葛根素影響雌激素受體α啟動子甲基化調控成骨細胞增殖分化的研究
2015-12-28 07:55:19呂海宏湯旭磊傅松波
河北醫科大學學報 2015年4期
呂海宏,湯旭磊,傅松波
(蘭州大學第一醫院內分泌科,甘肅 蘭州 730000)
·論著·
葛根素影響雌激素受體α啟動子甲基化調控成骨細胞增殖分化的研究
呂海宏,湯旭磊,傅松波
(蘭州大學第一醫院內分泌科,甘肅 蘭州 730000)
[摘要]目的探究葛根素對大鼠成骨細胞代謝調控的途徑,為絕經后骨質疏松治療提供理論依據。方法取新生SD大鼠的顱骨,膠原酶法培養成骨細胞,分別加入不同濃度(5、10、25、50、100 μmol/L)葛根素分組培養,應用巢式甲基敏感性聚合酶鏈反應方法觀察雌激素受體α(estrogen receptor α,ERα)啟動子A區甲基化,反轉錄-聚合酶鏈反應方法觀察ERα mRNA表達,MTT法和對硝基苯磷酸酯方法觀察體外培養成骨細胞的增殖及堿性磷酸酶活性。結果隨著葛根素濃度增加,ERα 基因啟動子甲基化程度逐漸減弱;同時發現細胞中ERα mRNA表達逐漸增強,25 μmol/L葛根素濃度時表達最強(P<0.01)。葛根素能刺激成骨細胞增殖,提高堿性磷酸酶活性(P<0.01)。結論葛根素抑制ERα啟動子甲基化及增加ERα mRNA表達,可能通過此途徑促進成骨細胞增殖分化。
[關鍵詞]成骨細胞;葛根素;DNA甲基化;雌激素受體α
doi:10.3969/j.issn.1007-3205.2015.04.005
絕經后骨質疏松是臨床常見的流行病,具有很高的患病率和骨折導致的致殘率。研究認為骨質疏松的發病機制主要是由于骨重建失衡導致此病發生,絕經后婦女骨吸收能力明顯大于骨形成,造成骨重建失衡,因此主導骨重建的成骨細胞和破骨細胞的調控受到更多重視。體內外骨質疏松模型相關研究表明雌激素受體α(estrogen receptor α,ERα)與成骨細胞的增殖分化有關[1]。葛根素是結構及生物活性均類似于雌激素的非甾體類化合物,為雜環多酚類化合物,可以與雌激素受體結合,具有弱雌激素活性,但葛根素對成骨細胞代謝調控的具體機制目前并不十分清楚。本研究旨在探討葛根素與ERα甲基化的關系以及對成骨細胞增殖分化的調控,報告如下。
1資料與方法
1.1原代成骨細胞培養及處理取新生(<24 h)SD大鼠,清洗,乙醇浸泡10 min,取顱蓋骨,PBS沖洗3遍,剪碎頭蓋骨。移入10 mL0.25%胰蛋白酶溶液,5 mLⅡ型膠原酶溶液,依次振蕩消化,棄上清液。沉淀細胞團塊用DMEM/F12液沖洗并懸浮于培養液中。細胞懸液混勻以1×106/mL密度接種于50 mL培養瓶培養、換液,傳代培養備用。實驗前PBS沖洗,接種細胞于培養板上,然后加入無血清的DMEM/F12,分別加入0、5、10、25、50、100 μmol/L不同濃度葛根素分組培養。
1.2實驗儀器及試劑CO2恒溫培養箱和低溫離心機(德國Heraeus);SPECORD 40紫外分光光度儀(英國analyticjena公司);倒置相差顯微鏡(日本OLYMPUS公司);PCR儀(Eppendorf公司生產);紫外凝膠成像儀(美國Thermo Forma公司生產)。Puerarin、Biozol及TRIZOL試劑(Sigma-Aldrich);ZR Genomic DNA 成套試劑盒、DNA 片段恢復成套試劑盒及RT-PCR試劑盒(Tiangen Biotech Company);PNPP、亞硫酸氫鈉、Triton X-100、Tris-HCl、1%茜素紅及2%堿性磷酸酶等試劑(Sino-American Biotechnology Company)。
1.3方法
1.3.1培養細胞染色鑒定①茜素紅染色法: PBS沖洗成骨細胞2遍,然后用95%酒精固定10 min,1%茜素紅(pH 4.2)染色,PBS沖洗觀察。②堿性磷酸酶染色法: PBS沖洗成骨細胞2遍,95%酒精固定10 min,2%堿性磷酸酶(pH 8.2)染色,PBS沖洗觀察。
1.3.2DNA提取培養傳代細胞中加入不同濃度葛根素進行培養,按照DNA提取試劑盒說明書常規操作進行成骨細胞基因組DNA的提取。
1.3.3亞硫酸鹽修飾和DNA恢復①將DNA濃度稀釋為20 μg/μL,取10 μL DNA加入50 mL雙脫水,5.5 mL3 mmol/L氫氧化鈉,混勻后孵育42 ℃30 min,使DNA充分溶解變性。②每個試管分別依次加入對苯二酚、3 mmol/L亞硫酸氫鈉(NaHSO3)(pH 5.0),混勻,低溫離心,按照說明書指示將經過上述處理的DNA脫鹽,異丙醇沖洗樣品,加入50 mL37 ℃熱水恒溫箱進行溫育,低速離心1 min。③再將10 mol/L NaAc(pH 5.2)和無水乙醇加入試管混勻,離心,棄溶液,留白色沉淀。DNA沉淀用-20 ℃預冷的70%酒精反復沖洗后過夜,真空干燥,用20 mL水溶解沉淀。洗脫的DNA樣品立即儲存在-20 ℃備用。
1.3.4體外培養細胞ERα啟動子甲基化測定①巢式甲基化特異性PCR(nested methylation-specific polymerase chain reaction,nMS-PCR)引物設計:根據Genebank中查詢到的登錄號為AY425004的ERα基因DNA序列,設計nMS-PCR的內外引物。nMS-PCR法包括兩步PCR反應,第一步PCR使用的是不含任何CG序列的外側引物,第二步PCR使用的內側引物,其中ERα的內側引物位于ERα基因序列的啟動子區,共包含有8個CG序列,引物由上海生物工程公司合成,見表1。②nMS-PCR方法:應用Touchdown(TD) PCR方法擴增,按照試劑盒第一輪25 μL PCR反應體系加入試劑,ER-pN 20 μmol/L 0.3 μL,DNA 2 μL,2×Taq PCR Mastermix 6.3 μL,15 mmol/L的Tris-HCl(pH 8.0),25 mmol/L KCl,2.5 mmol/L MgCl2,250 μmol/L 4種dNTP,1 μL上、下游引物,3 μL DNA模板,加入25 μL蒸餾水。第二輪PCR反應DNA模板應用第一輪PCR擴增產物。隨著熱啟動,樣品應用Touchdown(TD) PCR方法進行20輪的擴增。

表1 ERα 的nMS-PCR引物Table 1 nMS-PCR primer of ER-α
1.3.5RT-PCR法檢測ERα mRNA表達(1)RNA提取:培養傳代細胞加入不同濃度葛根素進行培養,取生長狀態良好的細胞,PBS沖洗2遍,加入1 mL TRIZOL,采用氯仿提取、異丙醇沉淀、75%乙醇洗滌、DEPC溶解RNA。詳細過程按照TRIZOL說明進行,紫外分光光度計測量總RNA濃度,以保證cDNA合成時模板量一致。按Invitrogen公司RNA分離試劑盒說明書進行操作,提取RNA備用。(2)cDNA制備:總RNA經鑒定純度和濃度符合要求后,用于制備cDNA。①加入1~2 μg總RNA和1 μL50 pmol/L Oligo(dT)18 primer至經過焦炭酸二乙酯(diethypyrocarbonate,DEPC)處理后的EP管中,再加入DEPC水,使體積總量至12 μL。②依次加入下列試劑混合離心,4 μL5×RT Buffer,1 μL RNase Inhibitor(10 U/mL),2 μL dNTP Mixture(10 mmol/L),1 μL ReverTra Ace。所得產物用于PCR擴增或置于-20 ℃保存。(3)PCR擴增目標cDNA:① 引物設計 根據Genebank中查詢到的登錄號NM000125的ERα基因mRNA序列,使用primer 5.0軟件設計引物序列如下,GADPH管家基因用做內參照,引物由上海生物工程技術公司合成。ERα引物序列為5′-CCCTTG-CTATGTTACTAAGCGTGAGTGCC ATAGGA-ATACAAGAGGGTGCT-3′;ERα forward:5′-AG-CACCCTGAAGTCTCTGGA-3′;ERα reverse:5′-GATGTGGGAGAGGATGAGGA-3′。②PCR反應體系組分25 μL PCR反應體系包括Taq Polymerase,4種dNTP,Tris-HCl(pH8.3),50 mmol/L KC1,1.5 mmol/L MgCl2,1 μL上下游引物,3 μL cDNA模板。③ PCR反應 預變性94 ℃3 min后;變性94 ℃,退火61 ℃,延伸72 ℃各進行45 s 30個循環;最后終延伸72 ℃5 min結束。④反應產物的瓊脂糖凝膠電泳分析 取出5 μL PCR反應產物,混合1 μL 6×瓊脂糖凝膠電泳加樣緩沖液,上樣于0.5 ×TBE緩沖液配制的l%瓊脂糖凝膠中,在0.5×TBE緩沖液中90V電泳45 min,紫外分析儀下觀察結果,凝膠成像分析系統中分析PCR結果。
1.3.6MTT法培養成骨細胞0.25%胰蛋白酶消化后傳代,相同細胞密度(2×103/孔)接種于96孔培養板,常規培養48 h,細胞匯合90%,加入無血清DMEM/F12,依次加入5、10、25、50、100 μmol/L葛根素作用72 h。PBS沖洗,每孔加入10 μL0.5%MTT,孵育3 h,加100 μL 10%SDS,酶標儀上波長570 nm處,測定吸光度值(A)。細胞量與MTT吸光度值正相關。成骨細胞增殖促進率=(A干預組-A對照組)/A對照組×100%。
1.3.7堿性磷酸酶(alkaline phosphatase,ALP)活性測定用0.25%胰蛋白酶溶液消化半匯合成骨細胞,相同細胞密度(2×103/孔)接種于96孔培養板,常規培養48 h,細胞匯合90%,加入無血清DMEM/F12溶液,然后分別依次加入5、10、25、50、100 μmol/L葛根素作用72 h后p-NPP法測定。取培養液10 μL于96孔板,加p-NPP-DEA溶液90 μL,37 ℃水浴20 min,終止反應。酶標儀上波長405 nm處,測定吸光度值(A)。通過樣品A值在ALP標準曲線上讀取酶活性值(U/L)。成骨細胞分化促進率=(Z干預組ALP-Z對照組ALP)/Z對照組ALP×100%。
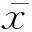
2結果
2.1培養細胞的鑒定
2.1.1活體細胞觀察倒置相差顯微鏡下,剛接種細胞呈球形,24 h后存活細胞基本貼壁,展開后細胞形態不規則,多呈三角形、多角形,有較多突起,單核呈卵圓形,含1~3個核仁,胞漿豐富。傳代培養,生長期細胞分裂相多見,突起互相連接,細胞重疊生長,呈鋪路石狀。細胞形態呈現梭形、多角形等多種形態(圖1)。
2.1.2ALP染色ALP為成骨細胞系合成分泌的標志性功能酶。分化成熟、功能活躍的成骨細胞質中ALP組織化學染色呈陽性反應。應用偶氮偶聯法染色,酶活性部位與偶氮染料形成不溶性藍色沉淀(圖2)。
2.1.3礦化結節茜素紅染色成骨細胞具有體外礦化的功能特征,細胞傳代培養,出現重疊生長,其間可見白色不透光圓形的礦化結節。茜素紅染色法礦化結節染色呈紅色,顯示成骨細胞的礦化結節(圖3)。

圖1 匯合狀OB(×100)Figure 1 Confluence of OB(×100)

圖2 OB ALP染色(×100)Figure 2 ALP dyeing of OB(×100)

圖3 OB礦化結節染色(×40)Figure 3 Mineral nodules dyeing of OB(×40)
2.2葛根素對細胞ERα啟動子A區甲基化的影響不同濃度葛根素處理傳代成骨細胞8 d后,ERα啟動子甲基化狀態的結果顯示,隨著劑量增加,基因啟動子A 區甲基化程度逐漸減弱,25 μmol/L葛根素濃度時表達最弱,此后增加濃度無明顯減弱。隨著葛根素濃度逐漸增加(5、10、25、50、100 μmol/L),甲基化條帶變得越來越弱,25 μmol/L葛根素濃度時表達最弱;未甲基化條帶變得越來越強,成骨細胞25 μmol/L葛根素濃度培養時表達最強(圖4)。

圖4 葛根素干預成骨細胞后ERα啟動子A區甲基化狀態U:引物ER-pU的DNA序列中,經NaHSO3處理后未甲基化胞嘧啶被修飾為胸腺嘧啶的未甲基化PCR 條帶;M: 甲基化胞嘧啶經NaHSO3處理后未被修飾,DNA序列仍是胞嘧啶的甲基化PCR條帶
Figure 4The methylation status of ERa in cells treated by different concentrations of puerarin
2.3甲基化水平對細胞ERα mRNA表達的影響為明確ERα基因啟動子甲基化程度與ERα mRNA表達水平是否有相關性。應用半定量 RT-PCR 方法檢測成骨細胞中ERα mRNA的表達,細胞用不同濃度梯度葛根素處理12 d后進行檢測。發現隨著葛根素濃度增加,ERα 基因啟動子甲基化程度逐漸減弱;同時也發現隨著濃度增加,細胞中ERα mRNA表達逐漸增強,25 μmol/L葛根素濃度時表達最強。見圖5、表2。

圖5 葛根素處理后成骨細胞ERα mRNA表達Figure 5 The expressions of ERα mRNA in cells treated by different concentrations of puerarin
2.4葛根素對細胞增殖的影響MTT法測定成骨細胞增殖促進率,隨著葛根素濃度增加,成骨細胞的細胞增殖呈現顯著增加,25 μmol/L濃度培養最強,此后隨濃度增加,未再進一步增加,并呈現下降趨勢,見表2。
2.5葛根素對細胞分化的影響p-NPP法測定細胞中堿性磷酸酶活性的絕對值與細胞量的比值來分析細胞分化能力,了解葛根素對成骨細胞分化的作用。成骨細胞分別加入0、5、10、25、50、100 μmol/L葛根素培養72 h,成骨細胞ALP活性隨著葛根素濃度增加,成骨細胞分化促進率明顯增加,25 μmol/L葛根素濃度培養時最強,此后增加濃度時,細胞分化促進率未再進一步增加,見表2。

表2 葛根素處理后成骨細胞ALP活性、增殖促進率和ERα/GAPDH比值Table 2 ALP activity,cells proliferation and ERα/GAPDH on osteoblasts treated by puerarin
*P<0.05與對照組比較#P<0.01對照組比較(Dunntt檢驗)
3討論
雌激素缺乏是絕經后骨質疏松的主要原因,雌激素替代療法能有效地預防絕經后骨質疏松癥的發生。但應用雌激素替代治療也引發了某些安全問題,長期單獨使用雌激素導致子宮內膜癌、乳腺癌及血栓栓塞性疾病發生率升高[2]。葛根素是從中藥葛根中提取的單體化合物,是葛根的主要有效成分之一,其化學名為4,7-二羥基8-D吡喃葡萄糖醛基異黃酮,相對分子質量為 416,屬于植物雌激素,具有雌激素樣活性,而不良反應卻較少[3]。
研究發現ERα存在于多種細胞中,包括成骨細胞膜或細胞質中,如果雌激素結合ERα,通過核內ER經典基因組效應,雌激素與ER結合后受體發生空間構象改變而形成二聚體,從而暴露DNA結合區使特異DNA序列與之結合,即靶基因調節區的雌激素效應元件(estrogen response element,ERE)與之結合,并募集輔助因子形成轉錄啟始復合物而啟動基因轉錄,將會激發一系列效應,調控細胞信號轉導和多種細胞活動,促使細胞的增殖和分化[4-7]。隨著甲基化檢測手段的進展,DNA甲基化逐漸成為表觀遺傳學領域的一個研究熱點,DNA甲基化抑制基因表達,而去甲基化增強基因表達。研究表明雌激素增強ERα基因表達,并與DNA甲基化有關。Penolazzi等[8]研究原代成骨細胞和骨髓來源MG-63 、SaOS-2細胞系ERα基因表達與ERα中F區啟動子內CpG島的甲基化,定量RT-PCR分析認為ERα中F區啟動子內CpG島的甲基化控制ERα基因轉錄,直接或間接影響其在特定組織基因表達。Dansranjavin 等[9]研究發現BMSC成骨表型基因呈高甲基化狀態,而RT-PCR法檢測OC、OPN、OSX、Runx2 mRNA表達呈陰性。
葛根素等植物雌激素防治骨質疏松的報道越來越引起人們重視,葛根素不僅抑制骨吸收,而且促使骨形成。一方面是通過影響堿性磷酸酶的活性,促進成骨細胞的分化;另一方面是通過雌激素受體介導促進成骨細胞增殖的骨形成效應[10]。葛根素與雌激素結構類似,研究葛根素對成骨細胞ERα啟動子內CpG島甲基化的影響,及其與成骨細胞ERα表達的關系,有助于我們更清楚葛根素調控成骨細胞代謝的關鍵環節。
本研究結果顯示隨著葛根素濃度梯度增加,成骨細胞 ERα啟動子A 區甲基化程度逐漸減弱,25 μmol/L葛根素濃度時表達最弱,此后未進一步減弱;同時成骨細胞ERα mRNA表達增加,有明顯的劑量依賴性,25 μmol/L葛根素濃度組達到峰值,此后隨著濃度增加,細胞ERα表達未見明顯增加。本研究結果也顯示葛根素增加成骨細胞增殖分化,25 μmol/L葛根素組濃度最高,此后隨著濃度增加呈現下降趨勢;葛根素濃度太低(<5 μmol/L)時,細胞增殖分化不明顯;達到適合濃度后(25 μmol/L)顯著增強;但是濃度太高(>25 μmol/L)時,成骨細胞增殖分化有減弱趨勢,這可能為葛根素毒性作用損傷細胞所致,仍需要進一步研究。
綜上所述,葛根素達到適合濃度后可能通過抑制成骨細胞 ERα啟動子A 區甲基化水平,增強細胞ERα mRNA表達,進而促使成骨細胞增殖分化。因此,葛根素可能將是絕經后骨質疏松治療的一種有效藥物,并且與雌激素相比,它的不良反應明顯較少,有利于絕經后骨質疏松的長期治療。
[參考文獻]
[1]Galea GL,Meakin LB,Sugiyama T,et al.Estrogen receptor α mediates proliferation of osteoblastic cells stimulated by estrogen and mechanical strain,but their acute down-regulation of the Wnt antagonist Sost is mediated by estrogen receptor β[J].J Biol Chem,2013,288(13):9035-9048.
[2]Fuhrman BJ,Schairer C,Gail MH,et al.Estrogen metabolism and risk of breast cancer in postmenopausal women[J].J Natl Cancer Inst,2012,104(4):326-339.
[3]Udomsuk L,Chatuphonprasert W,Monthakantirat O,et al.Impact of Pueraria candollei var.mirifica and its potent phytoestrogen miroestrol on expression of bone-specific genes in ovariectomized mice[J].Fitoterapia,2012,83(8):1687-1692.
[4]Chen FP,Hu CH,Wang KC.Estrogen modulates osteogenic activity and estrogen receptor mRNA in mesenchymal stem cells of women[J].Climacteric,2013,16(1):154-160.
[5]Zaman G,Jessop HL,Muzylak M,et al.Osteocytes use estrogen receptor alpha to respond to strain but their ERalpha content is regulated by estrogen[J].J Bone Miner Res,2006,21(8):1297-1306.
[6]Almeida M,Iyer S,Martin-Millan M,et al.Estrogen receptor-α signaling in osteoblast progenitors stimulates cortical bone accrual[J].J Clin Invest,2013,123(1):394-404.
[7]Almeida M,Martin-Millan M,Ambrogini E,et al.Estrogens attenuate oxidative stress and the differentiation and apoptosis of osteoblasts by DNA-binding-independent actions of the ERalpha[J].J Bone Miner Res,2010,25(4):769-781.
[8]Penolazzi L,Lambertini E,Giordano S,et al.Methylation analysis of the promoter F of estrogen receptor alpha gene: effects on the level of transcription on human osteoblastic cells[J].J Steroid Biochem Mol Biol,2004,91(1/2):1-9.
[9]Dansranjavin T,Krehl S,Mueller T,et al.The role of promoter CpG methylation in the epigenetic control of stem cell related genes during differentiation[J].Cell Cycle,2009,8(6):916-924.
[10]Lagari VS,Levis S.Phytoestrogens in the prevention of postmenopausal bone loss[J].J Clin Densitom,2013,16(4):445-449. GL,Meakin LB,Sugiyama T,et al.Estrogen receptor α mediates proliferation of osteoblastic cells stimulated by estrogen and mechanical strain,but their acute down-regulation of the Wnt antagonist Sost is mediated by estrogen receptor β[J].J Biol Chem,2013,288(13):9035-9048.
(本文編輯:許卓文)
[收稿日期]2014-12-19;[修回日期]2015-01-06
[基金項目]甘肅省中醫藥管理局科研課題(GZK-2013-26)
[作者簡介]呂海宏(1969-),男,陜西西安人,蘭州大學第一醫院副主任醫師,醫學博士,從事骨質疏松疾病診治研究。
[中圖分類號]R735.7
[文獻標志碼]A
[文章編號]1007-3205(2015)04-0385-06
Puerarin reduces methylation of estrogen receptor α promoter in osteoblasts and regulates its proliferation and osteoblastic differentiation
LV Hai-hong,TANG Xu-lei,FU Song-bo
(Department of Endocrinology,the First Hospital of Lanzhou University,Lanzhou 730000,China)
[Abstract]ObjectiveThe study shows that puerarin regulates pathway of metabolism in osteoblasts and provides the theoretical foundation for treating methods of postmenopausal osteoporosis.MethodsCulture of primary calvarial osteoblasts in rats were digested,and used in experiments.Then,the cells were treated with different concentrations of puerarin(5,10,25,50,100 μmol/L) in different groups,respectively.The methylation status of CpG islands in estrogen receptor α(ERα) gene promoter region of genome DNA was analyzed by nested-methylation-specific PCR;ER-α gene mRNA expression was examined by semiquantitative reverse transcription PCR analysis.The proliferation and viability of osteoblasts were assayed by MTT assay using the spectrophotometric method.Furthermore,the differentiation of osteoblast-like cells by p-nitrophenyl phosphate were identified.ResultsThe cultured osteoblasts treated by puerarin with different concentrations reduced in de novo methylation in promoter region of ERα gene with a concentration-treating manner,and a dose-dependent raising effect on osteoblasts proliferation and differentiation(P<0.01).Meanwhile,ER-α gene mRNA expressions in osteoblasts treated by puerarin with different concentrations became stronger and stronger,until in the 25 μmol/L treating group,their expressions were the stongest(P<0.01).ConclusionPuerarin reduces DNA methylation in promoter region of ERα in osteoblasts and enhences ERα gene mRNA expressions in osteoblasts.Above results possible influence its proliferation and osteoblastic differentiation.
[Key words]osteoblasts;puerarin;dna methylation;estrogen receptor α
