豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)(Epinephelus akaara)生長(zhǎng)、體組成及體脂肪酸組成的影響*
2016-01-15 01:40:15王驥騰姜宇棟楊云霞鄭普強(qiáng)盛建海
海洋與湖沼 2016年3期
王驥騰 姜宇棟 楊云霞 韓 濤 楊 敏 鄭普強(qiáng) 盛建海
(浙江海洋大學(xué)水產(chǎn)養(yǎng)殖系 舟山 316022)
脂肪是飼料的重要組成成分, 它不僅能為魚(yú)類(lèi)生長(zhǎng)提供能量, 還能為魚(yú)體提供必需脂肪酸。魚(yú)類(lèi)對(duì)脂肪的利用在很大程度上與其所含的必需脂肪酸相關(guān)(Stickney et al, 1989)。然而, 不同脂肪源的脂肪酸組成不同, 魚(yú)油因其富含高不飽和脂肪酸, 一直被作為傳統(tǒng)水產(chǎn)飼料的主要脂肪源(李思萌等, 2015; 張紅娟等, 2015)。近年來(lái), 隨著水產(chǎn)飼料行業(yè)的快速發(fā)展,漁用飼料生產(chǎn)對(duì)魚(yú)油的需求與日俱增, 魚(yú)油資源的有限性和價(jià)格的上漲使得養(yǎng)殖成本不斷升高, 尋找適宜的脂肪源替代魚(yú)油已成為本領(lǐng)域的研究熱點(diǎn)。相比魚(yú)油, 許多植物油源產(chǎn)量穩(wěn)定、價(jià)格低廉, 且含有較高的不飽和脂肪酸, 能在一定程度上為魚(yú)類(lèi)生長(zhǎng)提供所需脂肪酸。豆油是世界上產(chǎn)量最大的植物油,且相對(duì)于其它植物油源, 豆油含有更高含量的多不飽和脂肪酸(Figueiredo-Silva et al, 2005; NRC, 2011),因此它已成為最具潛力的可替代魚(yú)油的脂肪源。Peng等(2008)發(fā)現(xiàn), 使用豆油替代飼料魚(yú)油含量的60%—80%后, 黑鯛(Acanthopagrus schlegeli)的生長(zhǎng)沒(méi)有發(fā)生顯著變化。Izquierdo等(2003)在對(duì)金頭鯛(Sparus aurate)和歐洲齒舌鱸(Dicentrarchus labrax)的研究中以不同比例油源混合物替代 60%的魚(yú)油, 沒(méi)有對(duì)生長(zhǎng)和肉質(zhì)產(chǎn)生顯著影響。但是, 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)影響的研究仍未見(jiàn)報(bào)道。
赤點(diǎn)石斑魚(yú)(Epinephelus akaara), 俗稱(chēng)紅斑, 是我國(guó)名貴的海產(chǎn)經(jīng)濟(jì)魚(yú)類(lèi)之一, 其肉質(zhì)鮮美, 營(yíng)養(yǎng)豐富, 深受?chē)?guó)內(nèi)外市場(chǎng)歡迎。隨著人工育苗及苗種生產(chǎn)技術(shù)的初步成功, 赤點(diǎn)石斑魚(yú)的養(yǎng)殖正在成為一個(gè)新的水產(chǎn)養(yǎng)殖產(chǎn)業(yè)(Wang et al, 2016)。但是, 目前關(guān)于赤點(diǎn)石斑魚(yú)幼魚(yú)營(yíng)養(yǎng)需要量的研究較少, 僅見(jiàn)于對(duì)其最適飼料蛋白含量(Wanget al, 2016)及適宜飼料脂肪含量的研究(Jianget al, 2015)。因此, 本研究以豆油替代飼料中不同比例的魚(yú)油, 探究其對(duì)赤點(diǎn)石斑魚(yú)生長(zhǎng)、體組成及體脂肪酸的影響, 旨在為赤點(diǎn)石斑魚(yú)專(zhuān)用飼料的研發(fā)提供基礎(chǔ)數(shù)據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)飼料
以魚(yú)粉、酪蛋白為蛋白源, 玉米淀粉為糖源, 以豆油分別替代 25% (S25)、50% (S50)、75% (S75)和100% (S100)的魚(yú)油, 配制 4組等氮(50.26%—51.03%)、等能(20.82—21.01kJ/g)實(shí)驗(yàn)飼料(表 1)。實(shí)驗(yàn)飼料的脂肪酸組成見(jiàn)表2。
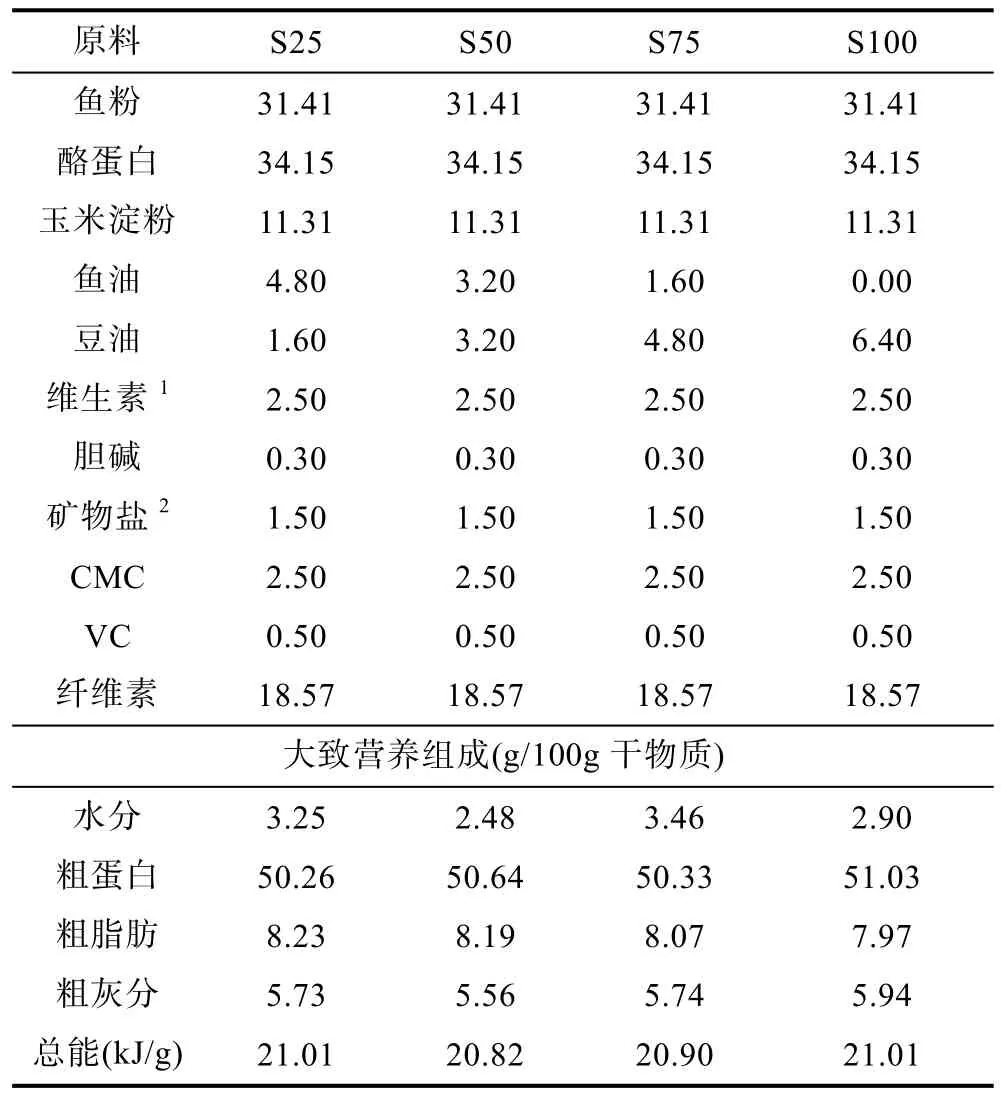
表1 實(shí)驗(yàn)飼料配方及營(yíng)養(yǎng)組成(%)Tab.1 Formulation and composition of experimental diets (%)
所有干物質(zhì)原料經(jīng)過(guò)粉碎, 過(guò) 60目篩后放入混合機(jī)中進(jìn)行充分混合, 其后加入油源和適量的蒸餾水再次混合均勻, 之后使用雙螺桿擠壓機(jī)將飼料制成2.5mm直徑的顆粒。飼料顆粒在45°C下干燥12h后保存在–20°C冰箱備用。
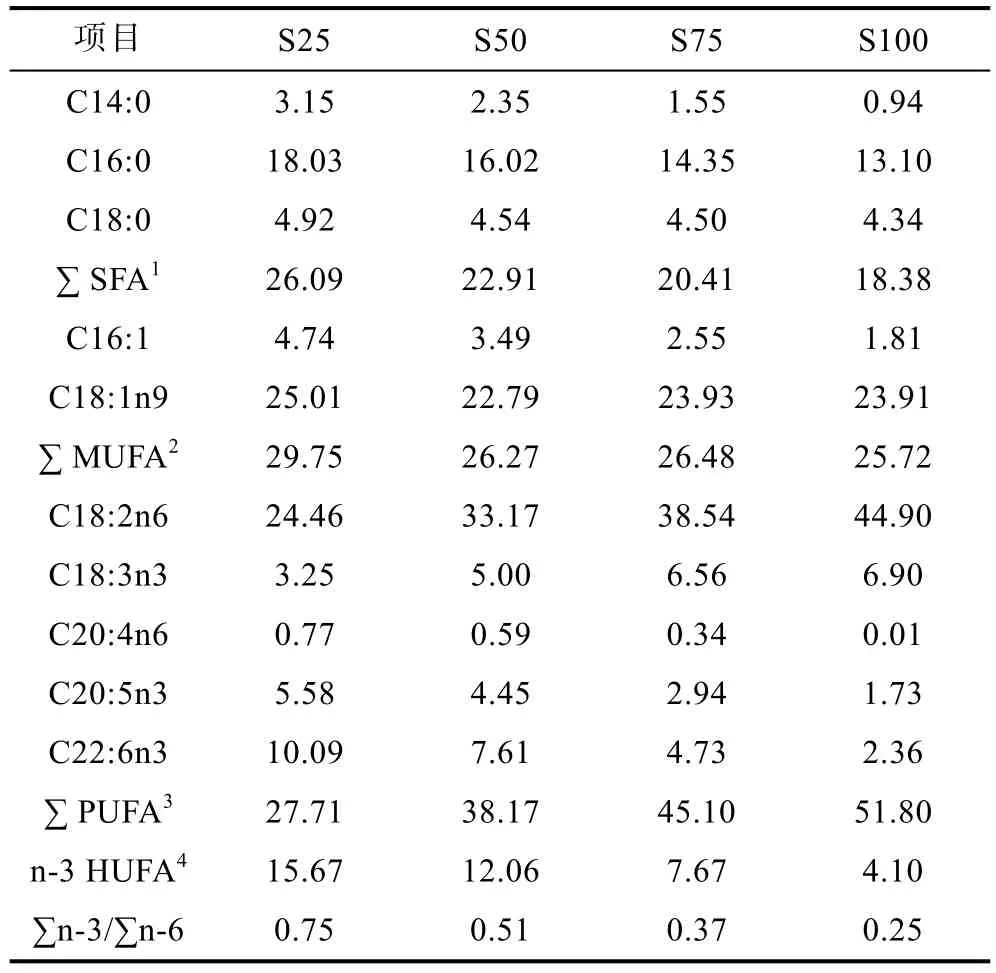
表2 實(shí)驗(yàn)飼料的脂肪酸組成Tab.2 Fatty acid composition of experimental diets
1.2 實(shí)驗(yàn)魚(yú)及養(yǎng)殖管理
實(shí)驗(yàn)用魚(yú)來(lái)自寧波一個(gè)商業(yè)海水魚(yú)育苗場(chǎng)的同一批次幼魚(yú), 飼養(yǎng)實(shí)驗(yàn)在浙江海洋大學(xué)營(yíng)養(yǎng)實(shí)驗(yàn)室(浙江舟山)進(jìn)行。實(shí)驗(yàn)開(kāi)始前, 幼魚(yú)每天用暫養(yǎng)顆粒飼料投喂2次, 暫養(yǎng)過(guò)程持續(xù)2周。暫養(yǎng)結(jié)束后, 隨機(jī)挑選 240尾健康且規(guī)格相近的幼魚(yú)隨機(jī)分配到 12個(gè)500L圓形PE養(yǎng)殖桶。實(shí)驗(yàn)設(shè)有4個(gè)處理, 每個(gè)處理分3個(gè)平行, 每個(gè)平行20尾幼魚(yú)(初始重大約為6g)。
實(shí)驗(yàn)為期6周, 每天9:00和18:00分2次進(jìn)行飽食投喂。使用配備微濾機(jī)、砂率缸、蛋白質(zhì)泡沫機(jī)、紫外消毒等設(shè)備的循環(huán)水過(guò)濾系統(tǒng), 實(shí)驗(yàn)期間, 水溫維持在(28±2)°C, 鹽度維持在(25±1)g/L, 溶解氧高于6mg/L, 亞硝酸鹽濃度低于0.1mg/L。
1.3 樣品采集
實(shí)驗(yàn)開(kāi)始前, 隨機(jī)留取18尾初始魚(yú)液氮冷凍后,置于超低溫冰箱中備用。養(yǎng)殖實(shí)驗(yàn)結(jié)束, 禁食24h后,對(duì)每個(gè)養(yǎng)殖缸的石斑魚(yú)進(jìn)行計(jì)數(shù), 稱(chēng)重并記錄。每個(gè)平行缸隨機(jī)獲取3尾魚(yú)用于全魚(yú)成分分析; 另從各平行隨機(jī)獲取6尾, 經(jīng) MS-222麻醉后稱(chēng)重解剖, 稱(chēng)取內(nèi)臟團(tuán)、肝臟及腸脂。取背部肌肉、肝臟經(jīng)由液氮冷凍后, 儲(chǔ)存于–76°C超低溫冰箱中, 用于水分、蛋白質(zhì)、脂肪含量及脂肪酸組成分析。
1.4 成分分析及脂肪酸分析
飼料和全魚(yú)成分分析采用AOAC(1995)方法。粗蛋白(N×6.25)通過(guò)凱氏定氮法測(cè)得(K355/K437, Buchi,Flawil, Switzerland); 粗脂肪使用索氏抽提法測(cè)得(E816, Buchi, Flawil, Switzerland); 水分采用冷凍干燥法測(cè)得(Freeze Dryer LL1500, Thermo Fisher Scientific, Waltham, USA), 灰分使用馬弗爐550°C灼燒法測(cè)定。
用于脂肪酸測(cè)定的飼料及全魚(yú)樣品在氯仿-甲醇液(2 : 1,V/V)中均質(zhì)用于提取脂肪。脂肪酸的甲酯化物(FAME)使用氣相色譜儀(GC 7890B, Agilent Technologies, Santa Clara, USA)進(jìn)行分析。以毛細(xì)管柱(60m×250μm; DB-23 Agilent, America)采用程序升溫(初始溫度為70°C, 以20°C /min程序升溫至150°C,其后以 6°C/min 升至 180°C, 再以 20°C/min 升至220°C, 保持 6min)進(jìn)行分離, 載氣為氮?dú)?氣壓:140.43kpa; 流速: 1mL/min)。進(jìn)樣溫度和檢測(cè)器溫度分別為 250°C 和 300°C。
1.5 數(shù)據(jù)統(tǒng)計(jì)分析
實(shí)驗(yàn)數(shù)據(jù)均以“平均值±標(biāo)準(zhǔn)差”表示。所有數(shù)據(jù)經(jīng) Microsoft Office Excel 2007初步整理后, 采用SPSS 18.0 (IBM, Chicago, USA) for Windows軟件進(jìn)行單因子方差分析(ANOVA), 若存在顯著性差異, 則用Duncan’s法多重比較, 檢驗(yàn)均值的差異顯著性, 顯著水平為P<0.05。
2 結(jié)果
2.1 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)生長(zhǎng)及飼料利用的影響(表3)
由表3可知, 盡管在各個(gè)飼料處理組之間的體增重(WG)和特定生長(zhǎng)率(SGR)未發(fā)現(xiàn)有統(tǒng)計(jì)學(xué)上的差異(P>0.05), 但是豆油 100%替代魚(yú)油組的 WG 及SGR值還是相對(duì)明顯較低。各實(shí)驗(yàn)組之間的日攝食量(DFI)沒(méi)有顯著性差異(P>0.05)。飼料系數(shù)(FCR)值受到飼料處理的顯著影響(P<0.05), 隨著飼料豆油替代水平的升高, 呈現(xiàn)先下降后上升的趨勢(shì), FCR最高值(1.96±0.42)出現(xiàn)在100%替代水平組, 25%—75%替代水平的3個(gè)組之間沒(méi)有顯著差異(P>0.05)。蛋白質(zhì)效率(PER)表現(xiàn)為相反的結(jié)果, 100%替代水平組呈現(xiàn)最低的值(1.06±0.21)。
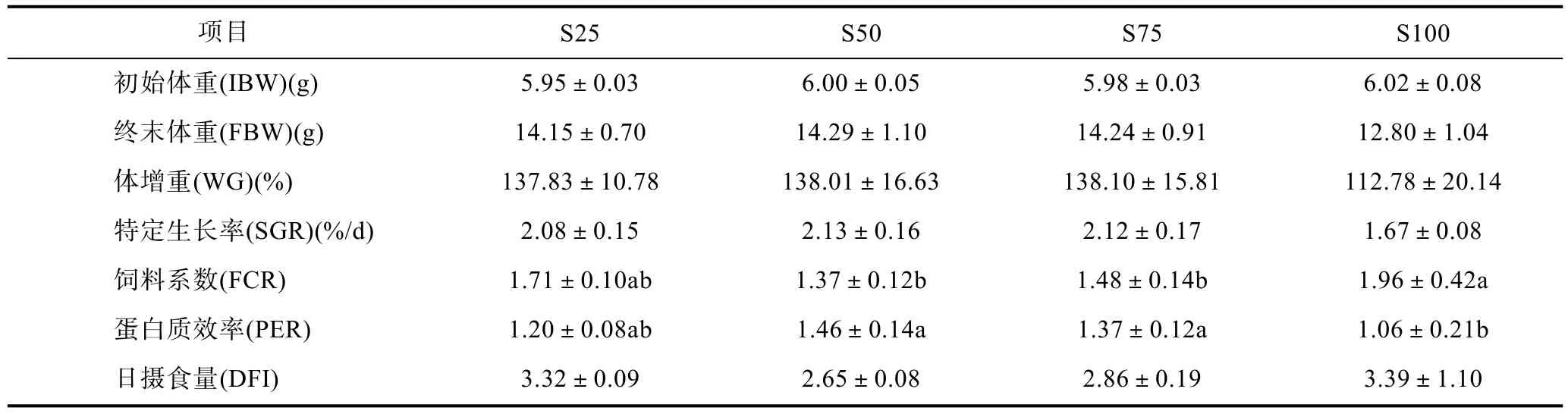
表3 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)生長(zhǎng)及飼料利用的影響Tab.3 Effect of dietary fish oil replacement by soybean oil on growth performance and feed utilization of red spotted grouper E.akaara
2.2 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)生物學(xué)指標(biāo)及體組成的影響(表4, 表5)
由表4可知, 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)生物學(xué)指標(biāo)的影響不顯著。100%豆油替代組呈現(xiàn)相對(duì)最高的肝體比(HSI)值, 但各處理組之間沒(méi)有顯著性差異(P>0.05)。全魚(yú)水分和蛋白含量沒(méi)有明顯差異(P>0.05)(表 5)。投喂 25%替代水平組的全魚(yú)脂肪含量最高(7.36±0.32), 顯著高于 75%和 100%替代水平組, 但是在 50%—100%替代水平處理組之間沒(méi)有顯著差異被發(fā)現(xiàn)(P>0.05)。肌肉水分、蛋白及脂肪含量沒(méi)有隨豆油替代水平的增加發(fā)生顯著變化(P>0.05)。
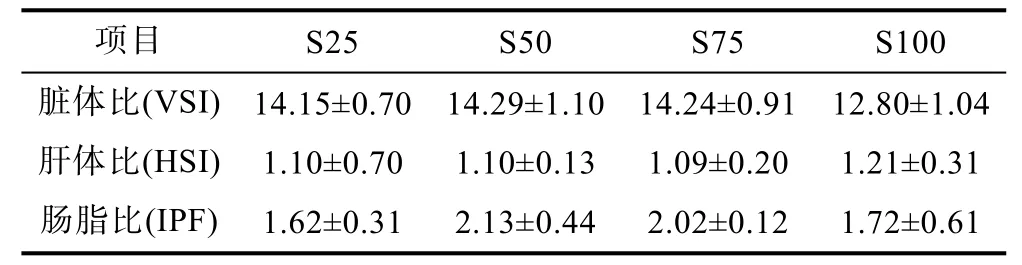
表4 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)生物學(xué)指標(biāo)的影響Tab.4 Effect of dietary fish oil replacement by soybean oil on biometric parameters of red spotted grouper E. akaara
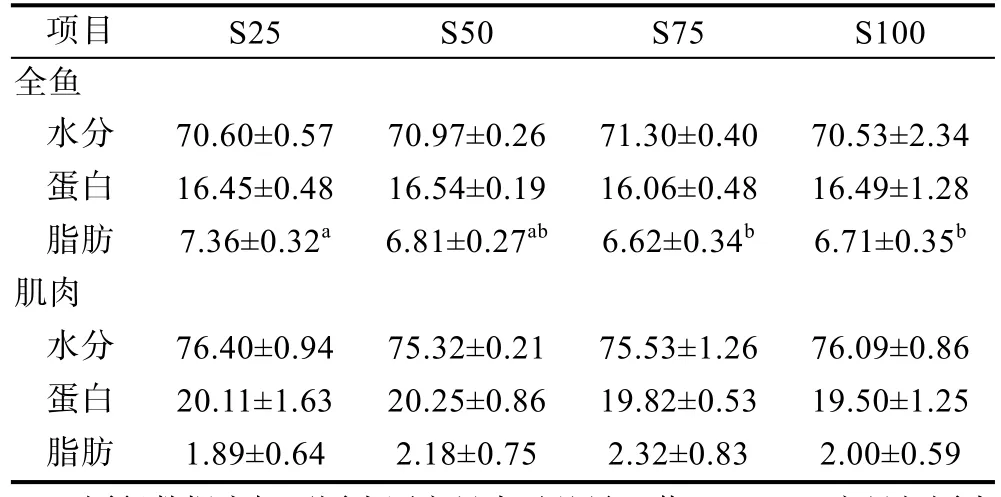
表5 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)體組成的影響Tab.5 Effect of dietary fish oil replacement by soybean oil on body composition of red spotted grouper E. akaara
2.3 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)魚(yú)體脂肪酸組成的影響(表6)
由表6可知, 赤點(diǎn)石斑魚(yú)魚(yú)體脂肪酸組成明顯受到飼料脂肪酸組成的影響。隨著飼料豆油水平的升高,魚(yú)體中亞油酸(C18:2n6)和多不飽和脂肪酸(PUFA)含量顯著升高(P<0.05); 全魚(yú)的 ARA(C20:4n6)、EPA(C20:5n3)和DHA(C22:6n3)含量在全豆油替代組最低,但是沒(méi)有顯著性差異被觀察在 25%—75%豆油替代組之間(P>0.05)。
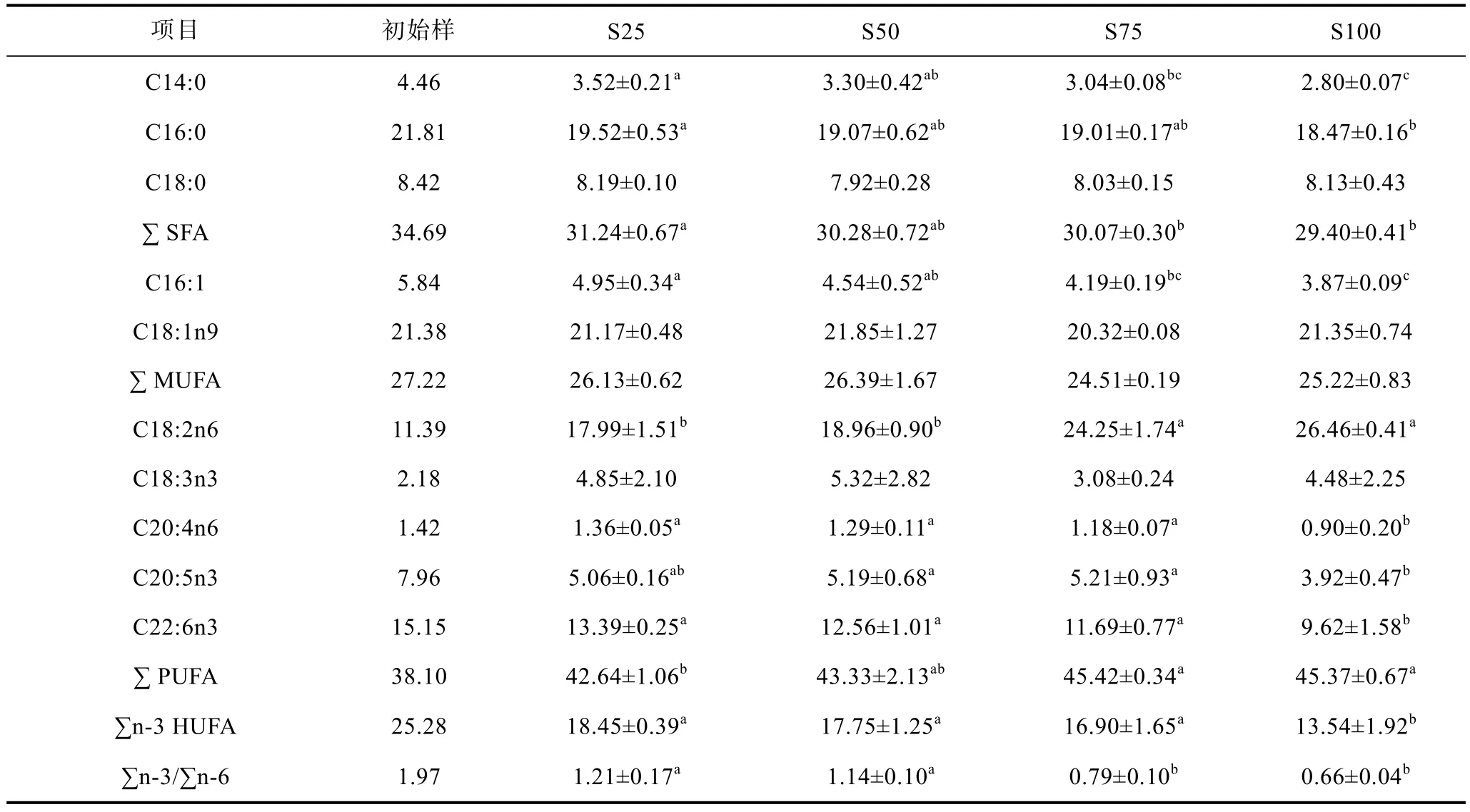
表6 豆油替代魚(yú)油對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)魚(yú)體脂肪酸組成的影響Tab.6 Effect of dietary fish oil replacement by soybean oil on body fatty acids composition of red spotted grouper E. akaara
3 討論
研究結(jié)果表明, 經(jīng)過(guò)6周的養(yǎng)殖實(shí)驗(yàn), 不同飼料處理未對(duì)赤點(diǎn)石斑魚(yú)幼魚(yú)的生長(zhǎng)產(chǎn)生顯著影響, 相似的結(jié)果也被報(bào)道在虹鱒(Oncorhynchus mykiss)(徐奇友等, 2009; Thanuthonget al, 2011a, b)和哲羅魚(yú)(Hucho taimen)(王炳謙等, 2006)的研究中。而在黑鯛(Penget al, 2008)和大菱鲆(Scophthalmus maximus)(Penget al, 2014)的研究中發(fā)現(xiàn), 飼料中過(guò)高的豆油替代水平會(huì)顯著降低養(yǎng)殖魚(yú)類(lèi)的生長(zhǎng)。在本實(shí)驗(yàn)中,盡管各處理組之間的WG值沒(méi)有顯著性差異存在, 但是全豆油組的生長(zhǎng)還是明顯低于其余各組。此外, 不同的飼料處理對(duì)FCR和PER值具有顯著性影響, 全豆油組有最高的FCR及最低的PER值, 這表明在飼料中使用豆油完全替代魚(yú)油降低了赤點(diǎn)石斑魚(yú)幼魚(yú)對(duì)飼料的利用率。這與 Xu等(2012)對(duì)長(zhǎng)鰭籃子魚(yú)(Siganus canaliculatus)的研究結(jié)果相似。
研究表明, 高不飽和脂肪酸(HUFA)對(duì)海水魚(yú)類(lèi)生長(zhǎng)、健康和繁殖發(fā)揮著重要作用, 特別是二十二碳六烯酸(DHA)和二十碳五烯酸(EPA), 為大部分海水魚(yú)類(lèi)的必需脂肪酸。Turchini等(2009)認(rèn)為, 飼料中過(guò)高的植物油會(huì)降低飼料HUFA的含量, 從而降低養(yǎng)殖魚(yú)類(lèi)的生長(zhǎng)和對(duì)飼料的利用效率。由于大多數(shù)海水魚(yú)類(lèi)自身合成EPA和DHA的能力十分有限, 因此飼料中通常需要添加EPA和DHA的總量在0.8%—2%之間才能滿(mǎn)足大部分海水魚(yú)類(lèi)的需求(NRC, 2011)。而在本研究中, 全豆油替代組飼料的EPA和DHA的總量?jī)H為 0.3%左右, 遠(yuǎn)低于大部分海水魚(yú)類(lèi)的需要量,這可能是在本實(shí)驗(yàn)條件下, 全豆油組有相對(duì)低的生長(zhǎng)和顯著低的飼料利用率的重要原因之一。此外, 與25%的替代水平組相比, 50%和 75%替代組飼料中有相對(duì)更低的EPA和DHA含量, 然而全魚(yú)中的EPA和DHA的含量在這三個(gè)處理組之間沒(méi)有存在顯著性差異。相似的結(jié)果也見(jiàn)于其它魚(yú)類(lèi)的報(bào)道, 如軍曹魚(yú)(Rachycentron canadum)(王驥騰等, 2007)、虹鱒(Caballero et al, 2002)、大西洋鮭(Salmo salar)(Bell et al, 2001)和大菱鲆(Bell et al, 1995)等。Skonberg等(1994)認(rèn)為這是由于魚(yú)類(lèi)會(huì)選擇性的保留某些重要的脂肪酸。通過(guò)對(duì)比飼料及全魚(yú)脂肪酸組成, 我們發(fā)現(xiàn),EPA和 DHA的保留率隨著替代水平的升高而升高,這在一定程度上表明EPA和DHA可能是赤點(diǎn)石斑魚(yú)的必需脂肪酸。盡管全豆油組的生長(zhǎng)和其它處理組之間沒(méi)有顯著差異存在, 但是全魚(yú)中EPA和DHA含量卻顯著低于其它各處理組, 這進(jìn)一步表明全豆油組飼料中的EPA和DHA含量是不足的。
通常情況下, 魚(yú)體脂肪酸組成在一定程度上反映了飼料中脂肪酸的組成(張紅娟等, 2015)。在本實(shí)驗(yàn)中, C18:2n6作為豆油中的特征脂肪酸, 在全魚(yú)魚(yú)體中的含量隨飼料中 C18:2n6含量的升高而顯著上升。然而, 隨著飼料中C18:3n3含量的升高, 全魚(yú)魚(yú)體中 C18:3n3的比例在各處理組之間卻沒(méi)有顯著性的差異存在。相似的結(jié)果也被發(fā)現(xiàn)在對(duì)虹鱒的研究中,作者認(rèn)為這可能是由于C18:3n3比C18:2n6在魚(yú)體中更容易被用于能量代謝(Thanuthong et al, 2011)。大量的研究表明, 盡管高水平的植物油替代魚(yú)油不會(huì)對(duì)生長(zhǎng)產(chǎn)生顯著性影響, 但是高的植物油替代水平往往會(huì)降低魚(yú)體中 HUFA的含量(Bell et al, 2001;Caballero et al, 2002)。海水魚(yú)作為人類(lèi)一個(gè)重要的食物來(lái)源, 除了為人類(lèi)提供了優(yōu)質(zhì)的蛋白質(zhì), 更重要的營(yíng)養(yǎng)價(jià)值是為人類(lèi)提供了n-3系列的HUFA(張紅娟等,2015), 特別是EPA和DHA, 對(duì)人體的心血管健康以及嬰幼兒的神經(jīng)發(fā)育十分有益。因此, 魚(yú)體中HUFA的含量通常體現(xiàn)出魚(yú)的營(yíng)養(yǎng)價(jià)值。在本研究中, 隨著飼料豆油對(duì)魚(yú)油替代水平的升高, 飼料中的HUFA以及 n-3/n-6表現(xiàn)出下降的趨勢(shì), 最低的值出現(xiàn)在全豆油組。考慮到食用魚(yú)的營(yíng)養(yǎng)價(jià)值和品質(zhì), 豆油對(duì)魚(yú)油的完全替代是不合適的。
臟體比、肝體比和腸脂比是用于評(píng)價(jià)魚(yú)體形態(tài)變化的常用指標(biāo)(姚林杰等, 2015), 這些指標(biāo)在很大程度上決定了食用魚(yú)的品質(zhì)優(yōu)劣及商品價(jià)值(金鑫等,2014)。植物油替代魚(yú)油對(duì)魚(yú)體體組成及肌肉組成的影響已經(jīng)被大量報(bào)道。Peng等(2008)研究發(fā)現(xiàn), 大菱鲆幼魚(yú)的肝體比隨著豆油替代水平的上升而升高,作者認(rèn)為這可能是由于在高比例的植物油替代組肝臟沉積了更多的脂肪。然而在本實(shí)驗(yàn)中, 臟體比、肝體比和腸脂比均未受豆油替代水平顯著影響。此外,與25%的替代組比, 大于75%的替代水平組有顯著更低的體脂肪含量。黃裕等(2015)認(rèn)為, 飼料中的PUFA會(huì)抑制脂肪酸合成酶基因的表達(dá), 促進(jìn)脂肪氧化分解基因的表達(dá), 從而抑制脂類(lèi)合成, 加速脂類(lèi)分解,使魚(yú)體脂肪含量降低。在本研究中, 大于75%的豆油替代水平組與25%的替代組相比, 含有更高的PUFA,這可能是大于 75%替代水平組有顯著低的體脂肪積累的原因之一。此外, 在各處理組之間, 全魚(yú)水分、蛋白含量和肌肉組成均沒(méi)有存在顯著性差異。相似的結(jié)果也見(jiàn)于對(duì)大菱鲆(彭墨等, 2014)、黑鯛(Peng et al,2008)、長(zhǎng)鰭籃子魚(yú)(Turchini et al, 2009)的報(bào)道。
4 結(jié)論
綜上所述, 在本實(shí)驗(yàn)條件下, 飼料豆油替代水平不宜超過(guò) 75%, 過(guò)高的豆油替代水平會(huì)對(duì)赤點(diǎn)石斑魚(yú)飼料利用及生長(zhǎng)產(chǎn)生不利影響; 此外EPA和DHA在體內(nèi)被選擇性保留, 可能是赤點(diǎn)石斑魚(yú)的必需脂肪酸, 而過(guò)高的豆油替代水平會(huì)降低魚(yú)體中 EPA和DHA含量而影響?hù)~(yú)的品質(zhì)。
王炳謙, 徐奇友, 徐連偉等, 2006. 豆油代替魚(yú)油對(duì)哲羅魚(yú)稚魚(yú)生長(zhǎng)和體成分的影響. 中國(guó)水產(chǎn)科學(xué), 13(6): 1023—1027
王驥騰, 韓 濤, 田麗霞等, 2007. 3種植物油源對(duì)軍曹魚(yú)生長(zhǎng)、體組成和脂肪酸組成的影響. 浙江海洋學(xué)院報(bào)(自然科學(xué)版), 26(3): 237—245
李思萌, 吳立新, 姜志強(qiáng)等, 2015. 飼料脂肪源對(duì)大菱鲆幼魚(yú)生長(zhǎng)性能和肌肉脂肪酸組成的影響. 動(dòng)物營(yíng)養(yǎng)學(xué)報(bào), 27(5):1421—1430
張紅娟, 陳秀玲, 張瑞玲等, 2015. 海水魚(yú)對(duì)脂肪的需求及脂肪源替代研究進(jìn)展. 水產(chǎn)科學(xué), 34(2): 122—127
金 鑫, 徐鋼春, 杜富寬等, 2014. 饑餓脅迫對(duì)刀鱭形體、體成分及血液生化指標(biāo)的影響. 動(dòng)物學(xué)雜志, 49(6): 897—903
姚林杰, 葉元土, 蔡春芳等, 2015. 團(tuán)頭魴幼魚(yú)飼料中 α-亞麻酸、亞油酸的適宜含量. 動(dòng)物營(yíng)養(yǎng)學(xué)報(bào), 27(3): 766—774
徐奇友, 許 紅, 李 嬋等, 2009. 用豆油代替魚(yú)油對(duì)虹鱒生長(zhǎng)、非特異性免疫和組織酶活性的影響. 大連水產(chǎn)學(xué)院學(xué)報(bào), 24(2): 104—108
黃 裕, 王際英, 李寶山等, 2015. 小麥胚芽油替代魚(yú)油對(duì)半滑舌鰨幼魚(yú)生長(zhǎng)、體成分、血清生化指標(biāo)及脂肪代謝酶的影響. 中國(guó)水產(chǎn)科學(xué), 22(6): 1195—1208
彭 墨, 徐 瑋, 麥康森等, 2014. 菜籽油替代魚(yú)油對(duì)大菱鲆幼魚(yú)生長(zhǎng)、脂肪酸組成及脂肪沉積的影響. 水產(chǎn)學(xué)報(bào),38(8): 756—765
AOAC, 1995. Official Methods of Analysis of AOAC International. 16th ed. Arlington, VA: Association of Official Analytical Chemists
Bell J G, Castell J D, Tocher D R et al, 1995. Effects of different dietary arachidonic acid: docosahexaenoic acid ratios on phospholipid fatty acid compositions and prostaglandin production in juvenile turbot (Scophthalmus maximus). Fish Physiology and Biochemistry, 14(2): 139—151
Bell J G, McEvoy J, Tocher D R et al, 2001. Replacement of fish oil with rapeseed oil in diets of Atlantic salmon (Salmo salar)affects tissue lipid compositions and hepatocyte fatty acid metabolism. The Journal of Nutrition, 131(5): 1535—1543
Caballero M J, Obach A, Rosenlund G et al, 2002. Impact of different dietary lipid sources on growth, lipid digestibility,tissue fatty acid composition and histology of rainbow trout,Oncorhynchus mykiss. Aquaculture, 214(1—4): 253—271
Figueiredo-Silva A, Rocha E, Dias J et al, 2005. Partial replacement of fish oil by soybean oil on lipid distribution and liver histology in European sea bass (Dicentrarchus labrax) and rainbow trout (Oncorhynchus mykiss) juveniles.Aquaculture Nutrition, 11(2): 147—155
Izquierdo M S, Obach A, Arantzamendi L et al, 2003. Dietary lipid sources for seabream and seabass: growth performance,tissue composition and flesh quality. Aquaculture Nutrition,9(6): 397—407
Jiang Y D, Wang J T, Han T et al, 2015. Effect of dietary lipid level on growth performance, feed utilization and body composition by juvenile red spotted grouper (Epinephelus akaara). Aquaculture International, 23(1): 99—110
National Research Council (NRC), 2011. Nutrient Requirements of Fish and Shrimp. Washington DC, USA: National Academies Press, 71—73
Peng M, Xu W, Mai K S et al, 2014. Growth performance, lipid deposition and hepatic lipid metabolism related gene expression in juvenile turbot (Scophthalmus maximus L.) fed diets with various fish oil substitution levels by soybean oil.Aquaculture, 433: 442—449
Peng S M, Chen L Q, Qin J G et al, 2008. Effects of replacement of dietary fish oil by soybean oil on growth performance and liver biochemical composition in juvenile black seabream,Acanthopagrus schlegeli. Aquaculture, 276(1—4): 154—161
Skonberg D L, Rasco B A, Dong F M, 1994. Fatty acid composition of salmonid muscle changes in response to a high oleic acid diet. The Journal of Nutrition, 124(9):1628—1638
Stickney R R, Hardy R W, 1989. Lipid requirements of some warmwater species. Aquaculture, 79(1—4): 145—156
Thanuthong T, Francis D S, Manickam E et al, 2011a. Fish oil replacement in rainbow trout diets and total dietary PUFA content: II) Effects on fatty acid metabolism and in vivo fatty acid bioconversion. Aquaculture, 322—323: 99—108
Thanuthong T, Francis D S, Senadheera S D et al, 2011b. Fish oil replacement in rainbow trout diets and total dietary PUFA content: I) Effects on feed efficiency, fat deposition and the efficiency of a finishing strategy. Aquaculture, 320(1—2):82—90
Turchini G M, Torstensen B E, Ng W K, 2009. Fish oil replacement in finfish nutrition. Reviews in Aquaculture,1(1): 10—57
Wang J T, Jiang Y D, Li X Y et al, 2016. Dietary protein requirement of juvenile red spotted grouper (Epinephelus akaara). Aquaculture, 450: 289—294
Xu S D, Wang S Q, Zhang L et al, 2012. Effects of replacement of dietary fish oil with soybean oil on growth performance and tissue fatty acid composition in marine herbivorous teleost Siganus canaliculatus. Aquaculture Research, 43(9):1276—1286

- 海洋與湖沼的其它文章
- CPA-核酸試紙條快速檢測(cè)副溶血性弧菌(Vibrio parahaemolyticus)方法的建立及其在海產(chǎn)品檢測(cè)中的應(yīng)用*
- 日本囊對(duì)蝦(Marsupenaeus japonicus)秋繁同生群仔蝦不同耐干露性能群體間的形態(tài)表型差異*
- COⅠ條形碼輔助分析雷州半島紅樹(shù)林區(qū)魚(yú)類(lèi)的物種多樣性*
- 小黃魚(yú)(Pseudosciaena polyactis)形態(tài)性狀與體質(zhì)量的相關(guān)性及通徑分析*
- 基于微衛(wèi)星標(biāo)記的洞庭青鯽與三個(gè)鯽品系群體遺傳多樣性分析*
- 青魚(yú)(Mylopharyngodon piceus)新發(fā)病病原類(lèi)志賀鄰單胞菌(Plesiomonas shigelloides)的分離鑒定*