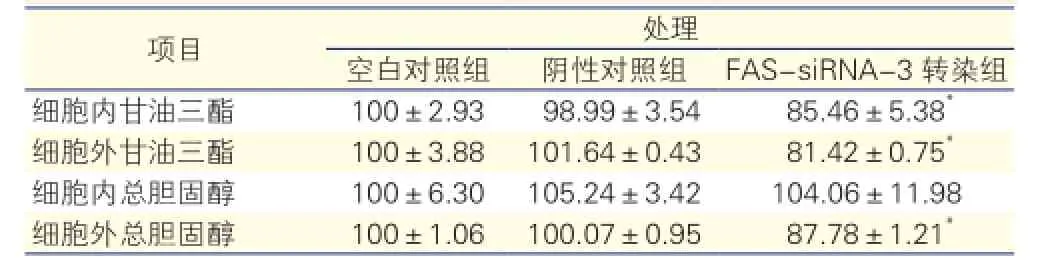
表1 FAS-siRNA-3處理HepG2細(xì)胞48h后細(xì)胞內(nèi)外甘油三酯和總膽固醇的相對(duì)百分含量
注:FAS:脂肪酸合酶;siRNA:小干擾RNA。與空白對(duì)照組和陰性對(duì)照組相比*P<0.05
?
FAS-siRNA-3處理對(duì)HepG2細(xì)胞脂代謝相關(guān)基因表達(dá)的影響(表2):與空白及陰性對(duì)照組比較,F(xiàn)AS-siRNA-3轉(zhuǎn)染組HL基因表達(dá)顯著增強(qiáng)(P<0.0001),MTP基因表達(dá)水平顯著降低(P<0.05),差異均有統(tǒng)計(jì)學(xué)意義。HMGR、SREBP1和APOB1000的表達(dá)水平在各組之間差異無(wú)統(tǒng)計(jì)學(xué)意義。
表2 FAS-siRNA-3處理HepG2細(xì)胞48 h后脂代謝相關(guān)基因mRNA的相對(duì)百分含量

表2 FAS-siRNA-3處理HepG2細(xì)胞48 h后脂代謝相關(guān)基因mRNA的相對(duì)百分含量
注:HL:肝脂酶基因;HMGR:3-羥基-3-甲基戊二酸單酰輔酶A還原酶基因;SREBP1:固醇調(diào)節(jié)元件結(jié)合蛋白-1基因;MTP:微粒體甘油三酯轉(zhuǎn)運(yùn)蛋白基因;APOB100:載脂蛋白B100基因;與空白對(duì)照組和陰性對(duì)照組相比*P<0.05**P<0.0001。余注見(jiàn)表1
?
3 討論
FAS是催化脂肪酸合成的關(guān)鍵酶, 已成為治療高脂血癥、肥胖、腫瘤等疾病的重要靶標(biāo)[11,12]。在人類(lèi)細(xì)胞中,F(xiàn)AS是含有兩個(gè)相同亞基的同源二聚體,兩亞基首尾相接,構(gòu)成一個(gè)環(huán)狀的多酶復(fù)合體。FAS每個(gè)亞基大小約為270 kD,含有乙酰轉(zhuǎn)移酶等7種酶活性,依次以乙酰輔酶A和丙二酸單酰輔酶A為原料合成16碳的軟脂酸,軟脂酸進(jìn)一步延長(zhǎng)和酯化成甘油三酯。人FAS定位于染色體17q25,含有42個(gè)內(nèi)含子和43個(gè)外顯子,長(zhǎng)約22 kb。FAS普遍表達(dá)于全身各組織中,在肝臟的表達(dá)量最高,其脂肪酸合成能力是脂肪組織的8~9倍[13]。
血液甘油三酯主要來(lái)源于肝臟的從頭合成。肝細(xì)胞合成的甘油三酯以極低密度脂蛋白(VLDL)的形式釋放入血。FAS在肝臟中的表達(dá)水平及活性與血漿甘油三酯水平顯著相關(guān)。Ma等[8]給雌性Wistar大鼠注射重酒石酸煙堿[1mg/(kg·d)]2周后,肝臟FAS表達(dá)顯著增強(qiáng),同時(shí)肝臟及血漿中的甘油三酯水平顯著增高。與之對(duì)應(yīng),大豆異黃酮干預(yù)非酒精性脂肪肝模型大鼠12周后,肝FAS表達(dá)水平顯著降低,同時(shí)肝臟及血清中的甘油三酯顯著降低[14]。
基于FAS在高甘油三酯血癥發(fā)病中的重要作用,F(xiàn)AS很可能成為臨床治療高甘油三酯血癥的一個(gè)重要靶點(diǎn),抑制FAS基因表達(dá)也很可能成為治療高甘油三酯血癥的重要手段。
RNA干擾技術(shù)根據(jù)靶基因mRNA序列設(shè)計(jì)互補(bǔ)的siRNA,體外合成后導(dǎo)入細(xì)胞或體內(nèi),在細(xì)胞內(nèi)siRNA與一些酶(包括內(nèi)切酶、外切酶、解旋酶等)結(jié)合形成RNA誘導(dǎo)的沉默復(fù)合物(RISC)。RISC識(shí)別和結(jié)合靶基因mRNA的同源區(qū)域,并在結(jié)合區(qū)內(nèi)切割mRNA,導(dǎo)致靶基因功能缺失,從而達(dá)到疾病治療的目的。RNA干擾技術(shù)已廣泛應(yīng)用于高脂血癥相關(guān)基因的沉默[15-19]。本研究首次運(yùn)用siRNA技術(shù)沉默F(xiàn)AS基因。研究結(jié)果顯示,siRNA有效抑制FAS基因表達(dá)后,細(xì)胞內(nèi)及細(xì)胞外培養(yǎng)液中甘油三酯含量較對(duì)照組顯著降低,提示通過(guò)siRNA沉默F(xiàn)AS基因可能是一條潛在的降低血漿甘油三酯水平的有效途徑。
肝細(xì)胞從頭合成甘油三酯后,通過(guò)合成和分泌VLDL將其轉(zhuǎn)出肝臟。MTP存在于肝細(xì)胞內(nèi)質(zhì)網(wǎng)腔內(nèi),是VLDL組裝必需的異源二聚體酯轉(zhuǎn)運(yùn)蛋白。MTP的主要功能是將新合成的甘油三酯轉(zhuǎn)運(yùn)至載脂蛋白B,逐漸形成成熟的VLDL顆粒。VLDL顆粒不僅轉(zhuǎn)運(yùn)內(nèi)源性甘油三酯,肝合成的膽固醇也通過(guò)其轉(zhuǎn)出。當(dāng)VLDL-甘油三酯被脂蛋白酯酶(LPL)水解后,VLDL殘粒可轉(zhuǎn)變成低密度脂蛋白(LDL)。因此,MTP不僅在肝甘油三酯輸出中發(fā)揮作用,而且與肝膽固醇的分泌有關(guān)。研究顯示,MTP抑制劑可顯著降低血漿總膽固醇水平[20,21]。在本研究中,F(xiàn)AS基因siRNA沉默后,MTP基因表達(dá)顯著下調(diào),同時(shí)細(xì)胞外總膽固醇含量顯著降低,這可能是因?yàn)镕AS基因表達(dá)被抑制后,HepG2細(xì)胞分泌VLDL顆粒減少,從而導(dǎo)致膽固醇分泌也減少,培養(yǎng)液中總膽固醇降低。FAS基因沉默可能對(duì)MTP基因的表達(dá)形成正反饋,使MTP基因表達(dá)顯著降低。肝細(xì)胞內(nèi)甘油三酯
和膽固醇的合成是協(xié)調(diào)一致的,當(dāng)肝甘油三酯合成受到抑制時(shí),膽固醇的合成也會(huì)相應(yīng)降低[22],因此,盡管在本研究中HepG2細(xì)胞分泌的膽固醇減少,細(xì)胞內(nèi)膽固醇含量也沒(méi)有顯著增加。
HL存在于肝血竇內(nèi)皮細(xì)胞,其功能是水解VLDL殘粒中的甘油三酯,促進(jìn)VLDL殘骸的清除。在本研究中,F(xiàn)AS基因沉默后HL基因表達(dá)顯著增高,這提示FAS基因沉默后不僅可以減少肝臟VLDL的合成與輸出,還可促進(jìn)肝臟對(duì)外周脂蛋白殘骸的吸收和清除。本研究沒(méi)有發(fā)現(xiàn)siRNA沉默F(xiàn)AS基因后SREBP1基因表達(dá)發(fā)生顯著性變化,提示FAS基因沉默后MTP基因和HL基因表達(dá)的改變可能是通過(guò)其它信號(hào)通路實(shí)現(xiàn)的。
本研究在細(xì)胞水平上揭示了FAS基因siRNA沉默可降低肝細(xì)胞內(nèi)及細(xì)胞外培養(yǎng)液中的甘油三酯水平,也可降低細(xì)胞外培養(yǎng)液中的總膽固醇水平,需進(jìn)一步的體內(nèi)外實(shí)驗(yàn)加以驗(yàn)證。
[1] Sarwar N, Danesh J, Eiriksdottir G, et al. Triglycerides and the risk of coronary heart disease: 10, 158 incident cases among 262, 525 participants in 29 Western prospective studies. Circulation, 2007, 115: 450-458.
[2] 楊偉憲, 楊錚, 竇克非, 等. 217例青年冠心病患者臨床特點(diǎn)分析.中國(guó)循環(huán)雜志, 2014, 29: 339-342.
[3] Stauffer ME, Weisenfluh L, Morrison A. Association between triglycerides and cardiovascular events in primary populations: a meta-regression analysis and synthesis of evidence. Vasc Health Risk Manag, 2013, 9: 671-680.
[4] Staniak HL, Salgado Filho W, Miname MH, et al. Association between postprandial triglycerides and coronary artery disease detected by coronary computed tomography angiography. Atherosclerosis, 2014, 233: 381-386.
[5] Miller M, Cannon CP, Murphy SA, et al. Impact of triglyceride levels beyond low-density lipoprotein cholesterol after acute coronary syndrome in the PROVE IT-TIMI 22 trial. J Am Coll Cardiol, 2008, 51: 724-730.
[6] 劉玲, 趙水平, 王鐘林. 甘油三酯與冠心病關(guān)系及其機(jī)制研究. 中國(guó)循環(huán)雜志, 1998, 13: 251-252.
[7] Fukuda N, Ito T, Ohwada K, et al. Upregulation of fatty acid synthesis and the suppression of hepatic triglyceride lipase as a direct cause of hereditary postprandial hypertriglyceridemia in rabbits. J Clin Biochem Nutr, 2013, 53: 114-121.
[8] Ma N, Nicholson CJ, Wong M, et al. Fetal and neonatal exposure to nicotine leads to augmented hepatic and circulating triglycerides in adult male offspring due to increased expression of fatty acid synthase. Toxicol Appl Pharmacol, 2014, 275: 1-11.
[9] Burdeos GC, Nakagawa K, Kimura F, et al. Tocotrienol attenuates triglyceride accumulation in HepG2 cells and F344 rats. Lipids, 2012, 47: 471-481.
[10] Duncan RE, Archer MC. Farnesol decreases serum triglycerides in rats: identification of mechanisms including up-regulation of PPARalpha and down-regulation of fatty acid synthase in hepatocytes. Lipids, 2008, 43: 619-627.
[11] Loftus TM, Jaworsky DE, Frehywot GL, et al. Reduced food intake and body weight in mice treated with fatty acid synthase inhibitors. Science, 2000, 288: 2379-2381.
[12] Hopperton KE, Duncan RE, Bazinet RP, et al. Fatty acid synthase plays a role in cancer metabolism beyond providing fatty acids for phospholipid synthesis or sustaining elevations in glycolytic activity. Exp Cell Res, 2014, 320: 302-310.
[13] 趙勵(lì)彥, 王莉莉, 李松. 脂肪酸合成酶抑制劑減肥作用機(jī)制的研究進(jìn)展. 中國(guó)藥理學(xué)通報(bào), 2006, 22: 780-784.
[14] 冷亮, 蔣卓勤, 紀(jì)桂元. 大豆異黃酮對(duì)非酒精性脂肪肝大鼠肝臟脂代謝的影響. 中華預(yù)防醫(yī)學(xué)雜志, 2011, 45: 335-339.
[15] Soutschek J, Akinc A, Bramlage B, et al. Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature, 2004, 432: 173-178.
[16] Tadin-Strapps M, Peterson LB, Cumiskey AM, et al. siRNA-induced liver ApoB knockdown lowers serum LDL-cholesterol in a mouse model with human-like serum lipids. J Lipid Res, 2011, 52: 1084-1097.
[17] Koornneef A, Maczuga P, van Logtenstein R, et al. Apolipoprotein B knockdown by AAV-delivered shRNA lowers plasma cholesterol in mice. Mol Ther, 2011, 19: 731-740.
[18] Frank-Kamenetsky M, Grefhorst A, Anderson NN, et al. Therapeutic RNAi targeting PCSK9 acutely lowers plasma cholesterol in rodents and LDL cholesterol in nonhuman primates. Proc Natl Acad Sci USA, 2008, 105: 11915-11920.
[19] Frank-Kamenetsky M, Grefhorst A, Anderson NN, et al. Effect of an RNA interference drug on the synthesis of proprotein convertase subtilisin/kexin type9 (PCSK9) and the concentration of serum LDL cholesterol in healthy volunteers: a randomised, single-blind, placebocontrolled, phase 1 trial. Lancet, 2013, 6736: 61914-61915.
[20] Cuchel M, Meagher EA, du Toit Theron H, et al. Efficacy and safety of a microsomal triglyceride transfer protein inhibitor in patients with homozygous familial hypercholesterolaemia: a single-arm, open-label, phase 3 study. Lancet, 2013, 381: 40-46.
[21] Cuchel M, Bloedon LT, Szapary PO, et al. Inhibition of microsomal triglyceride transfer protein in familial hypercholesterolemia. N Engl J Med, 2007, 356: 148-156.
[22] Oliva J, French SW, Li J, et al. Proteasome inhibitor treatment reduced fatty acid, triacylglycerol and cholesterol synthesis. Exp Mol Pathol, 2012, 93: 26-34.
Effect of Small Interfering RNA Silencing the Fatty Acid Synthase Gene on Lipid Metabolism in Human Hepatic Cell Line HepG2
ZHANG Xiu-yun, FENG Yan-ping, XIONG Dong, YE Wen-jing, ZHU Li-ren, FAN A-qiang, CHEN Jian-ye, SONG Yong-yan.
Department of Biochemistry, School of Preclinical Medicine, North Sichuan Medical College, Nanchong (637000), Sichuan, China
Objective: To investigate the effect of the gene interfering technology on fatty acid synthase (FAS) gene silencing for lipid contents in human hepatic cell line HepG2 and to study the lipid metabolism related gene expression in HepG2 cells.
Small Interfering RNA; Fatty acid synthase; Human hepatic cell line HepG2
2014-11-24)
(編輯:汪碧蓉)
四川省應(yīng)用基礎(chǔ)研究計(jì)劃(2013JY0072);四川省教育廳重點(diǎn)項(xiàng)目(12ZA232)
637000 四川省南充市,川北醫(yī)學(xué)院基礎(chǔ)醫(yī)學(xué)院生物化學(xué)教研室(張秀云、陳建業(yè)、宋永硯);川北醫(yī)學(xué)院臨床醫(yī)學(xué)系(馮艷萍、熊東、葉文靜、朱粒仁、范阿強(qiáng))
張秀云 講師 碩士 主要從事脂代謝紊亂與動(dòng)脈粥樣硬化研究 Email: 396530604@qq.com 通訊作者:宋永硯 Email: songyongyan2014@hotmail.com
R54
A
1000-3614( 2015 )07-0670-05
10.3969/ j. issn. 1000-3614. 2015.07.014
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問(wèn)題研究(2012年1期)2012-03-25 09:54:45
中國(guó)火炬(2010年12期)2010-07-25 13:26:22
中國(guó)火炬(2010年8期)2010-07-25 11:34:30

- 中國(guó)循環(huán)雜志的其它文章
- 遠(yuǎn)端缺血后處理對(duì)兔心臟缺血/再灌注損傷的保護(hù)作用及其機(jī)制研究
- 丹參酮ⅡA對(duì)自發(fā)性高血壓大鼠左心室心肌肥厚及心肌細(xì)胞凋亡的影響
- 吡哆胺和替米沙坦對(duì)自發(fā)性高血壓大鼠心肌重塑相關(guān)指標(biāo)的影響
- 探討白藜三醇抑制快速電刺激乳鼠心肌細(xì)胞氧化應(yīng)激損傷及其機(jī)制
- 普羅帕酮對(duì)兔心室肌細(xì)胞動(dòng)作電位的影響及對(duì)快鈉電流的使用依賴(lài)性阻滯作用
- 主動(dòng)脈憩室的多種影像學(xué)診斷和臨床應(yīng)用價(jià)值的比較