基于微衛星標記的洞庭青鯽與三個鯽品系群體遺傳多樣性分析*
2016-01-15 01:40:16劉良國陳海康楊品紅謝春華李夢軍
海洋與湖沼 2016年3期
劉良國 陳海康 許 倩 楊品紅 謝春華 李夢軍
(水產高效健康生產湖南省協同創新中心 動物學湖南省高校重點實驗室 湖南文理學院生命科學學院 常德 415000)
洞庭青鯽(Dongtingking crucian carp, Carassius auratus var. Dongtingking)是近年來本文作者在洞庭湖水系澧水北民湖水域發現的一個具有兩性繁殖方式的天然二倍體鯽魚新品系(楊品紅等, 2005, 2007;吳珊等, 2006)。由于其在經濟性狀上具有生長快、產量高、抗逆性強、味道鮮美等特點, 受到養殖戶和廣大消費者喜愛, 發展前景廣闊。在生物學特性上, 與國內已報道的其它鯽品系性狀相比, 洞庭青鯽與彭澤鯽和普通野鯽尤為相似: 彭澤鯽原產于江西省彭澤縣一帶, 最初的研究結果表明, 彭澤鯽也是一種二倍體兩性生殖魚類(傅永進, 1996), 但隨后報道的彭澤鯽均為染色體數目在 150條左右的雌核發育魚(Zhou et al, 2002; 劉良國等, 2004), 在染色體倍性和生殖方式上, 洞庭青鯽均與最初報道的彭澤鯽相似;同時, 我們采用核型分析方法對洞庭湖水系普通野鯽群體進行批量的染色體數目統計, 發現在洞庭湖水系中, 同時存在兩種在形態上極為相似、染色體數為100和150 ±的不同倍性野鯽群體, 而對洞庭青鯽染色體組型分析(楊品紅等, 2007)表明它與洞庭湖水系野生二倍體鯽倍性一致。洞庭青鯽作為一個新型的鯽魚品系, 它與彭澤鯽、二倍體和三倍體野鯽到底存在怎樣的遺傳關系, 這將是本論文重點關注的問題。
微衛星(Simple Sequence Repeat, SSR, 單一的序列重復)分子標記在真核生物基因組中廣泛存在, 因其具有多態性高、共顯性遺傳、可重復性強和高度特異性等優點, 現已廣泛應用于群體遺傳多樣性(周盼等, 2011; 宋春妮等, 2011)、基因連鎖與遺傳圖譜構建(Miyao et al, 1996)、品種鑒定、親本分析、純系檢驗等(Stockburgeet al, 2000; 劉靜霞等, 2003; Bessertet al, 2003; 文萍等, 2015)研究領域。為了了解洞庭青鯽與彭澤鯽、二倍體和三倍體野鯽的分子遺傳多樣性,揭示它們之間的遺傳關系, 本實驗選用 12個微衛星標記對洞庭青鯽與彭澤鯽、二倍體和三倍體野鯽的遺傳多樣性進行檢測分析, 實驗結果對于鯽的多倍體起源演化研究, 以及二倍體鯽的種質資源保護與選育種實踐具有重要意義。
1 材料與方法
1.1 材料
洞庭青鯽(F6)和彭澤鯽取自大湖股份湖南省水產工程技術研究中心實驗基地, 野生二倍體和三倍體鯽采自洞庭湖水系的沅水和澧水。實驗魚采回后, 首先通過活體染色體制片的方法, 進一步批量確定不同鯽品系的染色體倍性, 然后, 每個鯽品系群體各取樣30尾, 分別剪其背部肌肉用無水乙醇保存備用。
1.2 基因組DNA提取
每尾鯽樣品分別取1.0g肌肉用于基因組DNA提取, 提取方法參照常規的酚-氯仿抽提程序進行。DNA提取后, 用0.8%瓊脂糖凝膠電泳進行質量檢測,同時用紫外分光光度計估算其濃度, 調整濃度至40ng/μL, 4°C 冰箱保存備用。
1.3 微衛星引物
本實驗選用在銀鯽(周莉等, 2001; 孫效文等,2009)和紅鯽(李建中等, 2005; 魯雙慶等, 2005)中均能擴增出清晰的DNA譜帶, 且重復穩定性好的12對微衛星引物序列(表 1), 由北京鼎國昌盛生物技術有限責任公司合成為藍色(FAM)熒光標記引物。
1.4 微衛星擴增
PCR反應體系為25μL, 其中40ng/μL DNA模板2.0μL, 10mmol/L 引物各 0.5μL, 10mmol/L dNTPs 0.5μL, 10×PCR buffer 2.5μL, Taq DNA polymease 0.5μL (2U/μL), ddH2O 18.5μL。PCR 擴增程序為: 95°C 5min, 接著 94°C 30s, 退火 30s (退火溫度見表 1),72°C 30s, 共 35 個循環, 最后 72°C 10min。將帶有藍色(FAM)熒光標記的 PCR產物與紅色分子量內標ROX 500混合, 采用ABI3730X Genetic Analyzer對其進行毛細管電泳檢測, 得到擴增片段大小、峰圖等原始實驗數據; 同時, 將擴增產物在 4%的變性聚丙烯酰胺凝膠上電泳(50W 恒功率 2.5h), ABI PRISM 377測序儀掃描, 得到電泳圖。

表1 微衛星引物序列、退火溫度及等位基因大小Tab.1 Primer sequences, annealing temperature, and alleles size of the microsatellite markers
1.5 數據統計與分析
對毛細管電泳測序儀檢測得到的原始擴增條帶進行統計, 將每一條帶視為該位點的 1個等位基因,利用Genepop V4.0和EXCEL MS TK-2007 軟件進行分析, 計算并統計 12個微衛星基因座位的等位基因數和等位基因頻率、觀測雜合度(observed heterozygosity,Ho)、期望雜合度(expected heterozygosity, He)和多態信息含量(polymorphism information content, PIC)。
通過 Lynch(1990)的方法計算群體間的遺傳相似性系數和遺傳距離, 根據遺傳距離, 再以 MEGA 4.0軟件包中的非加權配對算術平均法(unweighted pair group methods using arithmetic average, UPGMA)對四個鯽品系群體進行聚類分析。
2 結果與分析
2.1 微衛星PCR擴增結果及等位基因頻率
12對微衛星引物在 4個鯽品系群體中均得到較好的擴增結果, 圖1為引物MFW4和MFW20在4個群體中的部分變性聚丙烯酰胺凝膠電泳圖譜。在 12個基因座位中, 每個座位檢測到的等位基因數 4—12個, 平均等位基因數 6.58, 共檢測出 78個等位基因,其中洞庭青鯽群體有 27個, 二倍體野鯽群體 49個,三倍體野鯽群體54個, 彭澤鯽群體34個, 78個等位基因中有14個等位基因為4個群體所共有。與二倍體和三倍體野鯽群體相比, 洞庭青鯽、彭澤鯽等養殖群體的等位基因數明顯偏少, 基因座位 MFW4的 1,MFW19的2, MFW23的3, SCM4的5、7, SCM8的1, SCM10的3、5、7, SCM13的5, SCM17的1、2、8等一些等位基因在兩個野生鯽群體中均有出現, 而在洞庭青鯽和彭澤鯽養殖群體中缺失。12個基因座位在4個鯽品系群體中的等位基因頻率見表2。
2.2 4個鯽品系群體的遺傳多樣性
12個微衛星位點在同一鯽品系群體, 以及同一個微衛星位點在不同鯽品系群體的遺傳變異程度均存在較大差異(表 3)。洞庭青鯽、野生二倍體和三倍體鯽、彭澤鯽群體的平均觀測雜合度分別為 0.633、0.750、0.800、0.717, 平均期望雜合度分別為0.502、0.713、0.757、0.602, 平均多態信息含量分別為0.364、0.599、0.637、0.470, 其中野生二倍體和三倍體鯽群體的平均觀測雜合度、期望雜合度和多態信息含量均高于洞庭青鯽和彭澤鯽養殖群體, 表現為高度多態,而洞庭青鯽和彭澤鯽養殖群體表現為中度多態, 部分座位如洞庭青鯽的MFW19和SCM13兩個位點PIC值為0, 遺傳多態性消失。
2.3 群體間的遺傳相似性系數、遺傳距離及聚類分析
根據等位基因頻率計算得到不同鯽品系群體之間的遺傳相似性系數和遺傳距離(表4), 計算結果表明, 4個鯽品系群體間的平均遺傳相似系數在0.421—0.787, 平均遺傳距離為 0.213—0.579。其中洞庭青鯽與彭澤鯽群體之間、野生二倍體和三倍體鯽群體之間的遺傳相似性系數較高, 分別為 0.787、0.621,遺傳距離較小, 分別為0.213、0.379; 而洞庭青鯽、彭澤鯽與野生二倍體鯽的遺傳相似性系數較低, 分別為 0.421、0.482, 遺傳距離較大, 分別為 0.579、0.518。
根據群體間的遺傳距離矩陣(表 4), 采用 MEGA 4.0軟件包中的UPGMA聚類法分析4個鯽品系群體間的親緣關系(圖2), 圖2顯示, 4個鯽品系群體明顯分為兩個大支, 其中, 洞庭青鯽與彭澤鯽聚為一支,野生二倍體與三倍體鯽聚為另一支, 表明洞庭青鯽與彭澤鯽養殖群體的親緣關系較近, 野生二倍體與三倍體鯽的親緣關系較近。
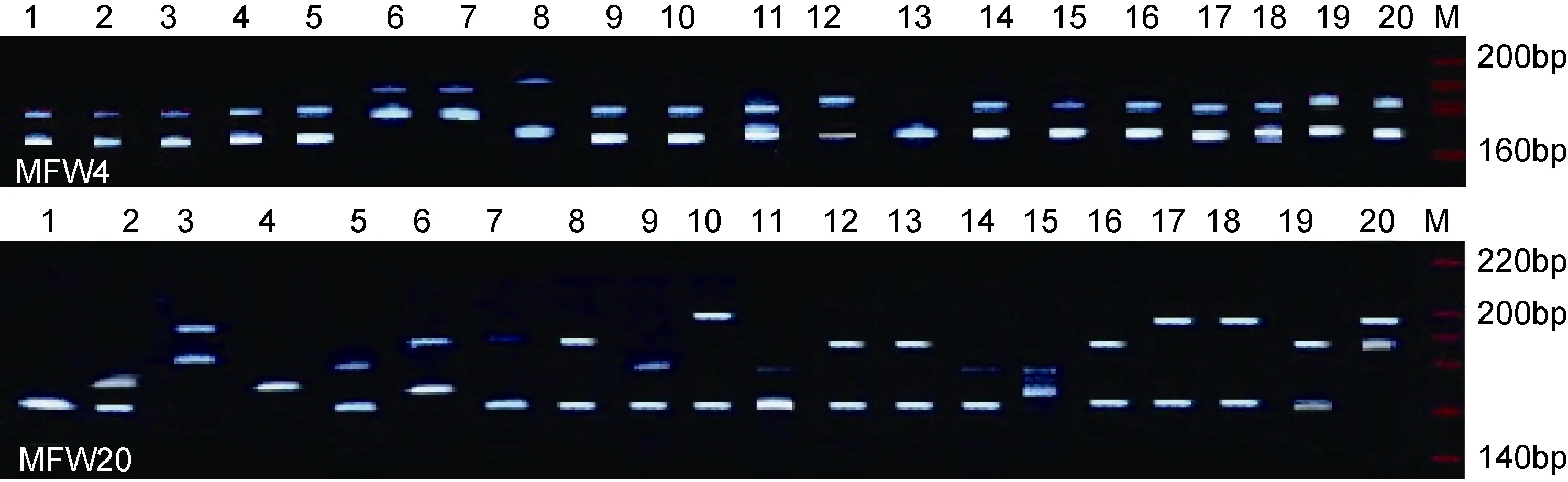
圖1 引物MFW4和MFW20在4個鯽品系群體中的部分擴增圖譜Fig.1 The SSR amplified results for the populations of 4 strains of crucian carp by the primer MFW4 and MFW20
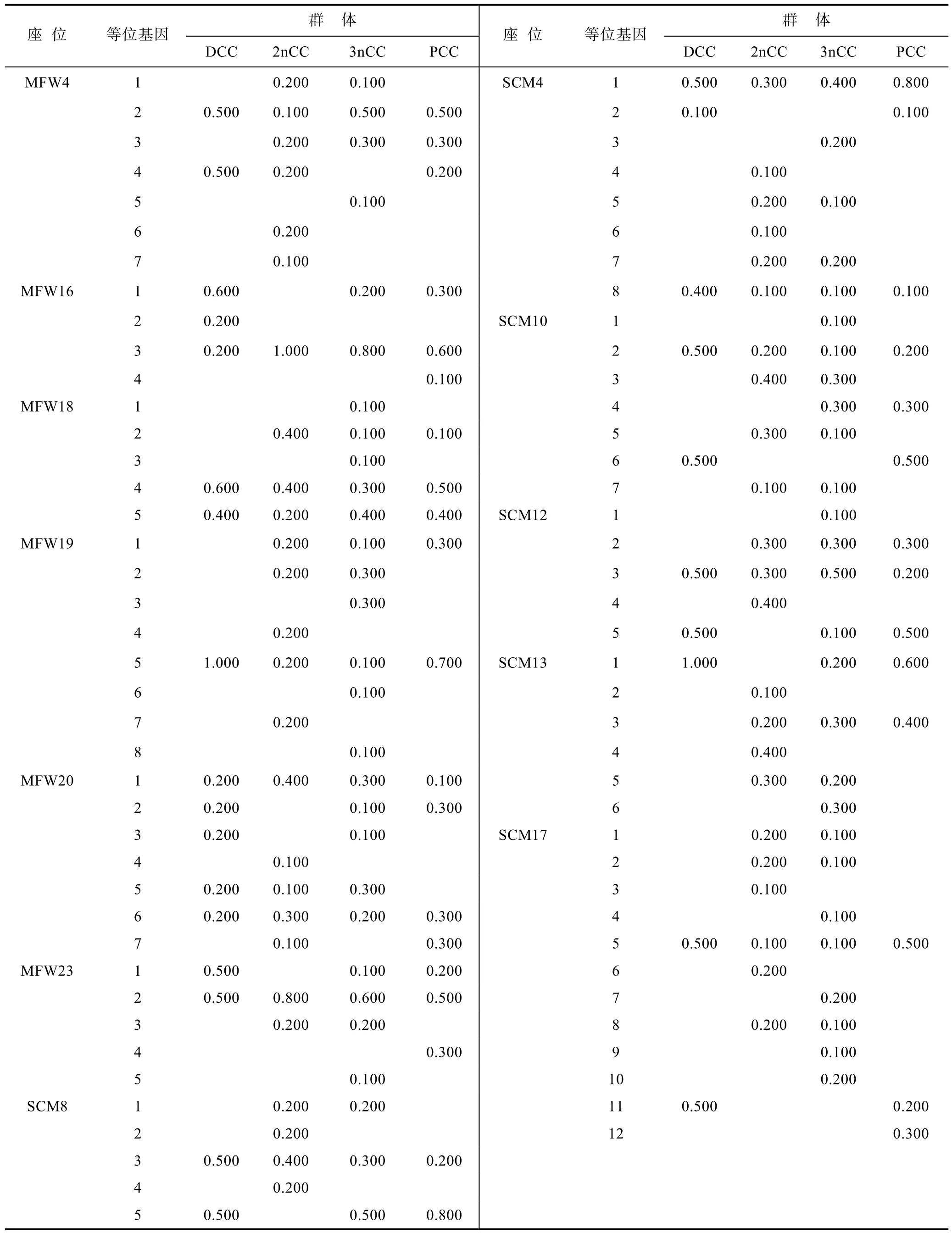
表2 4個鯽品系群體12個微衛星基因座位的等位基因頻率Tab.2 Frequency of 12 microsatellites DNA loci among the populations of 4 strains of crucian carp
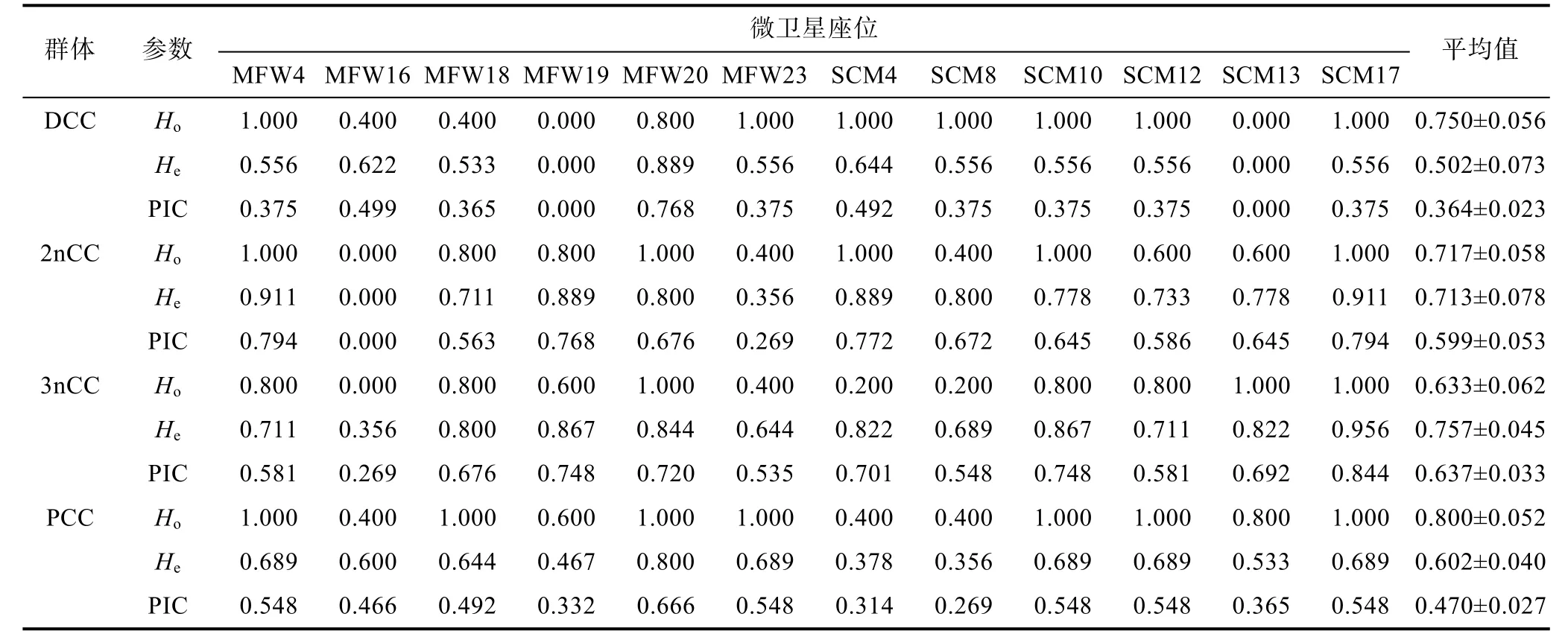
表3 4個鯽品系群體12個微衛星座位的觀測雜合度、期望雜合度和多態信息含量Tab.3 Observed heterozygosity, expected heterozygosity, and polymorphism information content at 12 microsattellite loci assessed for the populations of 4 strains of crucian carp
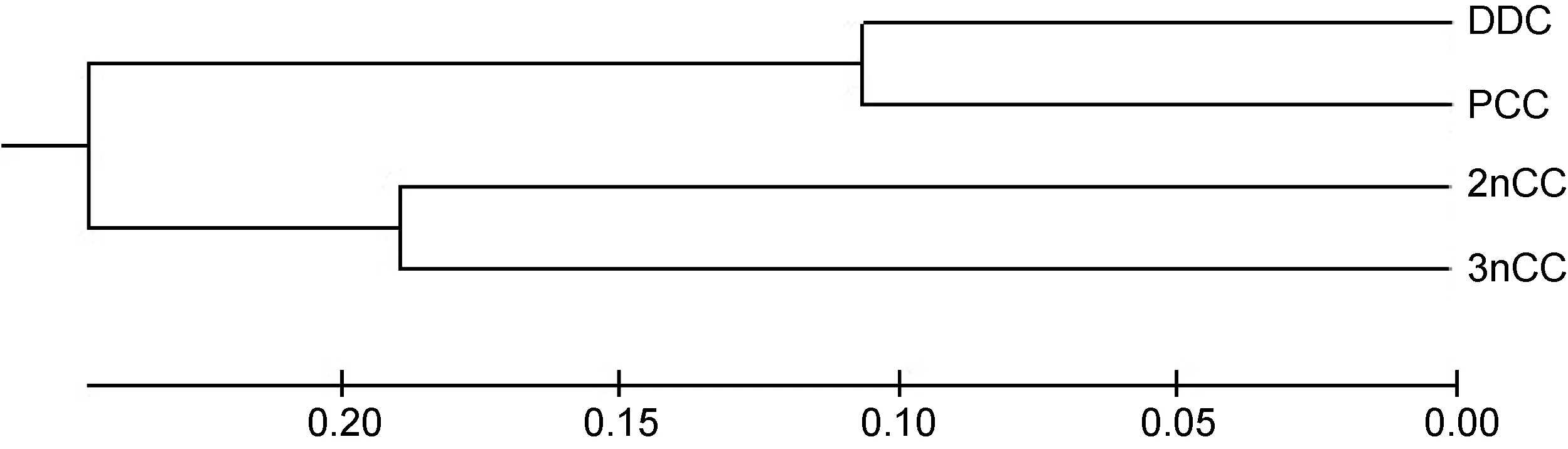
圖2 4個鯽品系群體的UPGMA聚類圖Fig.2 UPGMA dendrogram for the populations of 4 strains of crucian carp
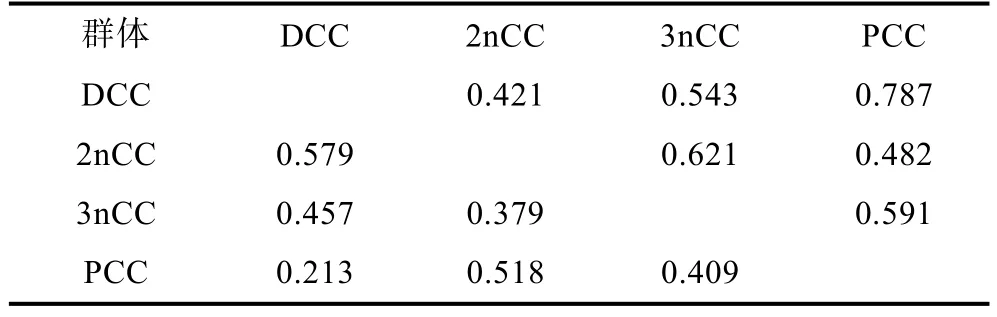
表4 4個鯽品系群體間的遺傳距離(對角線下方)及遺傳相似性系數(對角線上方)Tab. 4 The genetic distances (below diagonal) and genetic similarities (above diagonal) among the populations of 4 strains of crucian carp
3 討論
遺傳多樣性是指地球上生物所攜帶的各種遺傳信息的總和, 它包括生物遺傳變異大小和遺傳變異分布格局(即群體遺傳結構)等內容。它是物種適應多變的環境條件、維持生存和進化的基礎, 物種的遺傳多樣性越豐富, 其對環境的生存適應能力和進化潛力就越強(O’Connellet al, 1997; 張文靜等, 2003; 朱曉東等, 2007)。由于微衛星標記遵循孟德爾定律呈共顯性遺傳, 在檢測種群異質性方面有明顯優勢(Bártfaiet al, 2003), 因而能更多的揭示群體的遺傳變異水平。本研究利用12對微衛星標記對洞庭青鯽等4個鯽品系群體進行了比較分析, 12個微衛星位點共檢測到78個等位基因, 其中有14個等位基因為4個鯽品系群體所共有, 這些共有等位基因頻率相對較高, 推測是鯽進化過程中最原始、最保守的一部分,對于維持鯽物種的繁衍和穩定有著重要作用。其余等位基因, 如在野生鯽群體中存在而在養殖鯽群體中丟失, 或只在洞庭青鯽和彭澤鯽養殖鯽群體中出現而野生鯽群體中未見, 或只在某一個鯽品系群體中出現等情況(表 2), 可能是進化過程中由于基因的插入、缺失等突變機制所造成。
群體平均基因雜合度表示被檢測位點上群體中雜合子的頻率, 它是衡量群體雜合程度的指標(周盼等, 2011)。群體平均基因雜合度越低, 反映該群體的遺傳一致性越高, 即群體的遺傳變異越少, 遺傳多樣性越低(Leberg, 2002; 張燕萍等, 2013)。張志偉等(2006)報道了江蘇境內草魚1個野生群體和2個養殖群體的平均觀測雜合度為 0.5286—0.7000,平均期望雜合度為 0.5235—0.6939; 周盼等(2011)報道了三江水系4個野生草魚群體的平均觀測雜合度為0.7114—0.8045, 平均期望雜合度為0.6245—0.6739。本研究結果表明 4個鯽品系群體的平均觀測雜合度(Ho)介于 0.633—0.800, 平均期望雜合度(He)介于 0.502—0.757, 與以上兩個報道的結果相近, 說明本實驗選用的微衛星標記適合鯽品系群體的遺傳多樣性研究。從表3可見, 4個鯽品系群體的平均觀測雜合度與期望雜合度分析結果較為一致, 其中, 三倍體野鯽的平均觀測雜合度最高, 為 0.800, 二倍體野鯽次之, 為 0.750, 這說明洞庭湖水系野鯽群體的遺傳多態性豐富, 選育種潛力較大; 洞庭青鯽群體的平均觀測雜合度最低, 為 0.633, 說明洞庭青鯽養殖群體的遺傳多樣性水平較低, 這與洞庭青鯽這一優良鯽魚品系的多代的選育純化有關, 這一結果也與吳珊等(2006)對洞庭青鯽形態性狀的分析結果一致: 即洞庭青鯽子代與親代不僅沒有性狀分離現象, 而且基因有進一步純化的趨勢。
多態信息含量(PIC)是群體內遺傳變異的量度,可以作為微衛星位點變異程度指標(周盼等, 2011)。依據 Botstein等(1980)的劃分標準: 當 PIC>0.5時,該位點為高度多態位點; 當 0.25<PIC<0.5時, 為中度多態位點; 當 PIC<0.25時, 為低度多態位點。從本研究 12個微衛星位點來看(表 3), 高度多態位點主要集中在二倍體和三倍體野鯽群體, 二倍體野鯽平均PIC為0.599, 高度多態位點占總位點的83.3%,三倍體野鯽平均PIC為0.637, 高度多態位點占總位點的 91.7%, 說明洞庭湖水系二倍體和三倍體野鯽群體的遺傳多樣性豐富; 而中、低度多態位點主要集中于洞庭青鯽和彭澤鯽養殖群體, 洞庭青鯽平均PIC為 0.364, 中度多態位點占總位點的 75.0%,MFW19和SCM13兩個位點的PIC值為0, 彭澤鯽平均PIC為0.470, 中度和高度多態位點占總位點的比例各占 50.0%, 以上情況說明, 與基因雜合度分析的結果一致, 相比于野生鯽群體, 洞庭青鯽和彭澤鯽養殖群體的遺傳多樣性降低, 尤以洞庭青鯽遺傳多樣性下降為甚。
群體間的遺傳距離是遺傳變異的尺度, 遺傳距離的大小預示著親緣關系的遠近。本研究用微衛星實驗數據分析得到的遺傳距離和樹狀圖與先前ISSR的分析結果(劉良國等, 2015)基本一致, 即洞庭青鯽與彭澤鯽的遺傳距離最小, 樹狀圖上聚為一支, 二者親緣關系較近, 主要原因可能是洞庭青鯽與彭澤鯽均起源于天然水域, 因其都具有優良的生產性狀而被多代選育, 在相似的人工養殖環境下, 選擇壓力相近所致; 二倍體和三倍體野鯽在樹狀圖上聚為另一支,表明二者親緣關系也較近, 其原因可能要追溯到三倍體野鯽的起源上, 洞庭湖水系二倍體和三倍體野鯽生活于同一水體, 二者在形態上極為相似(劉良國等, 2012), 在極端環境(如溫度的劇變等)條件下, 二倍體鯽有可能產生二倍體卵子, 這些二倍體卵子再與正常的單倍體精子雜交形成三倍體, 因而在基因座位上, 三倍體野鯽表現出與二倍體野鯽的相似性。
據沈俊寶等(1983)和王蕊芳等(1988)報道, 由于環境急劇變化和天然雜交等因素, 自然水體中的二倍體鯽可以演化成三倍體, 而三倍體鯽具有雌核發育生殖特性, 繁殖力和適應性強, 種群優勢明顯, 這可能是當前天然水域中二倍體鯽數量遠少于三倍體鯽的重要原因。洞庭青鯽是近年來發現并選育出的生長速度快、營養價值高、遺傳性狀穩定的二倍體鯽品系, 是自然界中十分珍貴的鯽魚種質資源, 具有很高的經濟價值和科研價值。由于洞庭青鯽選育的時間較短, 至今才經過6代, 將來能否保持其二倍體穩定的遺傳性狀還未為可知。根據上述三倍體鯽的遺傳起源推測, 二倍體洞庭青鯽亦有可能在遭遇極端環境條件和雜交因素影響下, 部分個體特化為行雌核發育方式的三倍體, 因三倍體鯽的雌核發育生殖方式比二倍體鯽兩性生殖方式具有明顯的繁殖優勢, 在人工選育的條件下, 新的洞庭青鯽三倍體種群將會越來越多, 而二倍體洞庭青鯽種群將會逐漸消失。因此,加強對洞庭湖水系二倍體野鯽和洞庭青鯽種質資源的保護已經越來越迫切, 筆者認為, 可以從以下幾方面來加強保護: (1) 建立洞庭湖水系二倍體鯽種質資源保護區, 如在洞庭青鯽的原產地澧水北民湖等水域建立洞庭青鯽省級及以上水產種質資源保護區,加強二倍體鯽種質資源管理; (2) 設置洞庭湖水系禁漁期, 加強漁業資源管理, 杜絕電魚、毒魚等毀滅性的捕撈; (3) 通過增殖放流進行洞庭青鯽和野生二倍體鯽種群的恢復; (4) 加強對三倍體雌核發育鯽如銀鯽、彭澤鯽的人工飼養管理, 以防三倍體鯽逃逸到天然水體, 導致二倍體鯽種群的減少或消失。
王蕊芳, 施立明, 賀維順, 1988. 不同地理區域鯽魚染色體銀染核仁組織者的比較研究. 動物學研究, 9(2): 165—169
文 萍, 趙 建, 李 偉等, 2015. 基于微衛星多重PCR技術的黃喉擬水龜親子鑒定. 水生生物學報, 39(6):1134—1141
朱曉東, 耿 波, 李 嬌等, 2007. 利用30個微衛星標記分析長江中下游鰱群體的遺傳多樣性. 遺傳, 29(6): 705—713
劉良國, 楊春英, 楊品紅等, 2012. 洞庭湖水系沅水和澧水野鯽的染色體組型及資源保護. 動物學雜志, 47(2):112—119
劉良國, 楊品紅, 謝春華等, 2015. 洞庭青鯽(Carassius auratus var. Dongtingking)與三個鯽品系群體遺傳多樣性的ISSR分析. 海洋與湖沼, 46(2): 426—431
劉良國, 趙 俊, 粦陳湘等, 2004. 彭澤鯽兩個雌核發育克隆的染色體組型分析. 遺傳學報, 31(8): 780—786
劉靜霞, 周 莉, 魏麗華等, 2003. 紅白錦鯉人工雌核發育純系的微衛星標記分析. 水生生物學報, 27(6): 557—562
孫效文, 楊彥豪, 魯翠云等, 2009. 銀鯽與普通鯽群體遺傳結構的比較分析. 淡水漁業, 39(4): 34—39
李建中, 劉少軍, 張軒杰等, 2005. 異源四倍體鯽鯉及其原始親本遺傳變異的微衛星標記分析. 遺傳學報, 32(4):378—383
楊品紅, 王曉艷, 吳維新等, 2007. 洞庭青鯽的染色體核型分析及品種鑒定. 淡水漁業, 37(3): 3—7
楊品紅, 吳維新, 王曉艷等, 2005. 洞庭青鯽增、養殖技術講座(一)——洞庭青鯽的生物學特性. 內陸水產, 30(3):44—45
吳 珊, 吳維新, 2006. 洞庭青鯽形態性狀遺傳分析. 激光生物學報, 15(1): 90—93
沈俊寶, 王國瑞, 范兆廷, 1983. 黑龍江主要水域鯽魚倍性及其地理分布. 水產學報, 7(2): 87—94
宋春妮, 李 健, 劉 萍等, 2011. 蟳日本4個野生群體遺傳多樣性的微衛星分析. 水產學報, 35(7): 985—991
張文靜, 余育和, 沈韞芬, 2003. 微衛星DNA遺傳分析在原生動物學中的研究進展. 水生生物學報, 27(2): 185—190
張志偉, 曹哲明, 楊 弘等, 2006. 草魚野生和養殖群體間遺傳變異的微衛星分析. 動物學研究, 27(2): 189—196
張燕萍, 陳文靜, 汪登強等, 2013. 鄱陽湖水系草魚野生及增殖放流群體遺傳多樣性分析. 江蘇農業科學, 41(9):207—211
周 盼, 張 研, 徐 鵬等, 2011. 基于26個微衛星標記的三江水系草魚遺傳多樣性分析. 中國水產科學, 18(5):1011—1020
周 莉, 劉靜霞, 桂建芳, 2001. 應用微衛星標記對雌核發育銀鯽的遺傳多樣性初探. 動物學研究, 22(4): 257—264
傅永進, 1996. 彭澤鯽的生物學性狀及養殖技術. 淡水漁業,26(2): 25—26, 34
魯雙慶, 劉 臻, 劉紅玉等, 2005. 鯽魚4群體基因組DNA遺傳多樣性及親緣關系的微衛星分析. 中國水產科學, 12(4):371—376
Bártfai R, Egedi S, Yue G H et al, 2003. Genetic analysis of two common carp broodstocks by RAPD and microsatellite markers. Aquaculture, 219(1—4): 157—167
Bessert M L, Ortí G, 2003. Microsatellite loci for paternity analysis in the fathead minnow, Pimephales promelas(Teleostei: Cyprinidae). Molecular Ecology Notes, 3(4):532—534
Botstein D, White R L, Skolnick M et al, 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. American Journal of Human Genetics,32(3): 314—331
Leberg P L, 2002. Estimating allelic richness: effects of sample size and bottlenecks. Molecular Ecology, 11(11):2445—2449
Lynch M, 1990. The similarity index and DNA fingerprinting.Molecular Biology and Evolution, 7(5): 478—484
Miyao A, Zhong H S, Monna L et al, 1996. Characterization and genetic mapping of simple sequence repeats in the rice genome. DNA Research, 3(4): 233—238
O’Connell M, Wright J M, 1997. Microsatellite DNA in fishes.Reviews in Fish Biology and Fisheries, 7(3): 331—363
Stockburge E M, Green R D, Wood W O et al, 2000.Determination of the stringency of DNA microsatellite marker genotypes for use in individual animal identification.Animal Genetics, 53: 345—348
Zhou L, Gui J F, 2002. Karyotypic diversity in polyploid gibel carp, Carassius auratus gibelio Bloch. Genetica, 115(2):223—232

- 海洋與湖沼的其它文章
- CPA-核酸試紙條快速檢測副溶血性弧菌(Vibrio parahaemolyticus)方法的建立及其在海產品檢測中的應用*
- 日本囊對蝦(Marsupenaeus japonicus)秋繁同生群仔蝦不同耐干露性能群體間的形態表型差異*
- COⅠ條形碼輔助分析雷州半島紅樹林區魚類的物種多樣性*
- 小黃魚(Pseudosciaena polyactis)形態性狀與體質量的相關性及通徑分析*
- 豆油替代魚油對赤點石斑魚(Epinephelus akaara)生長、體組成及體脂肪酸組成的影響*
- 青魚(Mylopharyngodon piceus)新發病病原類志賀鄰單胞菌(Plesiomonas shigelloides)的分離鑒定*