超富營養水平下初始堿度對銅綠微囊藻生長影響的研究
2016-01-16 01:26:27王思瑩張君枝馬文林
綠色科技 2015年2期
關鍵詞:生長
王思瑩,張君枝,馬文林
(北京應對氣候變化研究和人才培養基地,城市雨水與水環境教育部重點實驗室,北京建筑大學 環境科學與工程系,北京 100044)
超富營養水平下初始堿度對銅綠微囊藻生長影響的研究
王思瑩,張君枝,馬文林
(北京應對氣候變化研究和人才培養基地,城市雨水與水環境教育部重點實驗室,北京建筑大學 環境科學與工程系,北京 100044)
摘要:以城市河湖天然水為試驗用水,利用碳酸氫鈉作為堿度調節劑,設置了不同堿度水平,研究了氮磷營養處于超富營養水平條件下,銅綠微囊藻各相關指標受堿度的影響。結果表明:堿度條件對藻類生長有顯著影響,存在最適宜的堿度條件約為2.46mmol/L;水體中堿度的改變,會影響銅綠微囊藻對N、P的利用;堿度過高,藻類將吸收更多的營養物質來維持其生長。
關鍵詞:堿度;銅綠微囊藻;生長
收稿日期:2014-12-18
作者簡介:王思瑩(1990—),女,北京人,北京建筑大學碩士研究生。
通訊作者:馬文林(1968—),女,山西人,博士,教授,主要從事水環境生態修復技術、固體廢棄物資源化處理與利用和適應氣候變化等研究。
中圖分類號:
文獻標識碼:A
文章編號::1674-9944(2015)02-0203-04
1引言
水體藻類生長離不開碳、氮、磷3種關鍵元素,而這3種元素也是引起水體富營養化的決定因素[1]。目前人們對水體富營養研究較多的是氮磷過量進入水體導致的影響[2],而對碳源的影響研究較少[3]。
在我國,每年都有大量有機物隨著廢水、雨水徑流和垃圾等途徑進入到地表水體中,它們中大部分在水微生物作用下被不斷分解,向水中釋放CO2。此外,大氣中CO2濃度的升高,也導致從大氣向水體轉移的CO2量增加。水中CO2濃度的增加,促使水中碳酸鹽平衡向碳酸鹽增加的方向轉變,引起水體發生無機碳源增強和堿度升高這樣兩個同步的水質效應。
藻類能夠以水中無機碳作為其營養碳源。從營養源的角度看,碳酸鹽濃度與藻類生長是正相關關系。但從碳酸鹽離子強度對藻類細胞離子滲透壓角度看,藻類生長有一個適宜的碳酸鹽強度。也即,水中碳酸鹽濃度增加對藻類生長的影響,是其作為碳源和碳酸鹽離子強度兩方面綜合作用的結果。
微囊藻是一類全球性分布的藍藻,在我國大部分富營養化水體中,銅綠微囊藻(Microcystisaeruginosa)在數量和發生頻率上均占優勢[4,5]。本論文以堿度為指標表征水中碳酸鹽體系強度變化,利用小試實驗研究初始堿度變化對銅綠微囊藻生長的影響,為揭示水中無機碳源對銅綠微囊藻生長影響進行理論基礎。
2材料與方法
2.1 實驗用水
取北京展覽館后湖湖水,回到實驗室內,進行過濾、消毒處理,然后檢測水中的堿度、總氮、總磷和pH值含量,結果為pH值8.2,TN 0.672mg/L,TP 0.0083mg/L,堿度1.12mmol/L。使實驗體系的營養水平處于超富營養水平(TP、TN濃度分別≥0.2mg/L和≥2.0mg/L),通過適量添加K2HPO4和NaNO3來調節水體的總磷濃度和總氮濃度分別為2.85mg/L和0.19mg/L,然后儲存備用。
2.2 實驗方法
以湖水自然堿度(1.12mmol/L,T0組)為對照,通過向湖水中添加不同劑量的NaHCO3溶液,調整實驗水的堿度分別為1.65mmol/L、2.46mmol/L、3.34mmol/L、4.01mmol/L、4.82mmol/L,簡計為T1組、T2組、T3組、T4組和T5組,每個組各設2個平行樣。為此,取500mL 廣口錐形瓶12個,先向每個錐形瓶中各加入200mL經過預處理的天然湖水。留出1組2個錐形瓶作為對照組,不向其中添加NaHCO3溶液,其余5組10個錐形瓶作為處理組,分別向各組錐形瓶中添加不同量的NaHCO3溶液。將這些錐形瓶置于滅菌容器內121℃高溫下滅菌20min,取出放涼備用。取處于對數生長期的銅綠微囊藻純藻種,經離心、洗滌處理后,分別定量接入錐形瓶中,然后放入人工氣候箱中進行培養。培養溫度26℃,光照強度3000lux,光暗比12∶12。在光照周期內,每天搖動錐形瓶3~4次,促進營養循環,減少藻類下沉和器壁效應,并隨機交換錐形瓶位置。
在實驗過程中每天采集水樣測定實驗體系中的藻體生物量,每兩天監測一次pH值、TN、TP和總堿度等濃度,得到在不同堿度下水質變化和藻類生長情況,并且對各指標的變化曲線進行分析,以研究堿度變化對藻類生長的影響規律。
2.3 監測指標與方法
本研究的測定項目包括pH值、碳酸鹽堿度、重碳酸鹽堿度、總堿度、總磷、總氮和浮游藻類生物量。其中,pH值采用酸度計測量;藻類生物量采用血球計數板方法測量;碳酸鹽堿度、重碳酸鹽堿度、總堿度采用自動電位滴定儀測量;總氮和總磷分別采用過硫酸鉀氧化法—紫外分光光度法和鉬銻抗分光光度法測量[6,7]。
2.4 藻類比增長速率計算
比增長速率采用公式(1)進行計算:
μ=ln(xn/xn-1)/(tn-tn-1)
(1)
式中:μ為比增長速率,xn、xn-1分別為第n天與第n-1天細胞計數值,tn、tn-1分別為對應于xn、xn-1的培養時間。
3結果與討論
3.1 堿度對藻類生長的影響
3.1.1 藻類生長曲線
每日對6個堿度條件培養系統中銅綠微囊藻的藻密度值進行測定,繪制藻的生長曲線如圖1所示。
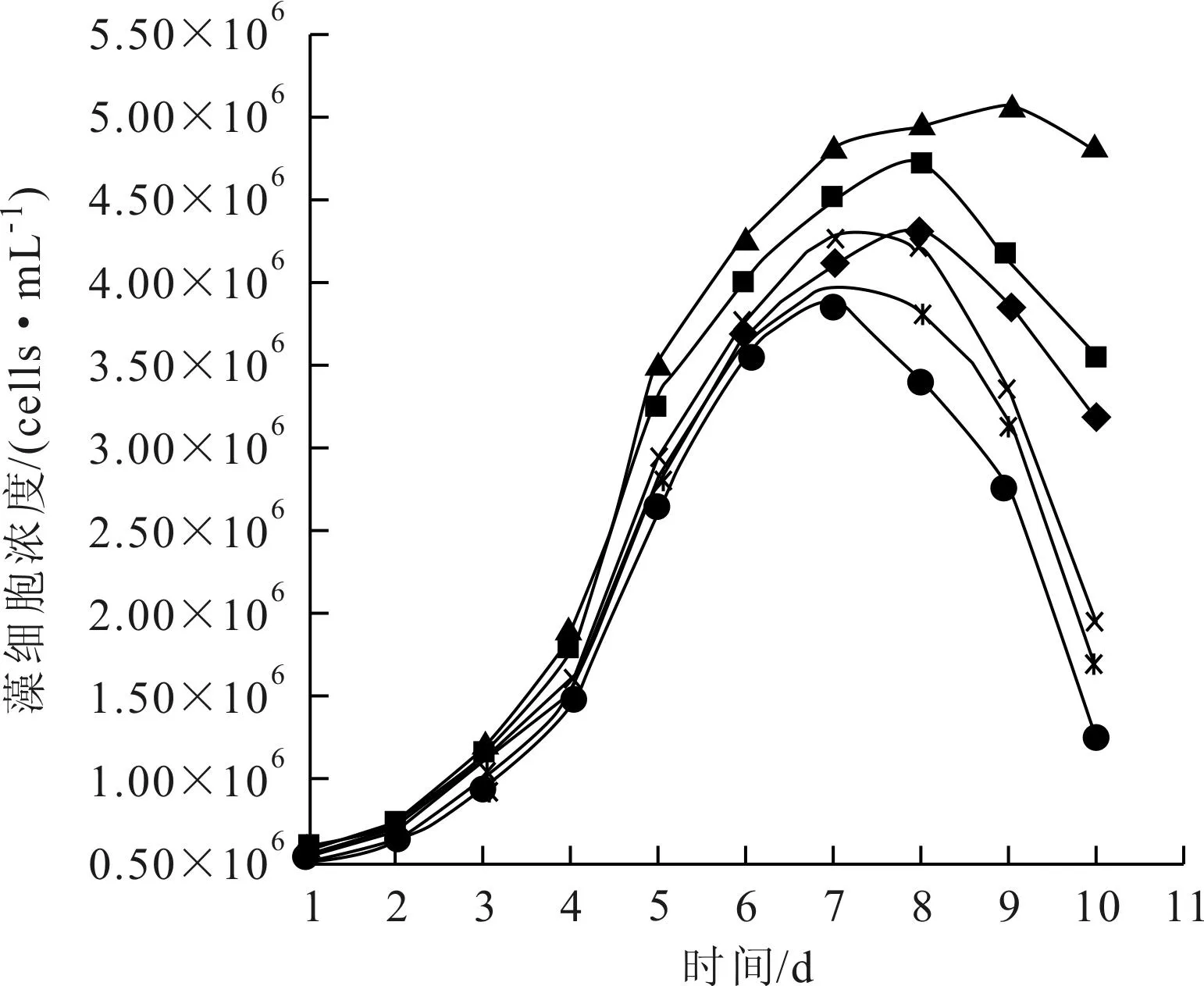
圖1 銅綠微囊藻的生長曲線(◆T0;■T1;▲T2;×T3;*T4;●T5)
從圖1看出,各組培養體系中銅綠微囊藻的生長曲線有相似的變化趨勢,都在第2天進入對數增長期,然后依次經歷減速增長期和衰減期。所不同的是,各組藻類生長的速度不一樣,培養體系中藻類生長從高向低的堿度順序分別為2.46mmol/L、1.65mmol/L、3.34mmol/L、1.12mmol/L、4.01mmol/L、4.82mmol/L。2.46mmol/L堿度條件下藻的生長速度一直高于其他各組,在達到穩定期時具有最高的生物量;其次是1.65mmol/L組;3.34mmol/L組和1.12mmol/L組,無論是生長速度還是穩定期的最高生物量,都很接近;4.01mmol/L組和4.82mmol/L組藻的生長速度和最高藻密度都比較低,而且衰亡時間也相對提前,4.82mmol/L組的最低。
因此,2.46mmol/L的堿度條件,最適合于銅綠微囊藻生長,高于或低于這個濃度,藻類的生長情況都趨于下降,且總的來說,降低堿度比提高堿度更適合藻類生長。
楊威[9]等人以不含Na2CO3的BG11培養基為基礎,,設計4組NaHCO3堿度水平培養組,培養條件與本實驗基本相同,探討堿度水平對銅綠微囊藻生長的影響。研究表明,銅綠微囊藻在不同碳酸氫鹽處理中的總體生長趨勢順序是2.4 mmol/L>4.8mmol/L>0.2mmol/L>7.3mmol/L,在實驗條件下最適宜的碳酸氫鹽堿度水平約為2.4mmol/L。與本研究得出的堿度條件影響規律相一致。
3.1.2 最大比增長率
最大比增長率是一定條件下藻類潛在增長率的最高表現[10]。根據比增長率計算公式對各種堿度條件下的銅綠微囊藻的比增長率進行連續計算,分別求出對數生長期內的最大比增長率μmax以及在對數生長期和穩定期的平均比增長率μave,列于表1中。

表1 不同堿度對藻類生長速率的影響
從表1的數據看出,在單獨培養過程中,6組藻所達到的最大比增長率,T2組的最高,T5組的最低,說明適量加入碳酸氫鹽對藻類的生長有促進作用,但添加過量時會表現出抑制作用來。按照最大比增長率排列順序依次為T2> T1> T3>T0> T4> T5組,這與穩定期時各組最高藻細胞密度是相一致的。
3.2 pH值和堿度的變化
實驗過程中,對各培養體系的pH值、總堿度、重碳酸鹽堿度以及碳酸鹽堿度進行測定,這些指標隨時間的變化情況如圖2~圖5所示。
圖2表示的是實驗過程中各個培養體系的pH值隨時間的變化情況。實驗初期,各組培養體系,由于添加NaHCO3的量不一樣,表現出不同的pH值,添加NaHCO3越多的體系,其pH值越高。而隨著實驗的進行,最后各組pH值趨于接近的水平。

圖2 pH隨時間的變化(◆T0;■T1;▲T2;×T3;*T4;●T5)
溶解于水體中的無機碳共有4種存在形式:CO2、H2CO3、HCO3-和CO32-,它們之間存在著一定的動態平衡轉化關系,其平衡方程為:CO2+H2O?H2CO3?H++HCO3-?2H++CO32-。現今越來越多的研究者發現CO2其實并不是水生植物獲取無機碳的唯一途徑,HCO3-同樣能夠作為藻類的重要無機碳源已經得到普遍認可[11,12]。影響實驗體系pH值變化的因素包含2個作用相反的方面,分別是藻類光合作用吸收利用無機碳源(CO2和HCO-3)和藻類呼吸作用釋放CO2。藻類生長過程中,利用水體中游離態CO2或HCO-3進行光合作用,平衡向左移動,H+濃度減少,故pH值增大[13]。而藻類呼吸過程釋放二氧化碳,平衡將向右移動,pH值將下降。圖2中不同實驗體系的pH值變化情況,是這兩個過程的綜合作用結果。pH值升高的階段,表明這個時期藻類生長旺盛,生長過程吸收利用二氧化碳和碳酸鹽對水中pH值的影響作用強于呼吸過程釋放二氧化碳的影響;pH值下降的階段則是相反的作用機制。對比圖2中pH值變化曲線和圖1生長曲線,二者之間有較好的相關性。T2組生長量最高,實驗結束時該組的pH值也最高。
圖3~圖5表示了各實驗體系中總堿度、碳酸鹽堿度和碳酸氫鹽堿度的變化情況,總堿度和碳酸氫鹽堿度有一致的變化趨勢,與碳酸鹽堿度變化趨勢相反,說明實驗體系的總堿度主要受碳酸氫鹽堿度的控制。
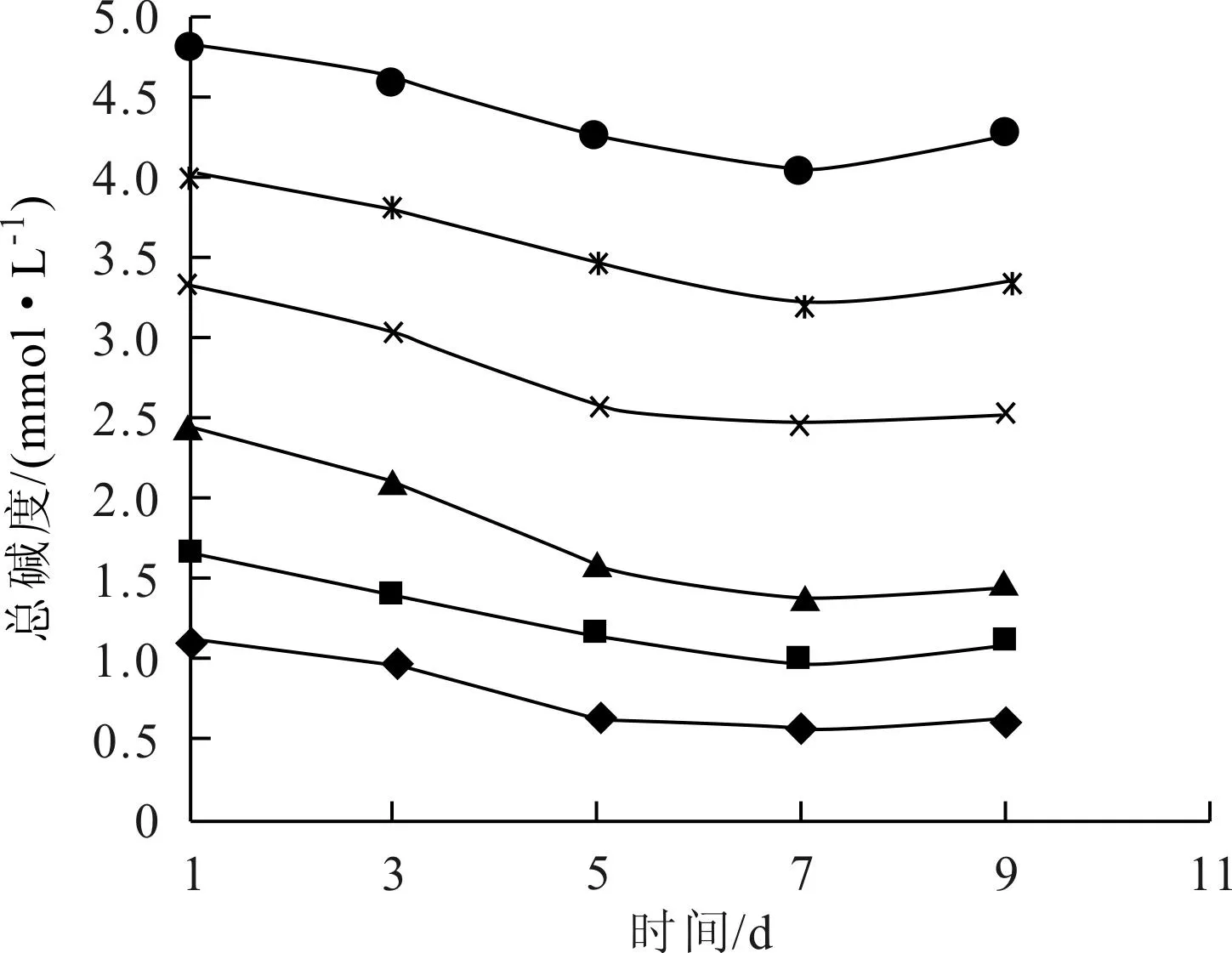
圖3 總堿度隨時間的變化(◆T0;■T1;▲T2;×T3;*T4;●T5)

圖4 碳酸鹽堿度隨時間的變化(◆T0;■T1;▲T2;×T3;*T4;●T5)
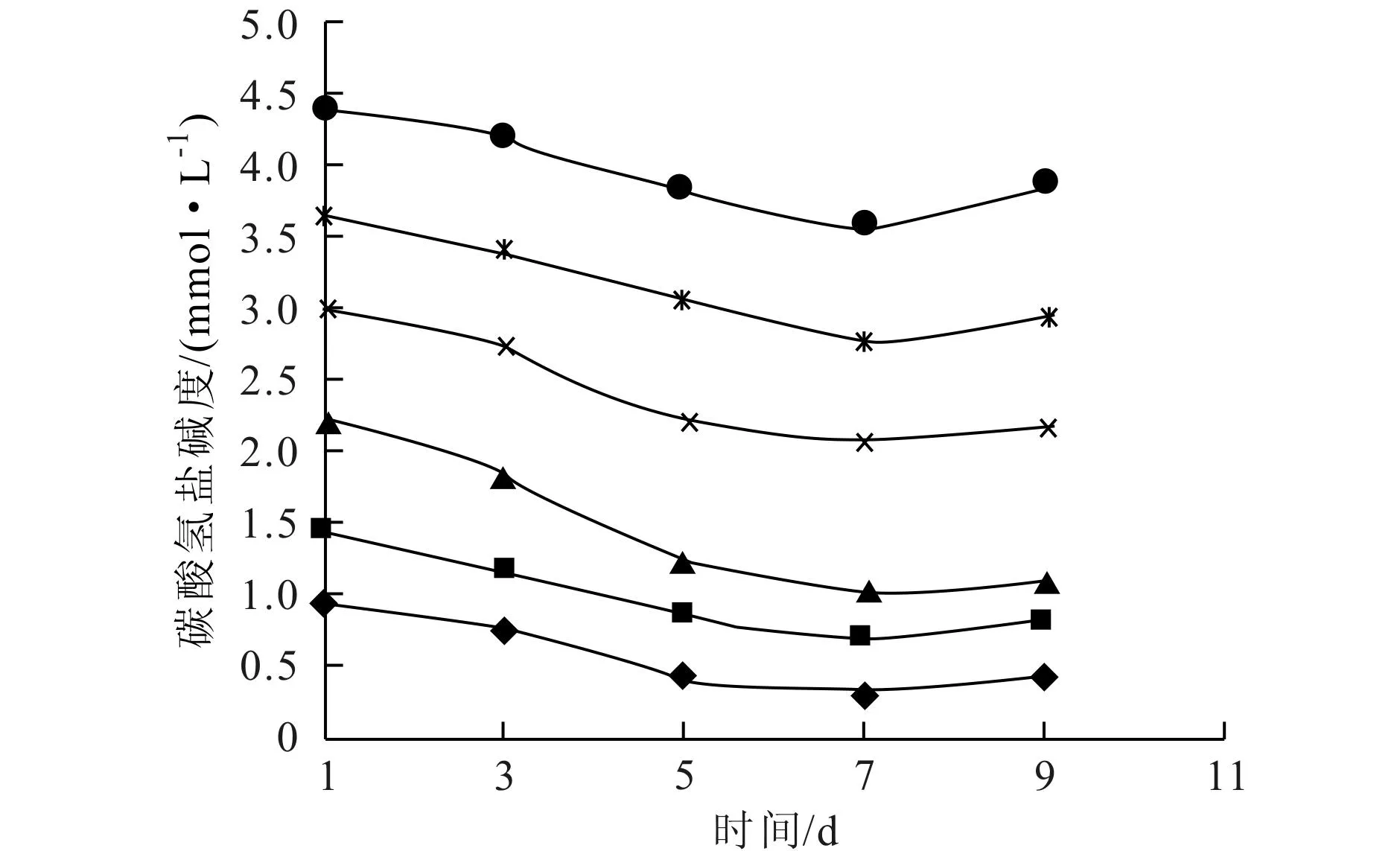
圖5 碳酸氫鹽堿度隨時間的變化(◆T0;■T1;▲T2;×T3;*T4;●T5)
藻類生長不能直接吸收碳酸鹽,只能利用水體中游離態CO2或HCO-3進行光合作用[13]。Stumm和Morgan[14]指出,淡水水體的pH值在8 左右時,碳酸氫鹽—碳占水體中總無機碳的90%左右;而pH值為9 左右時,游離CO2濃度接近于零。故在弱堿性條件下(pH值8~9) ,總無機碳中碳酸氫鹽是主要的存在形態,也是藻類能夠間接或直接利用的主要碳源形態[9]。在培養初期和中期,各體系中藻的生長量大,從水中大量吸收CO2和HCO-3,使得體系總堿度和碳酸氫鹽堿度都降低,并因此使得pH值升高,碳酸鹽平衡體系向右移動,水中碳酸鹽濃度升高。實驗后期,藻類開始先后出現衰亡跡象,生長和呼吸作用都減弱,對碳酸氫鹽的利用減少,同時在一定的pH值條件下,水中碳酸鹽體系達成新的平衡。從各圖堿度曲線看,第5天和第7天的堿度變化比較大,對應于圖1中,第3~7d銅綠微囊藻生長迅速,表明藻類生長對體系堿度變化確實存在顯著影響。
3.3 TN和TP的變化情況
羅固源等人[15]研究表明:在TN/TP為12∶1時藻類的產生周期最短,產生量最多。為使實驗用水的初始氮磷營養水平達到中富營養化程度,又能保證銅綠微囊藻的正常生長,故調節初始氮磷營養水平分別約為2.85mg/L和0.19mg/L,總氮與總磷的濃度比大約為15∶1。實驗過程中水的TN和TP變化情況如圖6和圖7所示。
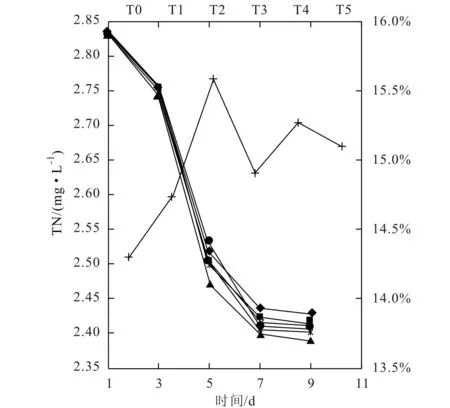
圖6 TN隨時間的變化及7天內TN的利用率(◆T0;■T1;▲T2;×T3;*T4;●T5;+利用率)
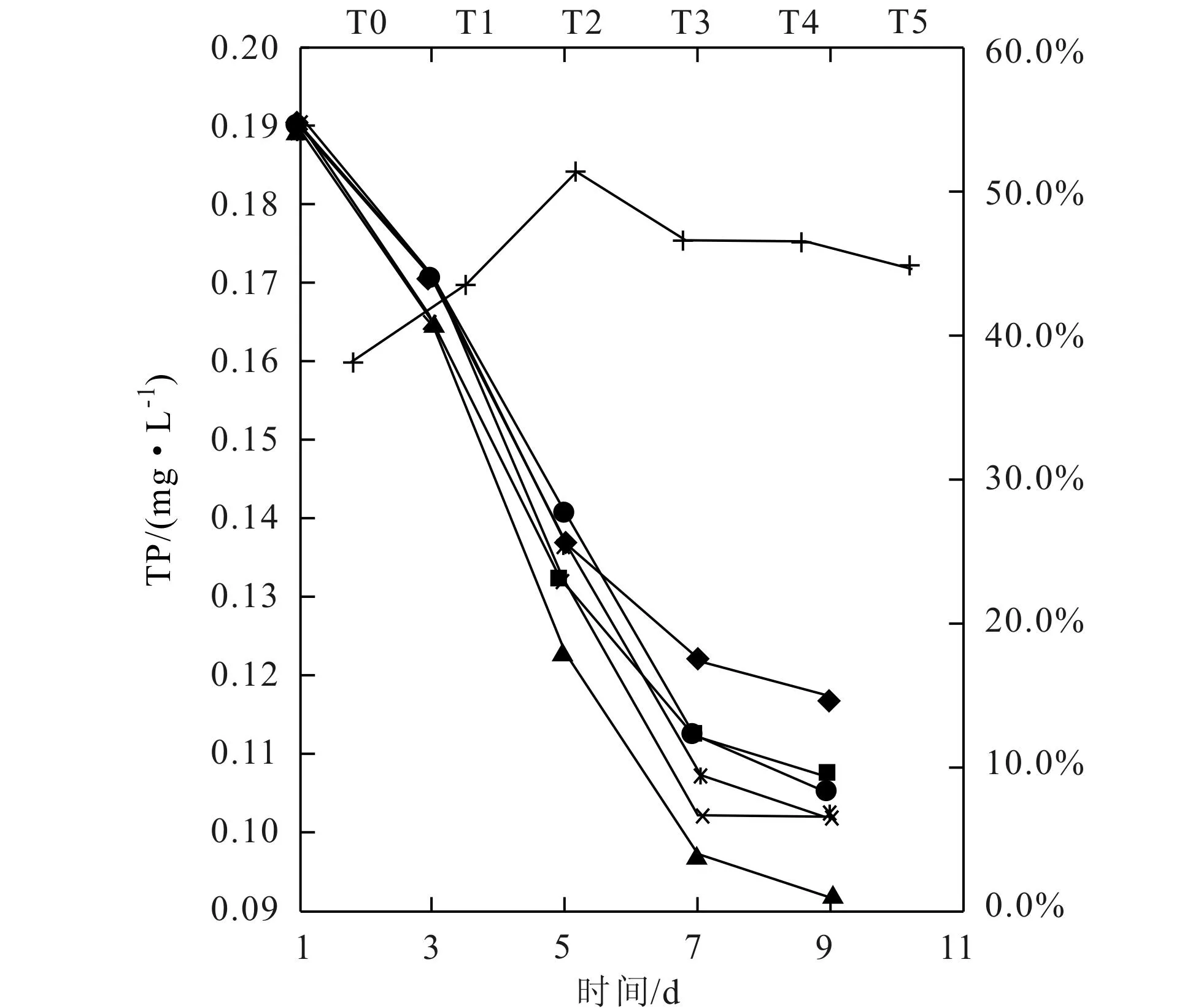
圖7 TP隨時間的變化及7天內TP的利用率(◆T0;■T1;▲T2;×T3;*T4;●T5;+利用率)
圖6表明,隨實驗的進行,各體系TN濃度普遍呈下降趨勢,在第7天趨于平衡。但不同堿度下,實驗結束時各組的TN濃度稍有差異。T2組的TN濃度一直是6個組別中最低的,表明T2組具有最大的TN利用效率。T5組的TN濃度在實驗初期處于6個組別的中等水平。
由圖7可以看出,TP的變化趨勢與TN的基本相同,但各組間的差異比TN的要顯著。銅綠微囊藻的生長主要吸收利用水中的可溶性磷PO43-,但是由于堿度的影響,使得各組水藻的長勢不同,所以對可溶性磷的利用率也不同的。實驗結束時,各組實驗體系中TP濃度由高到低的順序依次為:T0、T1、T5、T4、T3和T2,相應地各組對磷的利用率分別為:38.1%、43.4%、51.3%、46.3%、46.3%、44.7%,該順序與藻類生長曲線稍有不同,T4組和 T5組的TP的利用率相對較高,分析原因可能是過高的堿度環境,使得銅綠微囊藻在生長時消耗更多的可溶性磷。從曲線變化趨勢可以看出,TP濃度在第3~7d下降程度較大,與藻類的生長趨勢相對應。
比較藻對氮和磷的利用情況表明,銅綠微囊藻對TN的利用率比對TP的低。鄭朔方[16]等人研究表明,銅綠微囊藻對總磷的半飽和常數KSP=0.019,對總氮的半飽和常數KSN=0.111,說明銅綠微囊藻對總磷的吸收能力遠高于對總氮的吸收能力,這是該水藻能夠高效利用磷的原因所在。
4結論
(1)銅綠微囊藻在含不同堿度水平的培養液中生長,能夠間接或直接利用水中碳酸氫鹽作為光合無機碳源。藻類的生長和堿度水平存在某些相關性。
(2)初始堿度的不同會對藻類的生長產生影響,本實驗體系中最適宜的堿度條件約為2.46mmol/L,高于或低于這個濃度,藻類的生長情況都趨于下降,但總的來說,降低堿度比提高堿度更適合藻類生長。
(3)藻類生長會影響水體的酸堿水平,隨著藻類的生長,pH值逐漸升高,而水中堿度的成分也會因此相互轉換。
(4)水體中堿度的改變,會影響銅綠微囊藻對N、P的利用。各組實驗體系中對氮的利用率分別為14.3%、14.7%、15.6%、14.9%、15.3%、15.1%;對磷的利用率分別為:38.1%、43.4%、51.3%、46.3%、46.3%、44.7%。堿度過高,藻類將吸收更多的營養物質來維持其生長,N、P利用量也會相應提高。
參考文獻:
[1]鄭煥春,周 青.微生物在富營養化水體生物修復中的作用[J].中國生態農業學報,2009,17(1):197~202.
[2]馬經安,李紅清.淺談國內外江河湖庫水體富營養化狀況[J].長江流域資源與環境,2002,11(6):575~577.
[3]岳國峰,王金霞,朱明遠,等. 藻類無機碳營養的研究進展(Ⅰ)——研究起源及研究方法[J].研究綜述,2003,27(5):15~18.
[4]金相燦. 中國湖泊環境[M].北京:海洋出版社,1995.
[5]Lu Yuan, Wen Jianfan. Isolation,Pure Cultivation and Total DNA Extraction of Microcystis aeruginosa. Kütz in Dianchi Lake[J].Journal of Sciences, 2001,13(3):285~288.
[6]國家環境保護總局,水和廢水監測分析方法編委會.水和廢水監測分析方法[M].4版.北京:中國環境科學出版社,2002: 120~124.
[7]全國主要湖泊、水庫富營養化研究課題組.湖泊富營養化調查規范[M].北京:中國環境科學出版社,1987.
[8]金相燦,章宗涉.湖泊富營養化調查規范[M].北京:中國環境科學出版社,1909:257~268.
[9]楊威,孫凌,袁有才.堿度水平對銅綠微囊藻和四尾柵藻生長和競爭的影響[J].農業環境科學學報,2007,26(4):1264~1268.
[10]方濤,敖鴻毅,劉劍彤.滇池水體理化環境狀況時空分布格局研究[J].水生生物學報,2004,28(2):124~130.
[11]岳國峰,王金霞,朱明遠.藻類無機碳營養的研究進展(Ⅱ)——藻類利用無機碳的機理及其調節[J].海洋科學,2003,27(6),31~34.
[12]Jones J I,Young J,Eaton J,et al.The influence of nutrient loading, dissolved in organic carbon and higher trophic levels on the interaction between submerged plants and periphyton[J].Journal of Ecology, 2002,90(1):12~24.
[13]孫凌,闞元卿,張冬梅.無機碳對浮游藻類生長和群落結構的影響[J].環境污染與防治,2007,29(5):352~356.
[14]Stumm W, Morgan J J .水化學—天然水體化學平衡導論[M].湯鴻霄,譯.北京: 科學出版社, 1987:91~170.
[15]羅固源,康康,朱亮.水體中TN/TP與藻類產生周期及產生量的關系[J].重慶大學學報:自然科學版,2007,30(1):142~145.
[16]鄭朔方,楊蘇文,金相燦.銅綠微囊藻生長的營養動力學[J].環境科學,2005,26(2):152~156.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
