NH4+對合成氣乙醇發酵過程的影響及其酶學機理*
2015-12-25 02:00:58張炎達王風芹彭一丁謝慧宋安東
食品與發酵工業 2015年12期
張炎達,王風芹,彭一丁,謝慧,宋安東
(河南農業大學生命科學學院,農業部農業微生物酶工程重點實驗室,河南鄭州,450002)
全球能源消耗預計到2030年將增加57%,化石燃料的大規模使用同時還帶來諸多的能源緊缺、環境壓力加劇和燃料價格大幅度上漲等問題[1],這就使得各國政府急切尋求一種可持續環保的再生綠色能源,其中生物燃料乙醇得到了世界廣泛關注[2]。合成氣乙醇發酵是目前公認的一種極具潛力和競爭力的生物燃料乙醇生產技術[3],該技術具有原料來源廣泛、生物催化特異性高、能耗低、耐毒性強和H2∶CO的無限制比率等優點[4]。合成氣是由氣化原材料,如農林業廢棄物和生活有機廢物等各種生物質材料、工業廢氣、次品煤和天然氣等[5-6]經氣化裝置在高溫下含有水蒸氣和氧的條件下裂解為富含H2、CO和CO2三種主要成分的混合氣體,另外還包括其他組分,如:NH3、CH4、NOx、N2、C2H2、C2H4、C6H6、C2H6、SO2和焦油等[7],但根據原料和氣化設備不同,合成氣的組分和比列略有差異[8-9]。
合成氣厭氧乙醇發酵微生物目前的種類及數量較少,且乙醇發酵產量較低,如C.ljungdahlii和C.autoethanogenum等[10],而國內還未有新菌的報道。Wood-Ljungdahl途徑是該類菌體的唯一代謝路徑,且氫化酶(H2ase,hydrogenase)、一氧化碳脫氫酶(CODH,carbon monoxide dehydrogenase)、乙醇脫氫酶(ADH,alcohol dehydrogenase)和乙酸激酶(ACK,acetate kinase)為其四大關鍵酶,而H2ase為還原力代謝的必需酶[3,7],H2ase活性的高低與 CO 利用效率的大小成正相關。在合成氣乙醇發酵中,合成氣雜質氣體多對發酵產生有害影響,如Asma Ahmed等人[11]發現,微量的焦油和NO能抑制發酵和促進菌體休眠。同時,NO抑制合成氣發酵菌C.carboxidivorans的 H2ase活性而阻礙 H2利用[12]。XU 等[13]還得出合成氣中的NH3為H2ase的非競爭抑制劑的結論。研究發現,雜質氣體NH3極易溶解于發酵培養基內,能快速轉化為NH4+且其濃度不斷累積[14],同時雜質NH3還會腐蝕設備,工業上常利用濕式洗滌技術進行脫氨處理[15],因而NH3對未來合成氣乙醇發酵工業化影響較大。相比,有關合成氣雜質氣體NH3對合成氣乙醇發酵過程的影響還未有報道,且NH3對發酵代謝關鍵酶的作用也未有研究,為此,本文系統研究了不同初始濃度的NH4+(NH4Cl)對菌株A-fm4,C.ljungdahlii和 C.autoethanogenum DSM10061利用生物質合成氣發酵乙醇的影響,并進一步測定菌株在菌體生長期和乙醇發酵期內4種關鍵酶的活性變化。以期研究NH3對合成氣乙醇發酵的影響和作用機理,為未來合成氣乙醇發酵工業化發展提供參考依據。另外,實驗中進行了菌株耐銨能力的研究,便于為日后耐銨菌株馴化提供菌種材料,希望獲得能夠利用含NH3粗合成氣組分發酵的優良菌株。
1 材料與方法
1.1 菌株
菌株 A-fm4富集自羊駝的新鮮糞便[16];菌株Closlridium ljungdahlii和Closlridium autoethanogenum DSM10061均購自德國微生物菌種保藏中心(DSMZ)。
1.2 生物質合成氣
實驗用合成氣按照玉米秸稈氣化氣組分配制,成分及比例如下:CO 85.5%,H210%,CO24.5%。購于河南源正科技發展有限公司。
1.3 培養基及配制方法
種子培養基(g/L)[17]:NaCl 0.80,NH4Cl 1.00,KCl 0.10,MgSO4·7H2O 0.20,CaCl20.04,KH2PO40.20,NaHCO31.00,酵母膏 1.00,MES[2-(N-嗎啡啉)乙磺酸]5.00,L-鹽酸半胱氨酸0.40,木糖5.00。添加微量元素溶液10 mL和維生素溶液10mL,pH 5.75。
發酵培養基(g/L)[17]:NH4Cl 1.00,NaCl 1.00,MgSO40.15,KH2PO40.10,CaCl20.04,胰蛋白胨2.00,酵母膏0.30,L-鹽酸半胱氨酸0.20,MES 10.00,礦質元素溶液10 mL,維生素溶液10 mL,pH 7.0。
實驗中發酵培養基初始NH4+濃度設置了6個梯度,分別為 0X、0.5X、1X、2X、4X 和 8X,因雜質氣體NH3極易溶解于發酵培養基內,且能夠快速轉化為NH4+使得 NH4+濃度不斷累積[14],所以實驗中利用NH4+的濃度進行研究,即,將原發酵培養基的NH4Cl濃度標記為1 X。
維生素溶液均采用0.22 μm的濾膜過濾除菌,待接種后添加至培養基內。
上述所有培養基均準確配制,每80 mL培養液分裝至200 mL厭氧培養瓶內(發酵培養基為每60 mL培養液分裝至300 mL厭氧培養瓶內),再轉入厭氧培養箱內添加刃天青0.5 mg/L,靜置直至瓶內顏色由粉色轉為無色時分別蓋上瓶塞,于121℃滅菌20 min(滅菌過程中瓶塞插入針頭)。滅菌后立刻拔去針頭且移至厭氧培養箱內,待溫度降至約37℃時進行后續實驗。
1.4 合成氣厭氧乙醇發酵
菌種活化參考文獻[16],將對數期的各菌種子液按10%接種量轉接至含有60 mL發酵培養液的厭氧培養瓶中,用注射器向厭氧瓶內加入240 mL合成氣(以上操作均在厭氧培養箱內完成)。37℃,150 r/min搖床發酵7 d,每24 h取樣測OD600值及乙醇含量。
1.5 產物測定及計算方法
1.5.1 乙醇和乙酸含量測定
將1.5 mL發酵液于4℃,8 000 r/min離心10 min,上清液用于兩者濃度的測量。測定條件為:FID檢測器,柱子為30 m ×0.32 mm ×0.3 μm HP-FFAP,氮氣作為載氣,起始3 min流速為1.9 mL/min,再以0.5 mL/min 升至 4 mL/min,分流比 50∶1,柱箱起始溫度40℃,保持1.5 min后以40℃/min達到210℃并保持2 min,色譜進樣口和檢測器的溫度分別為200℃和250℃。
1.5.2 細胞干重和指標計算
根據OD值及其對應的細胞干重(DCW)可獲得發酵菌株的干重(y,g/L)與OD值(x)關系回歸方程式。
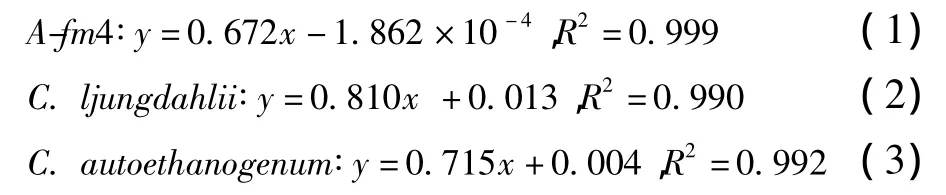
1.6 關鍵酶活性測定
關鍵酶H2ase、CODH和ADH活性測定方法分別參考文獻[18];關鍵酶ACK活性測定方法參考文獻[19]和[20],稍有改進。反應為:H2ase活性反應體系具體包含:0.4 mL 1 mol/L Tris-HCl(pH=7.5),0.2 mL 0.04 mol/L甲基紫精,0.2 mL 0.04 mol/L二硫蘇糖醇,0.1 mL體積分數為5%的Triton-X100,2 mL脫氣去離子水;CODH活性反應體系具體包含:0.8 mL 0.5 mol/L Tris-HCl(pH=6.8),0.2 mL 0.04 mol/L甲基紫精(MV),0.2 mL 0.04 mol/L二硫蘇糖醇,0.1 mL體積分數為5%的Triton-X100,1.2 mL脫氣去離子水。ADH活性反應體系具體包含:0.4 mL 1 mol/L Tris-Hcl(pH=7.5),0.12 mL 0.01 M NADH,0.4 mL 0.1 M 乙醛,0.5 mL 0.08 mol/L 二硫蘇糖醇,0.1 mL體積分數為5%的Triton-X100,1.5 mL脫氣去離子水;ACK活性反應體系具體包含:0.145 mL 1 mol/L Tris-HCl(pH=7.5),0.1 mL 2 mol/L乙酸鉀,0.25 mL 0.08 mol/L二硫蘇糖醇,0.1 mL體積分數為5%的Triton-X100,0.1 mL 0.1 mol/L MgCl2,0.1 mL 0.1 mol/L ATP,0.2 mL 3.5 mol/L 中性NH4OH-HCl,2 mL脫氣去離子水。且二硫蘇糖醇溶液現配現用。
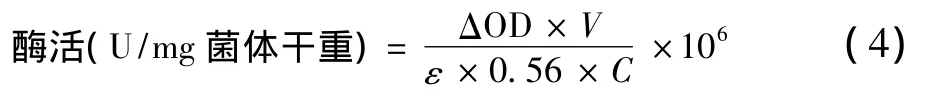
式中:△OD為吸收值每分鐘降低的變化值,min-1;V為反應體系的體積,mL;E為摩爾消光系數,103L/(mol·cm);0.56為96孔板的光程,cm;C為測定中所加入菌體干物質量,g;106為摩爾轉化為微摩爾的系數。酶活單位:單位干菌體每分鐘還原1 μmol MV(1e-1還原)。
1.7 NH4+對菌株生長半抑制濃度
NH4+對發酵菌株生長半抑制濃度(Ki)的計算參考文獻[21],通過對比不同發酵菌株的Ki值大小來衡量菌株的NH4+耐受能力強弱。
1.8 統計分析方法
為比較在不同初始NH4+濃度下對菌株發酵乙醇產量及其各發酵參數的影響,同時分析比較對兩時期4種關鍵酶活性水平的差異,實驗分別采用了方差分析和t值檢驗,以上各統計分析均由軟件SPSS16.0完成。
2 結果與討論
2.1 初始N H4+濃度對合成氣發酵菌株生長的影響
不同梯度濃度NH4+對發酵菌株的生長影響見圖1。菌株 A-fm4、C.ljungdahlii和 C.autoethanogenum的生物量分別在NH4+濃度0.5X、0X和0X于發酵4 d、1 d和2 d達到最大,菌體量分別為:0.045,0.166和0.097 g/L。菌株 A-fm4和 C.autoethanogenum DSM10061的生長過程受NH4+影響明顯,NH4+對菌株C.ljungdahlii的影響主要表現在生長穩定期及以后,且隨NH4+梯度濃度增大對菌株生長量和生長速率具有抑制能力。
由此可知,合成氣發酵菌株對無機氮源的利用有限,發酵液內NH4+累積濃度高于9.35 mmol/L時生長受到不同程度的抑制作用。據報道,Escherichia coli已作為一種模式生物進行研究細菌對NH4+的吸收和排除是經過一種高K+親和性的K+-NH4+交換系統(Kdp)[22],其中 Buurman 等[23]對 E.coli在高濃度NH4+環境中代謝活動的研究發現在高濃度NH4+條件下需要一種額外轉運系統,即Kdp,來負責NH4+的攝入和泵出維持胞內外NH4+正常梯度及胞內pH的穩定,且高濃度NH4+使得菌體對能量的利用效率顯著降低。同樣,葉貴子等[21]對產琥珀酸放線桿菌的研究也得到相似結果。雖然目前還未見針對合成氣厭氧發酵菌在Kdp系統與能量效率方面的確切報道,但可推測在本實驗發酵菌體細胞內也存在類似機制。另外,數據顯示K+的水合半徑為5.32,近于NH4+的5.37,NH4+對K+在細胞膜同一載體上可競爭同一結合部位,這一競爭會破壞K+對細胞正常的生理功能,且隨NH4+濃度的增大不斷顯著,進而導致對菌體的生長抑制。
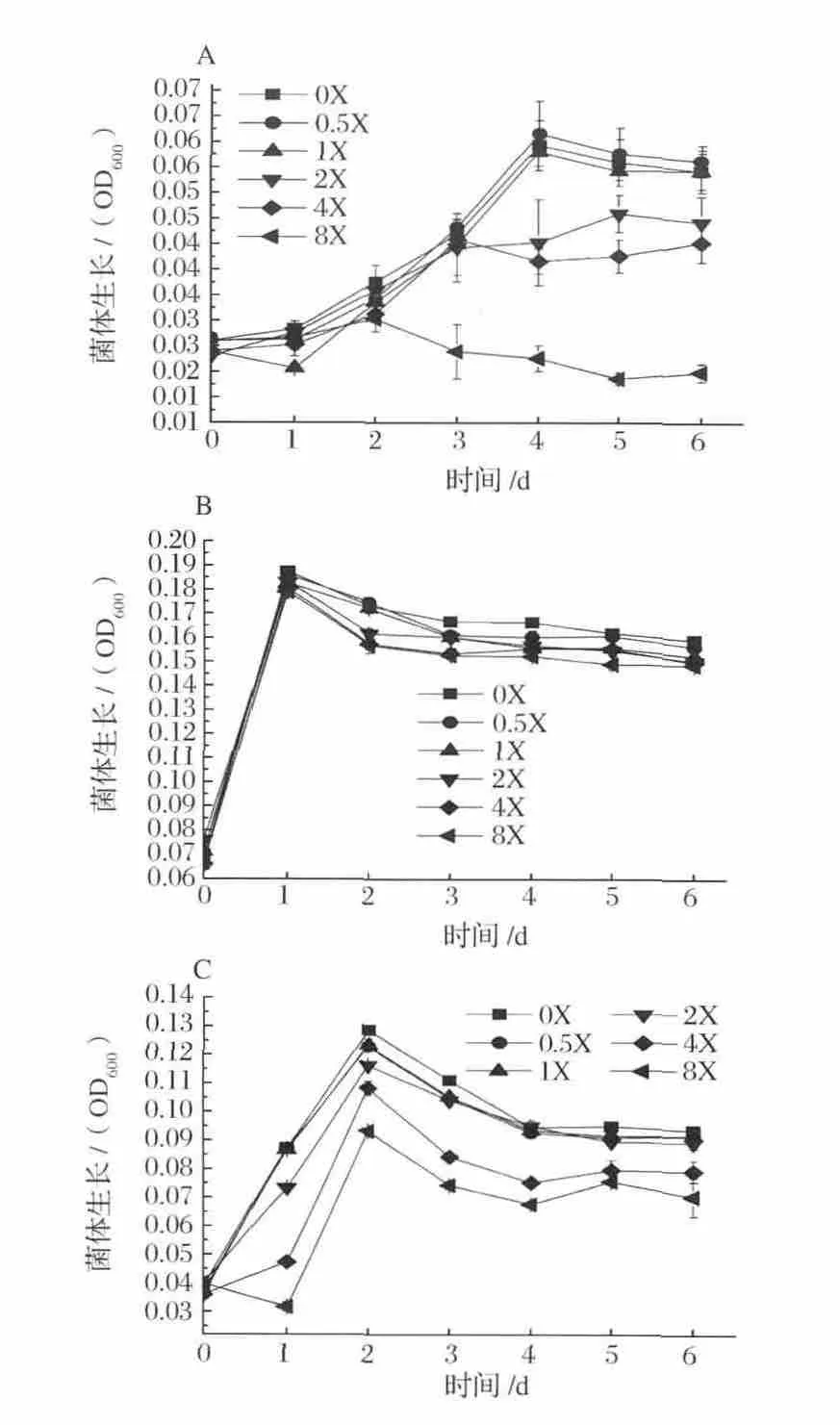
圖1 不同梯度NH4+濃度下發酵菌生長曲線Fig.1 Growth curve of organisms at different concentrations of NH4+
2.2 初始NH4+濃度對合成氣發酵乙醇特性的影響
菌株 A-fm4、C.autoethanogenum DSM10061和C.ljungdahlii的乙醇產量均在NH4+濃度0X下于發酵5 d、2 d和3 d達到最大,其乙醇量分別為124.56,137.71和137.67 mg/L(圖2)。其中,菌群A-fm4的乙醇發酵受NH4+濃度影響最大,其次為菌株C.autoethanogenum DSM10061,再次是菌株 C.ljungdahlii。NH4+濃度高于0.5X(9.35 mmol/L)時開始減緩菌株乙醇發酵速率,當濃度高于4X(74.8 mmol/L)其抑制作用更顯著,因此不存在或低濃度NH4+會促進乙醇發酵,高濃度NH4+能降低乙醇發酵能力。
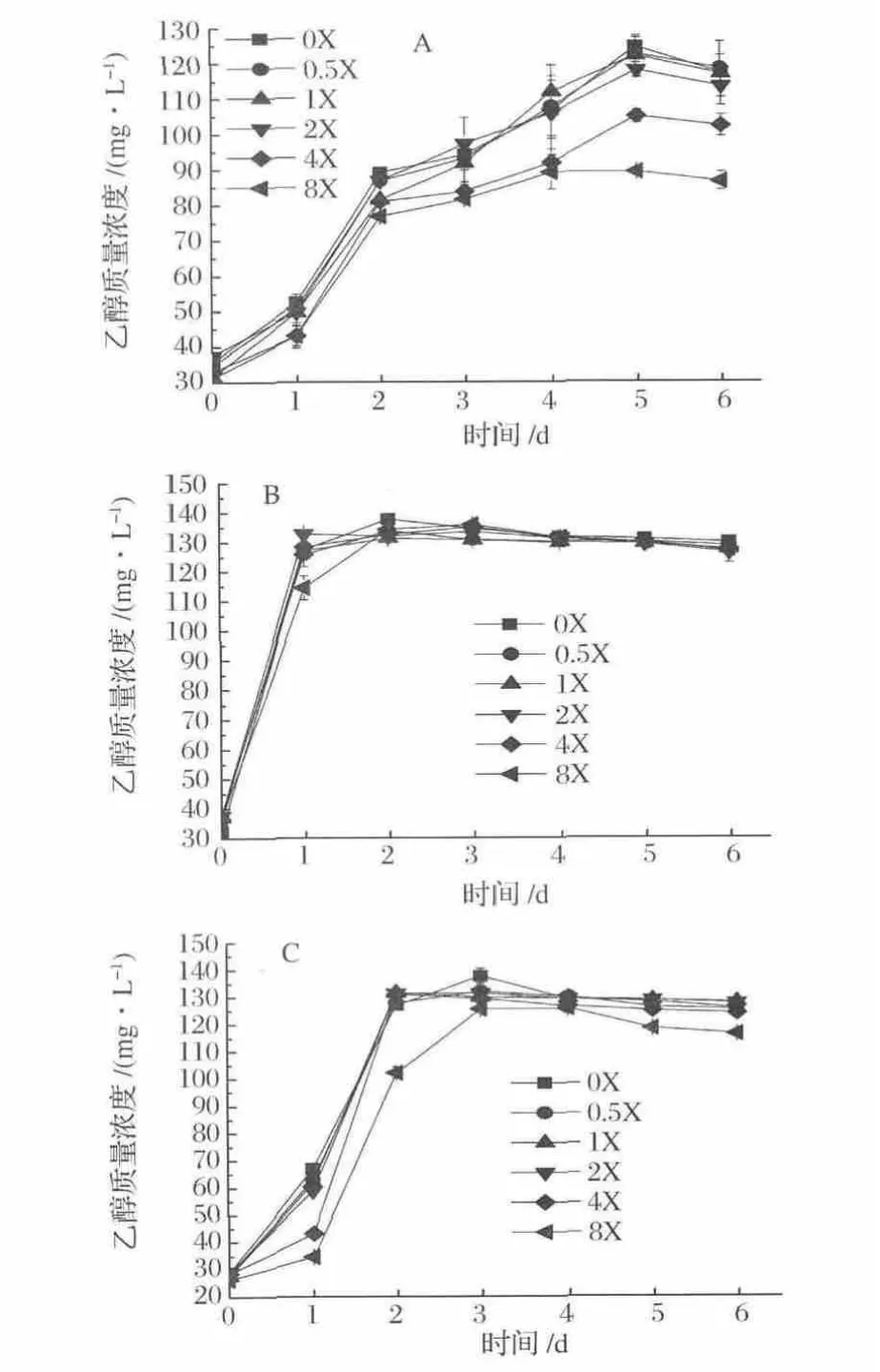
圖2 不同梯度NH4+濃度下乙醇產量Fig.2 Ethanol production fermented at different concentrations of NH4+
2.3 初始N H4+濃度對合成氣發酵乙酸特性的影響
圖3為不同梯度濃度NH4+對合成氣發酵菌株發酵乙酸特性的影響。菌株A-fm4、C.autoethanogenum DSM10061和C.ljungdahlii的乙酸產量分別在NH4+濃度2X、2X和0X或2X下于發酵5 d、4 d和3 d或4 d達到最大,乙酸量分別為218.19,302.72和304.80或304.95 mg/L。相比菌株A-fm4和C.autoethanogenum,菌株C.ljungdahlii乙酸發酵受 NH4+影響較弱,另外,NH4+濃度0.5X~4X(9.35~74.8 mmol/L)能不同程度地促進菌株A-fm4、C.autoethanogenum DSM10061和C.ljungdahlii乙酸發酵速率。結果表明,NH4Cl濃度在18.7~74.8 mmol/L會促進乙酸的發酵,濃度為149.6 mmol/L抑制乙酸發酵。
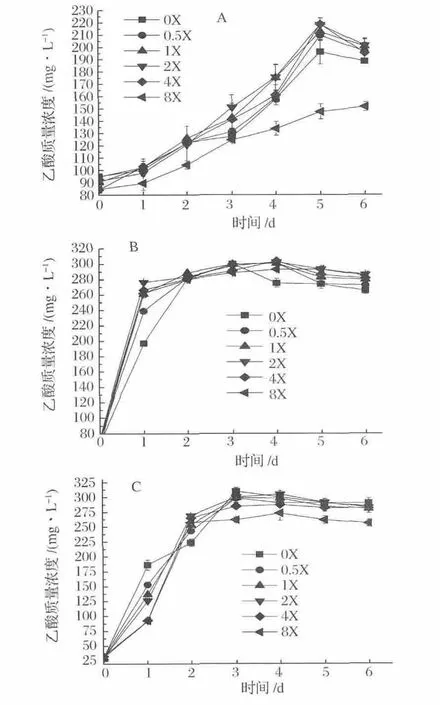
圖3 不同梯度NH4+濃度下乙酸產量Fig.3 Acetate production fermented at different concentrations of NH4+
綜合分析發酵菌株在不同梯度NH4+環境內的發酵情況可得:低濃度NH4+利于乙醇和乙酸發酵,高濃度NH4+對乙醇和乙酸均表現出抑制作用。當NH4+濃度高于9.35 mmol/L時,乙醇發酵隨濃度的增大而抑制顯著,NH4+濃度為18.7~74.8 mmol/L時會促進乙酸的發酵。發酵培養液內不含NH4+時利于更多的代謝流流向乙醇,促進乙醇且減弱乙酸發酵。
2.4 初始NH4+濃度對合成氣發酵代謝關鍵酶活性的影響
不同初始NH4+濃度對合成氣厭氧發酵菌株發酵代謝途徑關鍵酶活性變化的影響結果見圖4。H2ase和CODH酶活水平隨NH4+濃度增大而抑制極顯著(P<0.01),生長期酶活水平高于發酵期,NH4+濃度低于2X(37.4 mmol/L)促進菌株A-fm4該酶的高水平維持(A1~C1,A2~C2);除菌株 C.ljungdahlii外(B3),NH4+濃度大于2X(37.4 mmol/L)時ADH酶活大小受到抑制作用,其中菌株A-fm4生長期酶活水平大于發酵期,兩時期酶活均受NH4+影響顯著(A3),菌株C.autoethanogenum DSM10061生長期酶活水平受NH4+影響,而發酵期 ADH酶活大小趨于穩定(C3)。相比,菌株C.ljungdahlii兩時期ADH酶活均受NH4+影響較小;對于ACK,菌株A-fm4和C.ljungdahlii在低濃度和高濃度NH4+下其活性水平均受抑制,NH4+濃度1X~2X(18.7 mmol/L~37.4 mmol/L)可促進ACK活性,且生長期酶活水平一般高于發酵期(A4和B4),而菌株C.autoethanogenum DSM10061在NH4+濃度低于2X(37.4 mmol/L)時可維持較高活性水平,高濃度NH4+抑制酶活(C4)。因此,低濃度NH4+可促進關鍵酶活性,高濃度NH4+具有抑制作用,且生長期酶活水平較大于發酵期的值。
Wood-Ljungdahl代謝途徑中H2ase和CODH可分別通過催化氧化H2和CO產生還原力和質子,其中H2ase為代謝過程中還原力獲得的必需酶,H2ase活性水平會直接影響到 CO的利用效率,高活性H2ase可弱化由CO經CODH產生還原力的途徑,促進CO作為碳源和能源,而H2ase低活性時其結果相反[1]。由上述結果可知,NH4+濃度能影響 H2ase和CODH的活性大小,而對H2ase的活性影響顯著,因此從酶學層面上可解釋NH4+濃度大于2X時逐漸抑制發酵菌株的生長,而低濃度或不存在NH4+可促進菌體的生長(圖1)。另外,NH4+對H2ase和CODH的抑制機制可理解如下,即,NH3和NH4+在發酵液中快速轉化,NH3易通過細胞膜進入胞內,繼續電離NH4+[24],改變胞漿內外的原有正常NH4+梯度,使得H2ase和CODH暴露于更多NH4+環境內,自由NH4+的增多影響到酶結構域的電位變化導致其活性降低,NH4+濃度越高抑制酶活作用越強。研究證明,H2ase和CODH常依附于胞膜內側[25],鑒于高濃度NH4+會對其抑制及NH3易出入細胞膜的能力,可假設在菌株發酵中存在于菌體胞內的H2ase和CODH受NH4+抑制濃度可能為37.4 mmol/L。ADH是 Wood-Ljungdahl中乙醇代謝的關鍵酶,實驗中菌株 A-fm4的ADH酶活受NH4+濃度影響顯著,當NH4+濃度在2X以內時該酶活性維持較高水平(圖4-A3),進而促進乙醇發酵(圖2-A)。對于該菌群,理論上ADH酶活應在發酵期達到較高水平,實際上發酵期獲得最大乙醇產量但其活性降低,可能的解釋是ADH活性雖在后期降低,但足以滿足合成乙醇所必須的酶活大小,或是在發酵期ADH隨著發酵環境的改變ADH活性降低,但在菌體內的表達量卻在增加來滿足乙醇代謝[18]。針對菌株 C.ljungdahlii和 C.autoethanogenum DSM10061,ADH的活性受NH4+影響小,酶活水平相當,具有一定的耐受能力,有學者[26]對菌體C.ljungdahlli內參基因穩定性及重要基因表達量的研究發現,與乙醇合成相關的重要基因aor2在不同碳源內的表達量穩定,這可提示本實驗兩菌株在不同濃度NH4+發酵環境中該基因或ADH相關重要基因也存在類似穩定表達的能力進而使得ADH活性水平穩定,只是對于C.autoethanogenum DSM10061這種穩定表達在乙醇發酵期得到主要體現。另外,菌株C.ljungdahlii和C.autoethanogenum DSM10061在乙醇發酵期的ADH雖具有一定的耐受性(圖4-B3,B4),但乙醇產量卻低于低NH4+濃度(小于2X)的發酵乙醇產量,關鍵問題是在較高NH4+濃度下H2ase活性受到抑制,進而在代謝流中用于乙醇代謝的還原力減少導致乙醇合成降低。
合成氣厭氧發酵的乙酸發酵離不開關鍵酶ACK,實驗發現,NH4+濃度在18.7~37.4 mmol/L可促進其活性,相繼乙酸產量增加(圖3)。室溫下,乙酸解離常數pKa為4.75,在菌體胞漿的偏中性中其主要以 Ac-和 H+存在,因此在1X和2X下胞內NH4+的電離利于胞漿正常pH范圍的穩定,減少大量乙酸在胞內對酶促反應的抑制,隨分子乙酸由細胞調節機制[27]不斷移出胞內使發酵液內乙酸濃度不斷增加。基于0X下促進乙醇而降低乙酸發酵,因此當不存在NH4+時利于更多的代謝流流向乙醇。
2.5 發酵菌株對NH4+的耐受能力對比
由表1知,NH4+對不同種類發酵菌的生長抑制程度不同,3種發酵出發菌中僅菌株A-fm4對NH4+最為敏感,其 Ki值為 123.61 mmol/L。相比,菌株 C.ljungdahlii和 C.autoethanogenum DSM10061的 Ki值分別為208.13 mmol/L和240.37 mmol/L,耐NH4+能力較強,這或許為未來該菌直接利用含氨氣的粗制合成氣發酵乙醇提供重要啟示。同時菌株C.ljungdahlii和C.autoethanogenum DSM10061可為未來工業菌株耐銨馴化提供菌種材料,耐銨菌種的使用利于合成氣厭氧乙醇發酵的工業化進程,為降低粗制合成氣脫氨設備的資金投入甚至刪減脫氨工藝工序都具有重要意義。
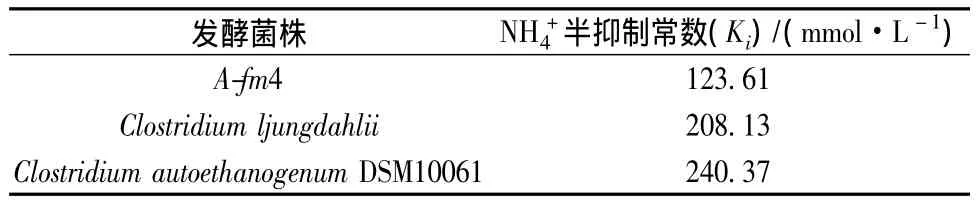
表1 NH4+對發酵菌株的半抑制常數(Ki)Table 1 Ammnia half-inhibitin constant(Ki)for fermentation microorganisms
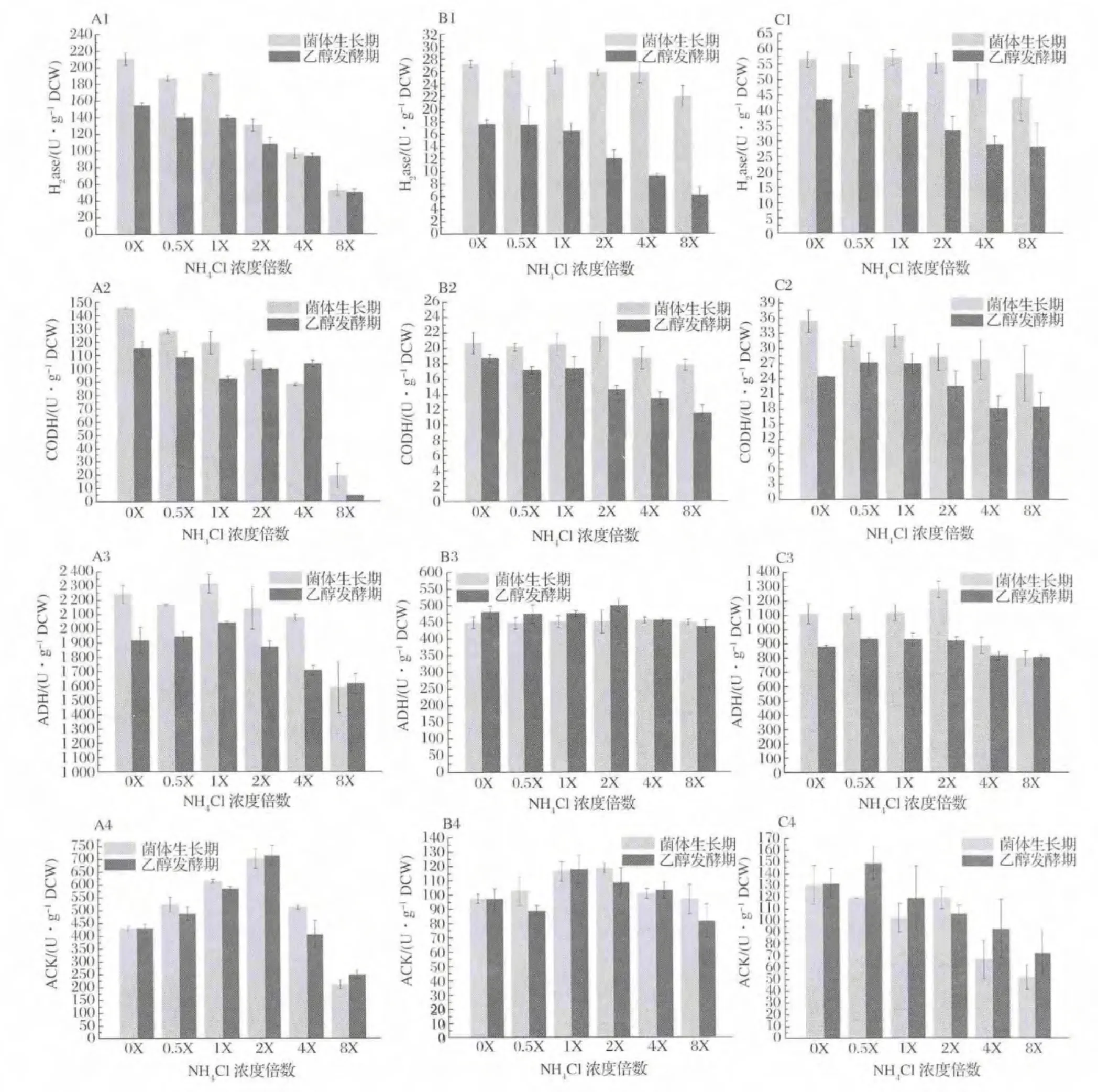
圖4 NH4+對菌株發酵過程中兩時期關鍵酶活性的影響Fig.4 The activity of key enzymes existed in growth period and ethanol fermentation period at different concentrations of NH4+
3 結論
(1)發酵培養基內不同初始濃度的NH4+對菌株A-fm4,C.ljungdahlii和 C.autoethanogenum DSM10061發酵過程中的生長影響顯著,其中對菌株C.ljungdahlii的影響主要表現在生長穩定期及以后。結果表明,菌株 A-fm4、C.ljungdahlii和 C.autoethanogenum DSM10061分別在NH4+濃度0.5X、0X和0X下獲得最大生物量,分別為0.045,0.166和0.097 g/L。當NH4+濃度高于9.35 mmol/L時,隨NH4+梯度濃度增大對菌株生長量和生長速率具有抑制能作用。
(2)綜合分析合成氣發酵過程可知:低濃度NH4+利于乙醇和乙酸發酵,高濃度NH4+對乙醇和乙酸均表現出抑制作用。當NH4+濃度高于9.35 mmol/L時,乙醇發酵隨濃度的增大而抑制顯著,NH4+濃度為18.7~74.8 mmol/L時會促進乙酸的發酵。發酵培養液內不含NH4+時利于更多的代謝流流向乙醇,促進乙醇且減弱乙酸發酵。
(3)關鍵酶 H2ase,CODH,ADH和 ACK均受高濃度(149.6 mmol/L)NH4+抑制,且菌體生長期的酶活水平一般高于發酵期,H2ase和CODH隨NH4+濃度升高而酶活下降極顯著,H2ase活性的高低顯著影響發酵過程中還原力的供給,直接影響菌體生長和乙醇發酵。除菌株A-fm4外,菌株C.ljungdahlii和C.autoethanogenum DSM10061的ADH受NH4+影響較小,推測該酶在胞內具有穩定表達機制。NH4+濃度1X~2X(18.7~37.4 mmol/L)可促進ACK活性,且不存在NH4+時也會降低該酶活性。
(4)菌株 C.ljungdahlii和 C.autoethanogenum DSM10061的Ki值分別為208.13 mmol/L和240.37 mmol/L,耐銨能力較強,可為未來耐銨菌種的馴化提供菌種材料,為將來降低脫氨設備資金投入和減少合成氣凈化的復雜工序提供技術準備。
[1] Latif H,Zeidan A A,Nielsen A T,et al.Trash to treasure:production of biofuels and commodity chemicals via syngas fermenting microorganisms[J].Current Opinion in Biotechnology,2014,27:79-87.
[2] Vohra M,Manwar J,Manmode R,et al.Bioethanol production:Feedstock and current technologies[J].Journal of Environmental Chemical Engineering,2014,2(1):573-584.
[3] Bengelsdorf F R,Straub M,Dürre P.Bacterial synthesis gas(syngas)fermentation[J].Environmental Technology,2013,34(13):1 639-1 651.
[4] Henstra A M,Sipma J,Rinzema A,et al.Microbiology of synthesis gas fermentation for biofuel production[J].Current Opinion in Biotechnology,2007,18(3):200-206.
[5] Kirkels A F,Verbong G P.Biomass gasification:Still promising?A 30-year global overview[J].Renewable and Sustainable Energy Reviews,2011,15(1):471-481.
[6] Phillips J,Klasson K,Clausen E,et al.Biological production of ethanol from coal synthesis gas[J].Applied Biochemistry and Biotechnology,1993,39(1):559-571.
[7] Abubackar H N,Veiga M C,Kennes C.Biological conversion of carbon monoxide:rich syngas or waste gases to bioethanol[J].Biofuels,Bioproducts and Biorefining,2011,5(1):93-114.
[8] Somerville C,Youngs H,Taylor C,et al.Feedstocks for lignocellulosic biofuels[J].Science(Washington),2010,790(329):790-792.
[9] Rollins M L,Reardon L,Nichols D,et al.National Energy Technology Lab.,Pittsburgh,PA(US);National Energy Technology Lab.,Morgantown,WV(US),2002.
[10] Cotter J L,Chinn M S,Grunden A M.Influence of process parameters on growth of Clostridium ljungdahlii and Clostridium autoethanogenum on synthesis gas[J].Enzyme and Microbial Technology,2009,44(5):281-288.
[11] Ahmed A,Cateni B G,Huhnke R L,et al.Effects of biomass-generated producer gas constituents on cell growth,product distribution and hydrogenase activity of Clostridium carboxidivorans P7T[J].Biomass and Bioenergy,2006,30(7):665-672.
[12] Ahmed A,Lewis R S.Fermentation of biomass-generated synthesis gas:Effects of nitric oxide[J].Biotechnology and Bioengineering,2007,97(5):1 080-1 086.
[13] XU D,Lewis R S.Syngas fermentation to biofuels:Effects of ammonia impurity in raw syngas on hydrogenase activity[J].Biomass and Bioenergy,2012,45:303-310.
[14] CHEN S,LING J,Blancheton J-P.Nitrification kinetics of biofilm as affected by water quality factors[J].Aquacultural Engineering,2006,34(3):179-197.
[15] Daniell J,K?pke M,Simpson S D.Commercial biomass syngas fermentation[J].Energies,2012,12(5):5 372-5 417.
[16] 宋安東,馮新軍,王風芹,等.合成氣發酵制取乙醇微生物的對比[J].生物工程學報,2013,29(3):242-249.
[17] 王風芹,張炎達,謝慧,等.生物質氣化氣發酵生產乙醇優良菌株的篩選[J].農業工程學報,2015,31(7):221-226.
[18] Ukpong M N,Atiyeh H K,De Lorme M J,et al.Physiological response of Clostridium carboxidivorans during conversion of synthesis gas to solvents in a gas‐fed bioreactor[J].Biotechnology and Bioengineering,2012,109(11):2 720-2 728.
[19] Aceti D J,Ferry J G.Purification and characterization of acetate kinase from acetate-grown Methanosarcina thermophila.Evidence for regulation of synthesis[J].Journal of Biological Chemistry,1988,263(30):15 444-15 448.
[20] Sridhar J Eiteman M A,Wiegel J W.Elucidation of enzymes in fermentation pathways used by Clostridium thermosuccinogenes growing on inulin[J].Applied and Environmental Microbiology,2000,66(1):246-251.
[21] 葉貴子,姜岷,陳可泉,等.耐銨型產琥珀酸放線桿菌的選育及銨離子對其生長代謝的影響[J].生物工程學報,2010,26(2):183-188.
[22] Marinus M,Neidhardt F,Curtiss I,et al.Escherichia coli and Salmonella:cellular and molecular biology[J].Escherichia coli and Salmonella:Cellular and Molecular Biology,1996,2:1 579-1 601.
[23] Buurman E T,De Mattos M J T,Neijssel O M.Futile cycling of ammonium ions via the high affinity potassium uptake system(Kdp)of Escherichia coli[J].Archives of Microbiology,1991,155(4):391-395.
[24] Khademi S,O'connell J,Remis J,et al.Mechanism of ammonia transport by Amt/MEP/Rh:structure of AmtB at 1.35 ?[J].Science,2004,305(5 690):1 587-1 594.
[25] Bagyinka C,Kovács K L,Rak E.Localization of hydrogenase in Thiocapsa roseopersicina photosynthetic membrane[J].Biochemical Journal,1982,202(1):255-258.
[26] 劉娟娟.Clostridium ljungdahlii內參基因穩定性評價及其在重要基因相對表達量研究中的應用[D].重慶:西南大學,2013:41-50.
[27] Russell J.Another explanation for the toxicity of fermentation acids at low pH:anion accumulation versus uncoupling[J].Journal of Applied Bacteriology,1992,73(5):363-370.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
