EZp對胃癌細胞核因子 κB靶基因的調節作用
2016-01-31 02:13:57吳雪雷蔡耀武莊志忠陳園靜郭仁杰鄭茂松
中國病理生理雜志 2015年12期
吳雪雷, 蔡耀武, 莊志忠, 陳園靜, 郭仁杰, 鄭茂松
(莆田學院附屬莆田市第一醫院 1腫瘤外科, 2病理科,福建 莆田 351100)
?
EZp對胃癌細胞核因子 κB靶基因的調節作用
吳雪雷1△,蔡耀武1,莊志忠1,陳園靜1,郭仁杰2,鄭茂松1
(莆田學院附屬莆田市第一醫院1腫瘤外科,2病理科,福建 莆田 351100)
[摘要]目的: 探討zeste同源物增強子2 (EZp) 蛋白的表達對胃癌細胞的影響以及可能的作用機制。方法: 用real-time PCR 和Western blot實驗檢測正常胃黏膜上皮細胞和不同胃癌細胞系中EZp的mRNA和蛋白表達;采用細胞生長、細胞遷移以及軟瓊脂增殖實驗測定EZp對胃癌細胞系致癌能力的影響;利用螢光素酶報告基因和real-time PCR檢測EZp對核因子κB靶基因的調節作用;用免疫共沉淀法檢測EZp和p65的相互作用。結果: 與正常胃上皮細胞相比,胃癌細胞系中的EZp過量表達(P<0.05)。EZp特異性抑制劑腺苷類似物DZNep處理或shEZp抑制了AGS和SNU-16細胞系的細胞活力。此外,DZNep和shEZp 抑制了AGS細胞遷移及軟瓊脂細胞形成的克隆數目(P<0.05)。shEZp 下調了核因子κB報告基因或白細胞介素8報告基因的活性以及核因子κB靶基因白細胞介素8、趨化因子配體5及趨化因子配體20的表達(P<0.05)。此外,EZp可以特異性與p65蛋白相互作用。結論: EZp 通過調節核因子κB靶基因介導胃癌細胞的生長。
[關鍵詞]胃癌; Zeste同源物增強子2; 核因子κB
流行病學研究顯示,幽門螺旋桿菌(Helicobacterpylori)感染引起的慢性炎癥與胃癌的發生有著緊密的關聯[1-2]。幽門螺旋桿菌可分泌多種毒力因子,比如細胞毒素相關基因A蛋白 (cytotoxin-associated gene A, CagA)、空泡細胞毒素(vacuolating cytotoxin A, VacA)等。而之前的數據顯示核因子κB(nuclear factor kappa B, NF-κB)介導慢性炎癥的發生。有研究表明CagA可激活核因子κB信號通路上調其下游的靶基因,包括細胞白介素(interleukin, IL)-1β、IL-6、IL-8以及腫瘤壞死因子α(tumor necrosis factor alpha, TNF-α)。而這些細胞因子可誘導炎癥的發生,從而促進腫瘤的生成[3-4]。
越來越多的研究認為表觀遺傳對于腫瘤的發生發展具有重要的調節作用。Zeste同源物增強子2(enhancer of zeste homolog 2, EZp)蛋白是多梳蛋白抑制復合體2(polycomb repressive complex 2, PRC2)中的組分之一,可特異性催化組蛋白p第27個賴氨酸三甲基化(pK27me3)。TCGA數據庫顯示EZp在胃癌組織中高表達與相關研究的結果一致[5-7]。本研究基于前人已有的信息探討胃癌細胞系中核因子κB通路與EZp的關聯,以及潛在的分子機制。
材料和方法
1材料與試劑
人胚腎細胞293T、人胃上皮細胞GES-1和人胃癌細胞AGS、 MKN-45、SNU-16、NCI-N87購自上海生博生物醫藥;DZNep是Selleck產品;EZp抗體是BD產品;T345磷酸化EZp 抗體購自Active Motif;V5和β-actin抗體、V5和HA抗體偶聯珠子、pLKO-shLuc 和shEZp(5’-ATTCTTGGTTTAAGATTTCCG-3’) 購自Sigma;HA抗體購自Santa Cruz;脂質體Lipofectamine 2000、Superscript III反轉錄酶、TRIzol、SYBR Green 購自Invitrogen;MTS試劑盒、Dual-Luciferase Reporter為Promega產品; Difco Noble 瓊脂為BD產品;TNF-α購自R&D;pCMV-HA-EZp購自Addgene;pcDNA-V5-p65 cDNA克隆自293T細胞。
2方法
2.1慢病毒包裝與感染使用磷酸鈣沉淀法轉染293T細胞。將細胞接種于100 mm培養皿中, 24 h后轉染10 μg pLKO-shLuc或者pLKO-shEZp, 8 μg pCMV-ΔR8.2和2 μg pCMV-VSV-G。轉染16 h后換成新鮮培養液。48 h后收集病毒上清并用0.45 μm 過濾器過濾并分裝冷凍于-80 ℃。AGS和SNU-16細胞培養于60 mm, 24 h后去掉培養液,加入2 mL含有5 mg/L 聚凝胺(polybrene)的慢病毒。感染12 h后,換成新鮮培養液,再過36 h后用2 mg/L的嘌呤霉素(puromycin)篩選細胞。
2.2MTS實驗檢測細胞活力將AGS和SNU-16細胞以每孔5×103個細胞接種于96孔板中,每孔加入100 μL RPMI-1640培養液。 培養24 h后換成分別含有0.25 μmol/L、 0.5 μmol/L、 1 μmol/L和 2 μmol/L DZNep的新鮮培養液,溶劑對照組加入相同體積的二甲基亞砜(dimethyl sulfoxide, DMSO)。72 h后向每孔加入20 μL MTS試劑,于培養箱中孵育2 h。然后用酶標儀測定各樣品在490 nm波長下的吸光度(A) 值,并計算相對細胞生長率。 生長率(%) =加藥孔A值/對照孔A值×100%。
2.3細胞遷移實驗AGS shLuc對照細胞和shEZp細胞以每孔5×105接種于6孔板中, 每孔加入2 mL培養液。24 h后(約有95%融合率)用20 μL tip頭用力均勻地在孔中劃痕。PBS洗滌2次,加入RPMI-1640培養液。在不同的時點測量細胞遷移的距離。細胞遷移率 (%) =不同時點劃痕距離/0 h劃痕距離×100%。
2.4軟瓊脂集落形成實驗于6孔板中加入0.6% Noble 瓊脂,于室溫冷卻40 min后,加入含0.3% Noble瓊脂的5 000個AGS shLuc對照細胞和shEZp細胞。每種細胞接種3個孔。室溫冷卻30 min后,6孔板于培養箱中培養4周后倒置顯微鏡下計數含16個細胞以上的克隆。
2.5螢光素酶報告基因活性的檢測AGS細胞以1×108/L 鋪于24孔板中。24 h后轉染每孔20 pmol的對照小干擾RNA和EZp小干擾RNA(5’-ATTCTTGGTTTAAGATTTCCG-3’)。培養24 h后換成新鮮培養液,再轉染 0.4 μg的NF-κB報告基因或IL-8報告基因,以及0.1 μg陽性對照Renilla螢光素酶報告基因。 24 h后加入含有50 μg/L TNF-α培養液孵育5 h。之后去除培養液加入50 μL PLB裂解液裂解細胞與室溫孵育20 min。每個樣品取5 μL于白色不透明96孔板中以雙熒光報告基因檢測試劑盒檢測熒光活性。各組相對熒光活性=核因子κB或IL-8報告基因熒光強度/Renilla熒光強度。
2.6實時熒光定量PCR人正常胃黏膜細胞以及胃癌細胞系以 2×108/L培養于 60 mm 培養皿中。24 h后去掉培養基,加入1 mL TRIzol裂解提取總RNA。 1 μg總RNA用SuperScript III反轉錄酶合成cDNA。20 μL的反應體系含10 μL 2×SYBR Green, 2 μL 引物,1 μL cDNA以及7 μL 去離子水。PCR反應條件為95 ℃ 20 s; 95 ℃ 3 s, 60 ℃ 30 s,擴增40個循環。所有PCR反應均在ABI 7500 Fast Real-Time PCR System上進行。或者,AGS細胞鋪板24 h后,加入50 μg/L TNF-α,分別于不同的時點收集細胞。TRIzol裂解提取總RNA。1 μg總RNA用SuperScript III反轉錄酶合成cDNA。EZp的上游引物序列為5’-AATCAGAGTACATGCGACTGAGA-3’, 下游引物序列為5’-GCTGTATCCTTCGCTGTTTCC-3’;IL-8的上游引物序列為5’-GACCACACTGCGCCAACAC-3’, 下游引物序列為5’-CTTCTCCACAACCCTCTGCAC-3’; CXCL5的上游引物序列為5’-AGCTGCGTTGCGTTTGTTTAC-3’,下游引物序列為5’-TGGCGAACACTTGCAGATTAC-3’;CCL20的上游引物序列為5’-CTGGCTGCTTTGATGTCAGT-3’, 下游引物序列為5’-CGTGTGAAGCCCACAATAAA-3’;GAPDH的上游引物序列為5’-CTGGGCTACACTGAGCACC-3’, 下游引物序列為5’-AAGTGGTCGTTGAGGGCAATG-3’。
2.7免疫共沉淀和Western blot實驗293T細胞鋪板于6 cm培養皿24 h后,共轉染4 μg pCMV-HA-EZp和/(或) 4 μg pcDNA3-V5-p65,用空載體pc-DNA3平衡總DNA量。36 h后,收集細胞并用RIPA裂解液裂解。收集上清,加入10 μL V5或者HA抗體偶聯的珠子于4 ℃孵育2 h。1 000 r/min離心 1 min, 用RIPA裂解液洗3次,去上清并加入50 μL 2×樣品液于95 ℃煮5 min。收集上清用10%聚丙烯酰胺凝膠電泳 (SDS-PAGE)分離。接著電轉至硝酸纖維素膜,室溫用5%脫脂牛奶封閉1 h,加入I抗4 ℃孵育過夜,用含0.1% 吐溫 20 的TBS緩沖液漂洗3次,每次5 min。加入相應辣根過氧化物酶(horseradish peroxidase,HRP)標記的 II 抗室溫孵育1 h后,化學發光試劑孵育 5 min,暗室曝光顯影。β-actin作為內參照。
3統計學處理
用SPSS 16.0統計軟件進行分析。數據均采用均數±標準差(mean±SD)表示。顯著性差異檢驗采用單因素方差分析及Bonferroni校正的t檢驗,以P<0.05為差異有統計學意義。
結果
1胃癌細胞系中EZp mRNA和蛋白的表達
我們首先比較了4種不同的人胃癌細胞系與正常胃黏膜上皮細胞中EZp的mRNA表達水平,4種胃癌細胞EZp的mRNA表達水平均比正常細胞高(P<0.01)。同樣通過Western blot檢測EZp總蛋白水平,胃癌細胞中的表達高于正常細胞(P<0.05),同時檢測EZp T345磷酸化發現其表達水平與總蛋白呈正相關,且正常胃上皮細胞的磷酸化EZp表達較弱(P<0.05),見圖1。以上結果說明EZp可能對胃癌細胞的生長以及致癌能力有調節作用。由于AGS和SNU-16中的表達水平相對較高,后續的實驗均用其作為研究對象。
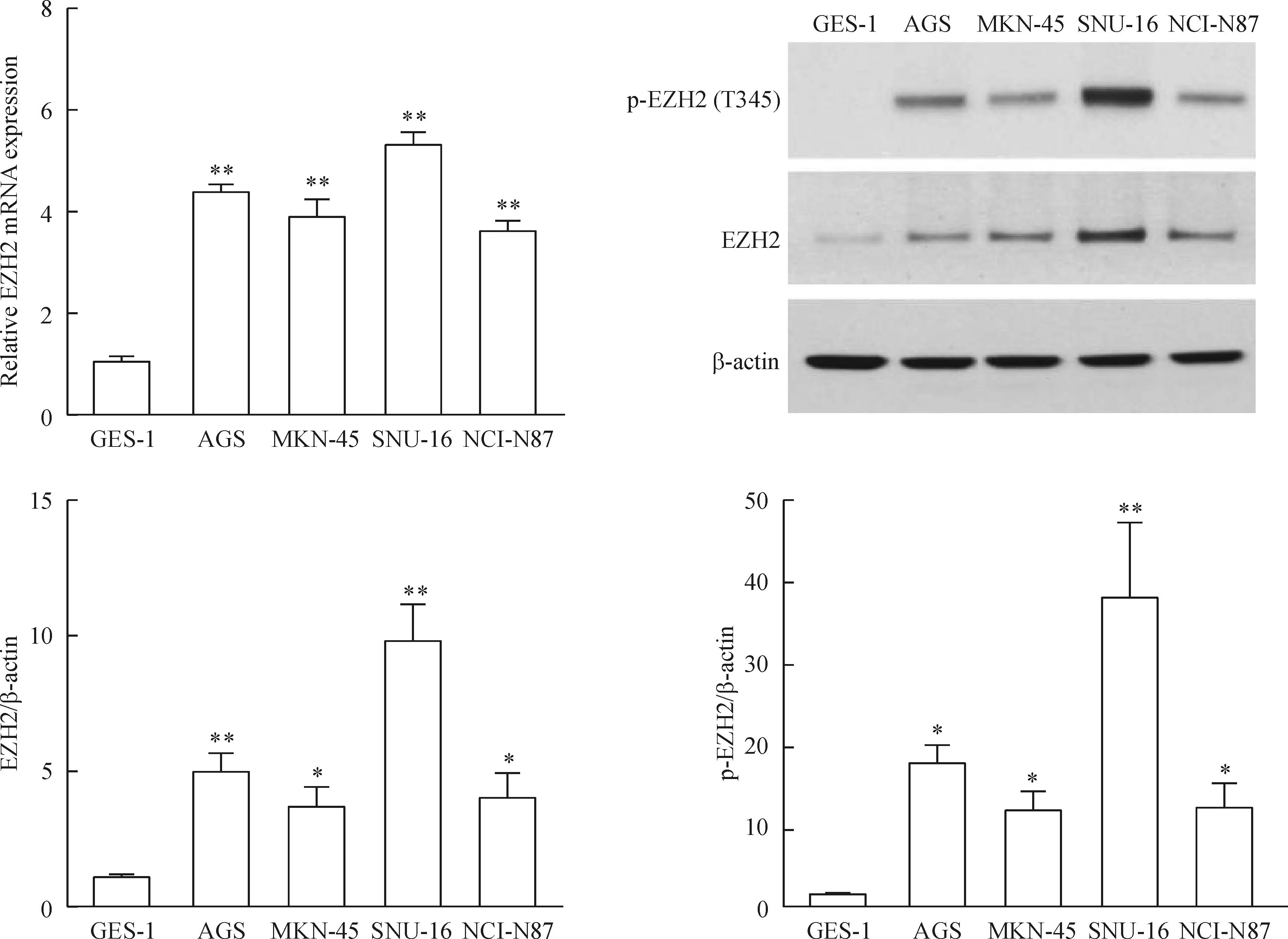
Figure 1.The expression of EZp in normal gastric epithelial cells and different gastric cancer cell lines. Mean±SD.n=3.*P<0.05,**P<0.01vsGES-1.
圖1正常人胃上皮細胞與不同胃癌細胞EZp mRNA和蛋白的表達水平
2抑制EZp或沉默EZH2對胃癌細胞生長的抑制作用
胃癌細胞AGS和SNU-16接種于96孔板后,以不同濃度DZNep處理72 h, MTS實驗檢測EZp抑制劑對細胞活力的影響。結果顯示,DZNep抑制細胞的生長呈劑量效應關系。當劑量達到2 μmol/L時,對AGS和SNU-16細胞的抑制率分別為(46.62±3.47)%和(43.78±2.58)%(P<0.01)。Western blot證明了DZNep對 EZp蛋白表達的抑制呈濃度依賴性關系。類似地, 利用慢病毒將EZH2 shRNA導入2種胃癌細胞,puromycin篩選,獲得穩定表達shEZp的細胞株,Western blot檢測到EZp表達水平降低。shRNA介導的EZp沉默抑制了胃癌細胞的增殖,見圖2。

Figure2.TheeffectsofEZH2inhibitororEZpsilencingoncellactivity.A:inhibitoryeffectofDZNeponAGSandSNU-16cellactivity;B:DZNepdown-regulatedEZH2proteinexpressioninadose-dependentmanner;C:knockdownofEZpinAGSandSNU-16cells;D:knockdownofEZpdecreasedtheactivityofAGSandSNU-16cells.Mean±SD. n=3.*P<0.05,**P<0.01vs0 μmol/L;#P<0.05 vsshLuc.
圖2EZH2抑制劑或EZp沉默對胃癌細胞增殖的影響
3抑制EZH2或沉默EZp對細胞遷移和集落形成的影響
以穩定表達對照shRNA和EZpshRNA的AGS細胞為對象做細胞劃痕實驗。劃痕后18h,對照組細胞的遷移率為(79.21±3.26)%, 而EZpshRNA細胞的遷移率為(47.85±4.89)%,二者之間差異有統計學意義(P<0.01)。36h后,對照組細胞劃痕完全消失即達到100%的遷移率,但是EZp 沉默細胞為81.54%,差異有統計學意義(P<0.01)。在軟瓊脂集落形成實驗中,對照組細胞形成較多的細胞克隆,克隆體積較大,而EZpshRNA細胞形成的細胞克隆顯著減少且沒有較大的克隆體積(P<0.05)。同樣,以EZH2抑制劑DZNep處理的細胞形成的細胞集落不管是數量還是體積均比DMSO處理細胞要少而小(P<0.05),見圖3。這說明EZH2增強AGS細胞的集落形成能力,使癌細胞的惡性度降低。

Figure3.SilencingofEZporinhibitionofEZH2reducedcellmigrationandcolonyformation(×40).A:shEZH2inhibitedAGScellmigrationinscratchassay;B:shEZH2AGScellsshowedlessandsmallercoloniesinanchorage-independentassay;C:DZNepreducedcolonynumberandsize.Mean±SD. n=3.*P<0.05,**P<0.01vsshLuc or DMSO.
圖3 EZp 沉默或EZH2 抑制劑對細胞遷移和集落形成的影響
4EZH2對NF-κB靶基因的影響
分別利用不同的螢光素酶報告基因NF-κB-Luc和IL-8-Luc,我們發現 EZp 沉默抑制了TNF-α介導的啟動子激活作用(P<0.01)。另外,通過實時螢光定量PCR法檢測NF-κB靶基因IL-8、CXCL5和CCL20的mRNA表達,發現在TNF-α處理4h后,與對照組相比,EZpshRNA下調了上述3個基因的mRNA表達(P<0.05)。以上結果顯示EZH2對TNF-α誘導的胃癌細胞中核因子κB靶基因具有正調節作用,見圖4。
5EZH2與NF-κBp65的相互作用
利用免疫共沉淀驗證EZH2與p65在哺乳動物細胞中是否相互作用。結果顯示,在V5-p65的沉淀產物中可以檢測到HA-EZH2,而在對照組中則檢測不到。同樣,在HA-EZH2的沉淀產物中可以檢測到V5-p65,但是對照組沒有檢測到蛋白,見圖5。這表明V5-p65與HA-EZH2在細胞中具有特異性的蛋白相互作用。
討論
越來越多的研究證實了幽門螺旋桿菌對于胃癌的發生起到誘因作用。幽門螺旋桿菌通過其自身分泌的毒力因子誘使慢性炎癥的形成。已有的證據認為核因子κB對胃癌具有促進作用[4]。然而,核因子κB是如何參與調節炎癥及惡性腫瘤發生的分子機制仍然不清楚。有報道表明核因子κB通過調節其下游的靶基因促進炎癥發生和腫瘤的生成[8-10]。
近年來表觀遺傳學在腫瘤的發生和發展中的調節作用受到了越來越多的關注。EZH2是PRC2復合體中的甲基轉移酶催化活性成分,在許多不同類型的腫瘤中過表達[11]。在胃癌中,EZH2通過抑制抑癌微小RNAs(microRNAs),比如miR-1246、miR-302a、miR-4448、miR-200和miR-429,或者下調RUNX3轉錄因子的表達,增強致癌能力[12-14]。此外,EZH2的翻譯后修飾也參與調解其活性[15]。
基于前人的研究,本文主要探討EZH2與核因子κB轉錄因子之間潛在的關系。我們在一系列的胃癌細胞中證實了EZH2總蛋白和蛋白磷酸化的過表達, 而正常胃上皮細胞的表達水平較弱。不同的研究說明EZH2蘇氨酸345和487磷酸化對腫瘤細胞的增殖和遷移具有促進作用[15]。EZpshRNA沉默抑制了癌細胞的生長、遷移和致癌力。同樣,小分子抑制劑DZNep處理胃癌細胞24h, 誘導濃度依賴性的EZH2降解。DZNep抑制細胞增殖和集落形成。TNF-α可誘導與炎癥相關的基因包括細胞因子和趨化因子的表達,而核因子κB是關聯TNF-α信號通路與下游基因的媒介物。有研究表明核因子κB的靶基因IL-8、CXCL5以及 CCL20可促進局部炎癥發生,細胞異常增殖,最終導致腫瘤的形成、遷移和侵襲。這些細胞因子一方面可招募炎癥細胞到上皮細胞周圍,而炎癥細胞分泌的其它細胞因子可促進上皮細胞的生長,另一方面可直接激活上皮細胞內促生存的相關信號通路[4,16-17]。本研究顯示,抑制EZp的表達顯著下調了核因子κB的轉錄活性以及上述細胞因子和趨化因子的表達水平。之前的報道聲稱在三陰性乳腺癌細胞中EZH2特異性地與p65相互作用并促進p65結合到靶基因啟動子比如IL-6、IL-8以及TNF-α,增強細胞因子的表達從而促進乳腺癌細胞的增殖[18]。本研究發現IL-8、CXCL5和CCL20同樣能被EZH2調控。通過蛋白質-蛋白質相互作用的研究,我們發現過表達的EZH2和核因子κB組分p65相互作用。EZH2可能是通過相似的機制達到調節作用。EZH2是組蛋白甲基轉移酶,具有催化活性。Lee等[18]的數據顯示在特定乳腺癌細胞中EZH2對核因子κB靶基因的調節不依賴于其酶活性。在胃癌細胞中,我們不能排除EZH2可能通過其酶活性調節核因子κB下游基因的表達。將來需要進一步的研究以證實這種可能性。本研究結果即EZH2介導的胃癌中核因子κB靶基因調控為闡明慢性炎癥促進胃癌發生以及潛在的靶向治療提供了實驗依據。

Figure4.SilencingofEZpinhibitedTNF-α-mediatedNF-κBtranscriptionalactivityanddownstreamtargetgeneexpression.A: EZpsilencingdecreasedTNF-α-inducedNF-κB-LucandIL-8-Lucactivity;B: EZpsilencingsuppressedNF-κBtargetgeneexpressioninducedbyTNF-α.Mean±SD. n=3.*P<0.05,**P<0.01vsscrambled ;##P<0.01vsscrambled + TNF-α.
圖4EZp沉默對TNF-α誘導的核因子κB的轉錄活性及其靶基因的抑制作用

Figure5.Over-expressedEZH2physicallyintereactedwithNF-κBp65inHEK293Tcells.A:interactionbetweenHA-EZH2andV5-p65detectedbyimmunoprecipitionwithV5antibody;B:bindingofV5-p65toHA-EZH2byimmunoprecipitionwithHAantibody.
圖5過表達的EZH2與NF-κBp65的相互作用
[參考文獻]
[1]WarrenJR.GastricpathologyassociatedwithHelicobacter pylori[J].GastroenterolClinNorthAm, 2000, 29(3):705-751.
[2]ChibaT,MarusawaH,UshijimaT.Inflammation-associatedcancerdevelopmentindigestiveorgans:mechanismsandrolesforgeneticandepigeneticmodulation[J].Gastroenterology, 2012, 143(3):550-563.
[3]EL-OmarEM,CarringtonM,ChowWH,etal.Interleukin-1polymorphismsassociatedwithincreasedriskofgastriccancer[J].Nature, 2000, 404(6776):398-402.
[4]Ben-NeriahY,KarinM.Inflammationmeetscancer,withNF-κBasthematchmaker[J].NatImmunol, 2011, 12(8):715-723.
[5]MatsukawaY,SembaS,KatoH,etal.Expressionoftheenhancerofzestehomolog2iscorrelatedwithpoorprognosisinhumangastriccancer[J].CancerSci, 2006, 97(6):484-491.
[6]ChoiJH,SongYS,YoonJS,etal.Enhancerofzestehomolog2expressionisassociatedwithtumorcellproliferationandmetastasisingastriccancer[J].APMIS, 2010, 118(3):196-202.
[7]CaiGH,WangK,MiaoQ,etal.ExpressionofpolycombproteinEZH2inmulti-stagetissuesofgastriccarcinogenesis[J].JDigDis, 2010, 11(2):88-93.
[8]MatsumotoY,MarsawaH,KnoshitaK,etal. Helicobac-ter pyloriinfectiontriggersaberrantexpressionofactivation-inducedcytidinedeaminaseingastricepithelium[J].Nat-Med,2007, 13(4):470-476.
[9]LambA,ChenLF.RoleoftheHelicobacter pylori-inducedinflammatoryresponseinthedevelopmentofgastriccancer[J].JCellBiochem, 2013, 114(3):491-497.
[10]HorYT,VoonDC,KooJK,etal.AroleofRUNX3ininflammation-inducedexpressionofIL23Aingastricepithelialcells[J].CellRep, 2014, 8(1):50-58.
[11]ChaseA,CrossNC.AberrationsofEZpincancer[J].ClinCancerRes, 2011, 17(9): 2613-2618.
[12]HibinoS,SaitoY,MuramatsuT,etal.Inhibitorsofenhancerofzestehomolog2 (EZH2)activatetumor-suppressormicroRNAsinhumancancercells[J].Oncogenesis, 2014, 3:e104.
[13]FujiiS,ItoK,ItoY,etal.Enhancerofzestehomologue2 (EZH2)down-regulatesRUNX3byincreasinghistoneH3methylaiton[J].JBiolChem, 2008, 283(25):17324-17332.
[14]NingX,ShiZ,LiuX,etal.DNMT1andEZH2mediatedmethylationsilencesthemicroRNA-200b/a/429geneandpromotestumorprogression[J].CancerLett, 2015, 359(2): 198-205.
[15]YamaguchiH,HungMC.RegulationandroleofEZH2incancer[J].CancerResTreat, 2014,46(3):209-222.
[16]SouttoM,BelkhiriA,PiazueloMB,etal.LossofTFF1isassociatedwithactivationofNF-κB-mediatedinflammationandgastricneoplasiainmiceandhumans[J].JClinInvest, 2011, 121(5): 1753-1767.
[17]HarantH,EldershawSA,LindleyIJ.Humanmacrophageinflammatoryprotein-3α/CCL20/LARC/Exodus/SCYA20istranscriptionallyupregulatedbytumornecrosisfactor-αviaanon-standardNF-kBsites[J].FEBSLett, 2001, 509(3):439-445.
[18]LeeST,LiZ,WuZ,etal.Context-specificregulationofNF-κBtargetgeneexpressionbyEZH2inbreastcancers[J].MolCell, 2011, 43(5):798-810.
(責任編輯: 林白霜, 羅森)

*[基金項目]國家自然科學基金資助項目(No. 81370706;No. 81372758);重慶市自然科學基金資助項目(No.cstc2013jcyjA10058);重慶市人力資源與社會保障局課題資助項目(渝留助2013011號);重慶市衛生局課題資助項目(No. 2012-2-001)
EZp-mediated regulation of NF-κB target gene expression in gastric cancerWU Xue-lei1, CAI Yao-wu1, ZHUANG Zhi-zhong1, CHEN Yuan-jing1, GUO Ren-jie2, ZHENG Mao-song1
(1DepartmentofOncology,2DepartmentofPathology,TheFirstAffiliatedHospitalofPutianCity,PutianUniversity,Putian351100,China.E-mail:xuelei_wu@163.com)
[ABSTRACT]AIM: To explore the mechanism by which over-expression of enhancer of zeste homolog 2 (EZp) in a panel of gastric cancer cell lines is involved in tumorigenesis of gastric cancer. METHODS: Real-time PCR and Western blot were employed to examine the mRNA and protein levels of EZp, respectively. MTS assay, cell migration and soft agar assay were performed to investigate the role of EZp in the regulation of stomach cancer behaviors. The effect of EZp on NF-κB target gene expression was determined by Luciferase reporter and real-time PCR. Co-immunoprecipitation was used to analyze the interaction of EZp and p65 in HEK293T cells. RESULTS: The expression levels of EZp were significantly increased in the gastric cancer cells compared with normal gastric epithelial cells. Pharmacological inhibition by DZNep or knockdown of EZH2 significantly compromised AGS and SNU-16 cell activity, cell migration and anchorage-independent cell growth. Moreover, siRNA knockdown of EZH2 impaired NF-κB downstream targets, such as IL-8, CXCL5 and CCL20. In addition, the interaction of EZp and p65 was detected. CONCLUSION: EZp mediates the growth of gastric cancer cells through the regulation of NF-κB downstream gene expression.
[KEY WORDS]Gastric cancer; Enhancer of zeste homolog 2; Nuclear factor-κB
通訊作者△Tel: 023-89012559; E-mail: gouxincq@163.com
[收稿日期]2015- 05- 07[修回日期] 2015- 09- 21
[文章編號]1000- 4718(2015)12- 2176- 07
doi:10.3969/j.issn.1000- 4718.2015.12.010
[中圖分類號]R730.23.12
[文獻標志碼]A
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
