雷竹克隆系統(tǒng)出筍期有機碳分布變化規(guī)律
2016-04-14 03:02:21冀琳珂謝錦忠
生態(tài)學報 2016年23期
冀琳珂,謝錦忠,張 瑋,盧 鵬,張 磊
中國林業(yè)科學研究院亞熱帶林業(yè)研究所,杭州 311400
雷竹克隆系統(tǒng)出筍期有機碳分布變化規(guī)律
冀琳珂,謝錦忠*,張 瑋,盧 鵬,張 磊
中國林業(yè)科學研究院亞熱帶林業(yè)研究所,杭州 311400
植物光合碳同化物為植物的生長提供所需的物質和能量。目前,對植物有機碳分布雖已展開了大量研究,但對竹類植物出筍期有機碳轉移機制的研究還比較缺乏。測定了分株數(shù)量為單株、雙株和三株的雷竹克隆系統(tǒng)出筍期分株各器官的有機碳含量,以期進一步了解雷竹克隆系統(tǒng)出筍期有機碳轉移變化規(guī)律。研究發(fā)現(xiàn):雷竹分株不同器官有機碳含量差異顯著,且在出筍期發(fā)生顯著變化,出筍前:枝(52.64%)>葉(47.18%)>稈(40.98%)>鞭(40.13%)>根(35.14%),出筍完成后:枝(48.20%)>稈(47.84%)>葉(45.53%)>鞭(45.52%)>根(44.29%),枝、葉有機碳含量呈先下降后上升趨勢,根、稈、鞭有機碳含量呈“N”型變化規(guī)律;單株、多株系統(tǒng)雷竹分株各器官有機碳含量降幅與出筍量成反比,隨分株數(shù)量增加,出筍量增加,而各器官有機碳含量降幅減小;雙株系統(tǒng)中1年生雷竹各器官有機碳含量降幅大于2年生竹,三株系統(tǒng)中3年生雷竹各器官有機碳含量降幅大于1年、2年生竹。這些結果表明:出筍影響雷竹各器官有機碳分配格局,出筍時各器官間有機碳資源發(fā)生轉移,其中枝、葉有機碳含量降低而根、稈、鞭有機碳含量增加;各器官間源-匯關系發(fā)生變化,分株間有機碳資源存在共享,分株數(shù)量增加出筍量增加且系統(tǒng)內分株的損耗減小;分株年齡是影響雷竹不同器官出筍期有機碳含量變化的影響因素之一。因此,調整雷竹林年齡結構對提高雷竹林出筍量及經(jīng)濟效益有十分重要的現(xiàn)實意義。
雷竹;克隆系統(tǒng);出筍期;有機碳分配格局
碳是重要的生命元素,是植物光合作用形成有機質不可或缺的元素之一。葉片是植物進行光合作用的主要場所(源),根、種子、果實等是消耗或貯存同化物的器官(匯),同化物在源-匯間的運輸和再分配是植物生存、生長、繁殖的物質基礎[1]。植物年齡不同、發(fā)育階段不同,不同功能器官的源-匯關系會發(fā)生改變,碳素資源的分配格局可能會不同[2]。不同植物、不同器官、植物的木質組成、各器官相對位置以及氣候、溫濕度、地區(qū)差異、立地條件、年齡等均會影響植物內部有機碳分配格局[3- 5],近年來的研究表明:溫帶樹種的含碳率范圍為47.1%—51.4%[6],思茅松(PinusKesiyavar.langbianensis)人工中、幼齡林樹干的含碳率顯著高于其他器官,且由基部向上呈下降的趨勢[7],人工紅松(Pinuskoraiensis)林單株而言枝含碳率高于根含碳率[8]。
克隆植物在源-匯關系驅動下物質或資源可以通過連接物或間隔子在克隆分株間進行運輸與分享[9- 10],因此,克隆植物各分株不但在垂直方向存在碳素源-匯關系,同時在水平方向相連的分株間也存在源-匯關系[11- 14]。克隆植物具有克隆整合、克隆可塑性及克隆繁殖等獨特的生長、發(fā)育、繁殖方式[15],并且通過生理整合能夠對所獲取資源進行再分配[16],因此克隆分株間的碳素分布、整合格局及對不同程度的環(huán)境因子(如溫、光、水、肥等)的各種響應機制及表征方式成為克隆植物生態(tài)學集中研究的重點問題[17- 18]。近年來對克隆植物的碳素分布及整合研究表明:毛竹(Phyllostachysedulis)各器官含碳率:枝>稈>葉>鞭>根[19];苦竹(Pleiobtastusamarus)各器官含碳率:稈>枝>葉>鞭>根[20];羊草(Leymuschinensis)的某些分株經(jīng)受遮光和去葉干擾處理時,未被干擾的分株都能向干擾分株轉移碳物質;對羊柴(Hedysarumlaeve)分株進行去葉處理時發(fā)現(xiàn),去葉子株從未去葉母株獲得的同化物相對于對照的未去葉分株有所增加[21];對結縷草(ZoysiajaponicaSteud)匍匐莖進行連接與切斷對比試驗,發(fā)現(xiàn)分株數(shù)量對其形態(tài)學指標均有顯著促進作用[22],分株年齡是決定處于生長發(fā)育早期的歐亞活血丹(Glechomalongituba)分株間碳素源-匯關系的主要因素[23];聚花過路黃(Lysimachiacongestiflora)的分株種群密度及生物量隨光照增強顯著增加;局部遮光可以改變林生山黧豆(Lathyrussylvestris)光合產(chǎn)物的整合格局[24];模擬增溫研究發(fā)現(xiàn)草本克隆植物地上生物量及葉片干質量都隨著溫度的升高而增加[25];在一定范圍內隨著水分的增加,草本克隆植物種群的葉片生物量占整個地上生物量的百分比顯著增加[26]。
竹子是典型的木本克隆植物,具有很強的生理整合作用和異質環(huán)境適應能力,竹子通過生理整合作用即可通過相連分株間同化物、養(yǎng)分的傳輸對所獲得的碳素資源進行再分配,從而提高整個系統(tǒng)的適合度。 目前關于植物有機碳分配格局的研究主要集中于針、闊葉喬、灌木樹種[27]及草本型克隆植物,對竹類木本克隆植物有機碳的研究較少,且主要集中于單株或林分碳儲量的研究,包括基于不同的模型比較和估計竹林的碳儲量及不同類型森林系統(tǒng)中竹類植物碳儲量的變化[28- 31],以及非結構性碳及地上生物量分配的研究[19,32],對于筍期克隆系統(tǒng)間碳素傳導及源-匯關系變化引起的碳流動研究尚不多見。雷竹(Phyllostachysviolascens)是我國竹類資源中重要的筍用竹種,屬中、小型散生竹,出筍早、筍期長、筍味鮮美、產(chǎn)量高、經(jīng)濟效益好,是浙江省筍用竹主產(chǎn)區(qū)主栽竹種之一。雷竹出筍生長迅速,從出筍到幼竹形成僅需40—60 d,研究筍-幼竹快速生長期的有機碳分布變化、轉移機制,對于竹子的生存、生長、繁殖擴張和資源利用等具有重要的生態(tài)學意義,對調整林分結構、合理培育竹林具有重要的現(xiàn)實意義。因此,本文以切斷試驗法[33]切鞭,濕燒法[17]測定出筍不同時期雷竹各器官有機碳含量,旨在探討:雷竹不同器官有機碳分配格局及源-匯關系驅動下不同分株數(shù)的雷竹克隆系統(tǒng)出筍期碳素分配轉移規(guī)律,以期為雷竹林的合理經(jīng)營提供一定的理論基礎。
1 材料與方法
1.1 研究區(qū)概況
試驗地區(qū)位于浙江省杭州市富陽區(qū)春建鄉(xiāng)云二村(119°57′E,30°03′N)。氣候屬亞熱帶季風氣候,四季分明,降水充沛,光照充足,溫和濕潤,氣候垂直變化明顯。年均溫度16.3 ℃,年均降水量1479.3 mm,年日照時數(shù)1759.2 h,無霜期約238 d。
1.2 研究方法
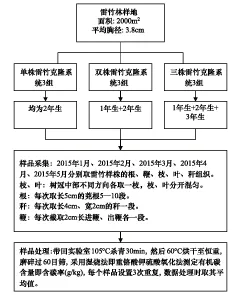
圖1 試驗研究方法Fig.1 Study method of experiment
2015年1月,在試驗區(qū)選擇生長良好的雷竹林地,在樣地內選擇地下鞭系相連的雷竹克隆系統(tǒng)進行切鞭處理,選取分株數(shù)量分別為單株、雙株和三株的克隆系統(tǒng)各3組,切鞭時進鞭、出鞭各保留50 cm以上, 鞭全長為2 m左右,單株克隆系統(tǒng)雷竹年齡均為2年生,雙株克隆系統(tǒng)為1年生、2年生雷竹各1株,三株克隆系統(tǒng)為1年生、2年生、3年生雷竹各1株。所選雷竹生長健壯,稈通直圓滿,無疤結、無損傷、胸徑徑階為4 cm。具體實驗過程見圖1。
1.3 數(shù)據(jù)處理與分析
實驗數(shù)據(jù)在Excel 2003統(tǒng)計軟件中進行整理和圖表制作,重復測量方差分析采用SPSS 19.0統(tǒng)計軟件中的one-way repeated measures ANOVA方法,多重比較采用Duncan方法完成。所有測定值均為平均值±標準偏誤差。
2 結果分析
2.1 雷竹克隆系統(tǒng)中各器官有機碳含量變化規(guī)律
出筍前各器官含碳率分配格局:枝(52.64%)>葉(47.18%)>稈(40.98%)>鞭(40.13%)>根(35.14%)。出筍完成后各器官含碳率分配格局:枝(48.20%)>稈(47.84%)>葉(45.53%)>鞭(45.52%)>根(44.29%)。出筍前后,各器官碳素分布格局發(fā)生變化,枝、葉含碳率降低,根、稈、鞭含碳率升高。
采用重復測量法分析出筍期不同分株數(shù)雷竹克隆系統(tǒng)各器官有機碳含量結果顯示,單株、雙株、三株雷竹克隆系統(tǒng)有機碳含量隨時間變化差異均顯著,各系統(tǒng)中雷竹分株不同器官間有機碳含量差異均顯著,處理時間及分株器官對各系統(tǒng)有機碳含量均有交互作用,即各系統(tǒng)有機碳含量隨著處理時間的變化而變化,并且時間因素因器官的不同而不同(表1)。
不同分株數(shù)雷竹克隆系統(tǒng)立竹器官有機碳含量的多重比較表明,單株雷竹克隆系統(tǒng)中雷竹各器官有機碳含量:枝>稈>鞭>葉>根;其中枝有機碳含量顯著高于葉、稈、鞭,枝、葉、稈、鞭有機碳含量顯著高于根;雙株雷竹克隆系統(tǒng)中雷竹各器官有機碳含量:枝>葉>稈>鞭>根,其中枝、葉、根、稈有機碳含量顯著高于根;三株雷竹克隆系統(tǒng)中雷竹各器官有機碳含量:枝>葉>稈>鞭>根;其中枝有機碳含量顯著高于葉、稈、鞭,枝、葉、稈、鞭有機碳含量顯著高于根(表2)。
表1 不同分株數(shù)雷竹克隆系統(tǒng)各器官有機碳含量重復測量方差分析結果
Table 1 ANOVA for repeated measurements in organic carbon content of the various organs in different rametsPhyllostachysviolascenssystem
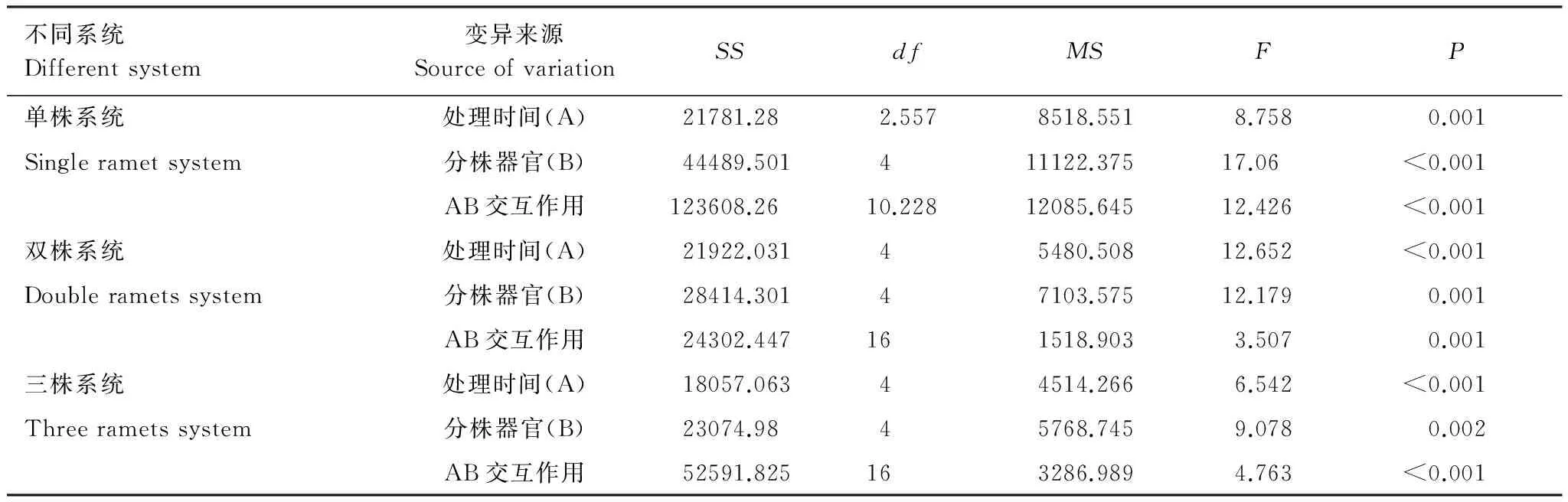
不同系統(tǒng)Differentsystem變異來源SourceofvariationSSdfMSFP單株系統(tǒng)處理時間(A)21781.282.5578518.5518.7580.001Singlerametsystem分株器官(B)44489.501411122.37517.06<0.001AB交互作用123608.2610.22812085.64512.426<0.001雙株系統(tǒng)處理時間(A)21922.03145480.50812.652<0.001Doublerametssystem分株器官(B)28414.30147103.57512.1790.001AB交互作用24302.447161518.9033.5070.001三株系統(tǒng)處理時間(A)18057.06344514.2666.542<0.001Threerametssystem分株器官(B)23074.9845768.7459.0780.002AB交互作用52591.825163286.9894.763<0.001
SS: 離差平方和;df: 自由度;MS: 均方
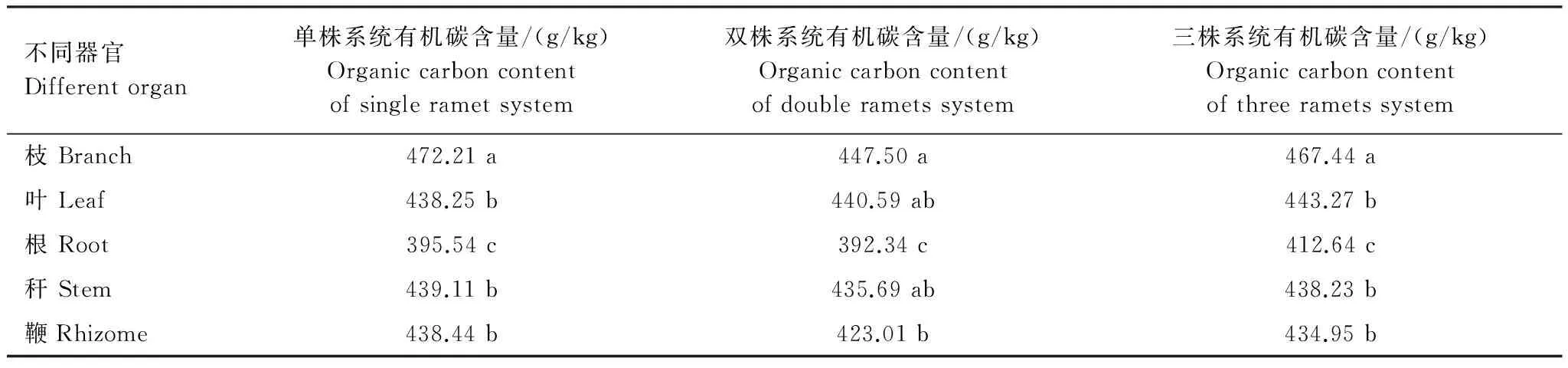
表2 不同分株數(shù)雷竹克隆系統(tǒng)中各器官出筍期有機碳含量差異性比較
*小寫字母表示同一系統(tǒng)不同器官比較; 不同小寫字母表示差異顯著(P<0.05),相同小寫字母表示差異不顯著(P>0.05)
由圖2可知,出筍期雷竹克隆系統(tǒng)中不同器官有機碳含量變化規(guī)律不同,隨處理時間的延長,枝、葉有機碳含量呈現(xiàn)先降低后升高的趨勢,根、稈、鞭有機碳含量呈“N”型變化趨勢;如圖1所示,單株雷竹克隆系統(tǒng)中,出筍前(1月)各器官有機碳含量差異顯著,各器官有機碳含量依次為:枝>葉>稈>鞭>根;隨出筍開始,雷竹枝、葉有機碳含量顯著降低,根、稈、鞭有機碳含量顯著升高,至3月份根、稈、鞭有機碳含量已顯著高于枝、葉;隨出筍期延長,雷竹各器官有機碳含量差異減小,至5月份筍期結束,除根外其余器官有機碳含量無顯著差異,各器官有機碳含量依次為:枝>稈>鞭>葉>根。

圖2 雷竹克隆系統(tǒng)各器官出筍期有機碳含量Fig.2 The organic carbon content of different organs in different phyllostachys violascens system字母表示同一時期同一系統(tǒng)內不同器官比較;不同小寫字母表示差異顯著(P<0.05),相同小寫字母表示差異不顯著(P>0.05)
雙株雷竹克隆系統(tǒng)中(圖2),出筍前(1月)各器官有機碳含量差異顯著,各器官有機碳含量依次為:枝>葉>稈>鞭>根;隨出筍開始,雷竹枝、葉有機碳含量顯著降低,根、稈、鞭有機碳含量顯著升高,至3月份稈、鞭有機碳含量已顯著高于枝、葉、根;隨出筍期延長,雷竹枝、葉有機碳含量逐漸上升而根、稈、鞭有機碳含量逐漸降低,至4月份枝、葉有機碳含量顯著高于根、稈、鞭;隨出筍期結束,各器官有機碳含量均有所上升,至5月份筍期結束,各器官有機碳含量依次為:稈>枝>葉>鞭>根。
三株雷竹克隆系統(tǒng)中(圖2),出筍前(1月)枝、葉有機碳含量顯著高于根、稈、鞭,各器官有機碳含量依次為:枝>葉>鞭>稈>根;隨出筍開始,雷竹枝、葉有機碳含量顯著降低,根、稈、鞭有機碳含量顯著升高,至3月份根、稈、鞭有機碳含量已顯著高于枝、葉;隨出筍期延長,雷竹枝、葉有機碳含量逐漸上升而根、稈、鞭有機碳含量逐漸降低,至4月份枝、葉有機碳含量高于根、稈、鞭;隨出筍期結束,各器官有機碳含量均有所上升,至5月份筍期結束,各器官有機碳含量無顯著差異,有機碳含量依次為:枝>稈>根>葉>鞭。
上述結果表明,雷竹不同器官有機碳含量分布格局為出筍前:枝>葉>稈>鞭>根,出筍完成后:枝>稈>葉>鞭>根,枝、葉有機碳含量顯著降低,根、稈、鞭有機碳含量顯著升高,不同雷竹克隆系統(tǒng)中各器官有機碳含量差異均顯著。不同分株雷竹克隆系統(tǒng)筍期各器官有機碳含量發(fā)生顯著變化,枝、葉有機碳含量均呈先降低后升高趨勢,根、稈、鞭有機碳含量均呈“N”型變化規(guī)律,但變化時期不盡相同,說明出筍影響雷竹各器官的有機碳含量分布,出筍期各器官間源-匯關系格局發(fā)生變化,雷竹不同器官間碳素資源在源-匯關系驅動下發(fā)生轉移,使各器官有機碳分布格局發(fā)生變化。
2.2 不同分株系統(tǒng)雷竹克隆系統(tǒng)有機碳含量變化規(guī)律分析
采用重復測量法分析不同分株數(shù)雷竹克隆系統(tǒng)出筍期各器官有機碳含量結果表明(表3):雷竹克隆系統(tǒng)中枝、葉、根、稈、鞭各器官有機碳含量隨時間變化差異均顯著(P<0.05),不同系統(tǒng)間,鞭有機碳含量差異顯著,其他器官均無顯著差異。不同分株數(shù)與監(jiān)測時間對枝、根有機碳含量有交互作用,對葉、稈、鞭無交互作用,即各器官有機碳含量隨筍期延長而不同,并且時間因素因分株數(shù)量不同而不同。
表3 不同分株數(shù)雷竹克隆系統(tǒng)各器官有機碳含量重復測量方差分析結果
Table 3 ANOVA for repeated measurements in organic carbon content of the various organs in different rametsPhyllostachysviolascenssystem
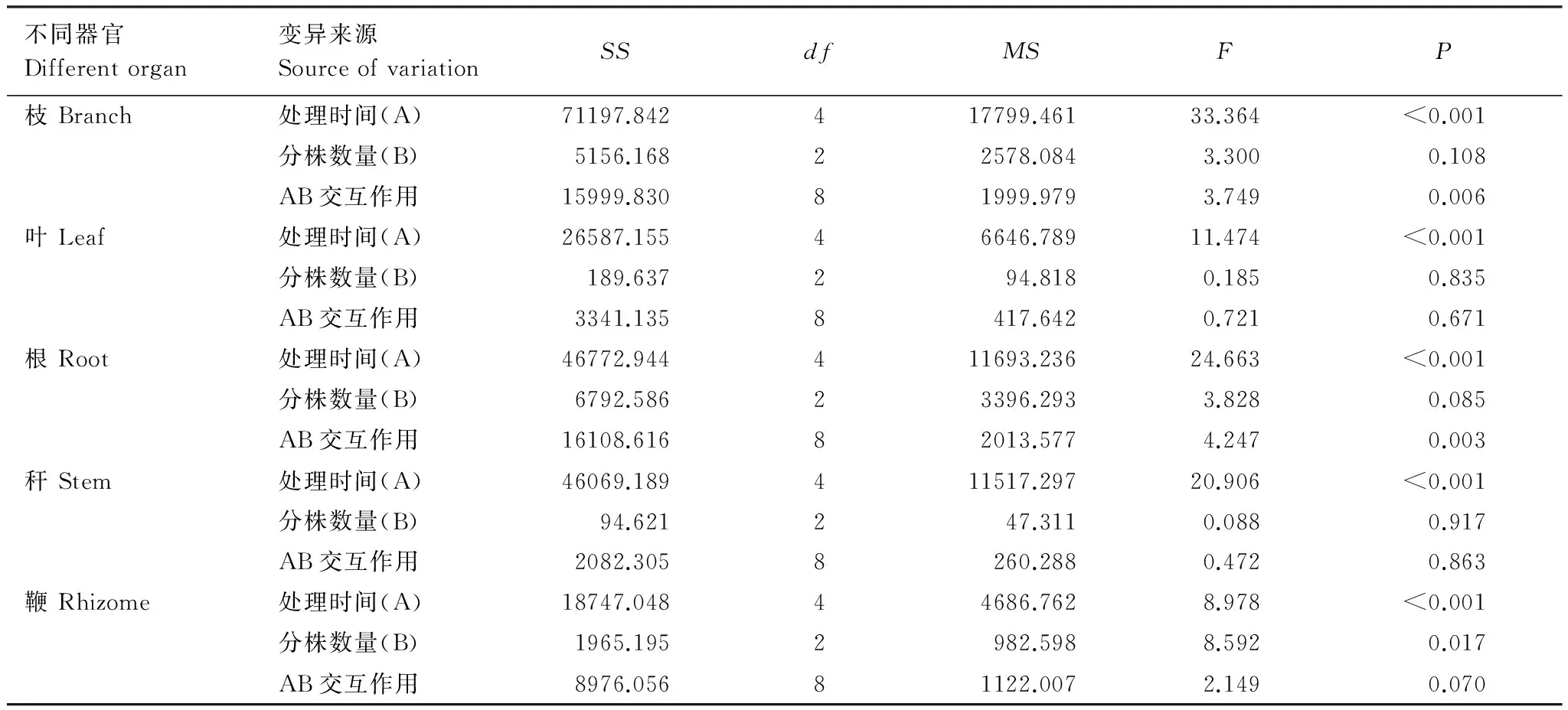
不同器官Differentorgan變異來源SourceofvariationSSdfMSFP枝Branch處理時間(A)71197.842417799.46133.364<0.001分株數(shù)量(B)5156.16822578.0843.3000.108AB交互作用15999.83081999.9793.7490.006葉Leaf處理時間(A)26587.15546646.78911.474<0.001分株數(shù)量(B)189.637294.8180.1850.835AB交互作用3341.1358417.6420.7210.671根Root處理時間(A)46772.944411693.23624.663<0.001分株數(shù)量(B)6792.58623396.2933.8280.085AB交互作用16108.61682013.5774.2470.003稈Stem處理時間(A)46069.189411517.29720.906<0.001分株數(shù)量(B)94.621247.3110.0880.917AB交互作用2082.3058260.2880.4720.863鞭Rhizome處理時間(A)18747.04844686.7628.978<0.001分株數(shù)量(B)1965.1952982.5988.5920.017AB交互作用8976.05681122.0072.1490.070
不同分株系統(tǒng)中各器官有機碳含量隨出筍期延長發(fā)生變化,由表4可知,單株、雙株、三株雷竹克隆系統(tǒng)枝有機碳含量1月份單株系統(tǒng)顯著高于雙株、三株系統(tǒng),其他時期無顯著差異;葉、稈有機碳含量筍期無顯著差異;根有機碳含量1月份差異顯著,三株系統(tǒng)高于單、雙株系統(tǒng),2月份無顯著差異,3月份雙株系統(tǒng)顯著高于單、三株系統(tǒng),4月份無顯著差異,至5月份三株系統(tǒng)顯著高于單、雙株系統(tǒng);鞭有機碳含量出筍前各系統(tǒng)無顯著差異,2月份單株系統(tǒng)顯著高于雙株、三株系統(tǒng),隨筍期延長至出筍完成各系統(tǒng)無顯著差異。
由表4可得,單株系統(tǒng)各器官含碳率降幅:枝(17.99%)>稈>鞭>葉>根(5.61%);雙株系統(tǒng)各器官含碳率降幅:枝(6.65%)>葉>稈>根>鞭(3.51%);三株系統(tǒng)各器官含碳率降幅:枝(10.99%)>葉>稈>根>鞭(1.44%)。總體來講,不同分株數(shù)雷竹克隆系統(tǒng)各器官有機碳含量降幅:單株系統(tǒng)>多株系統(tǒng)。不同分株數(shù)雷竹克隆系統(tǒng)出筍的含碳量不同,單株系統(tǒng)(24.62g)<雙株系統(tǒng)(29.30g)<三株系統(tǒng)(44.55g),由此可知,單株雷竹克隆系統(tǒng)與多株克隆系統(tǒng)間各器官有機碳含量降幅與出筍量成反比。
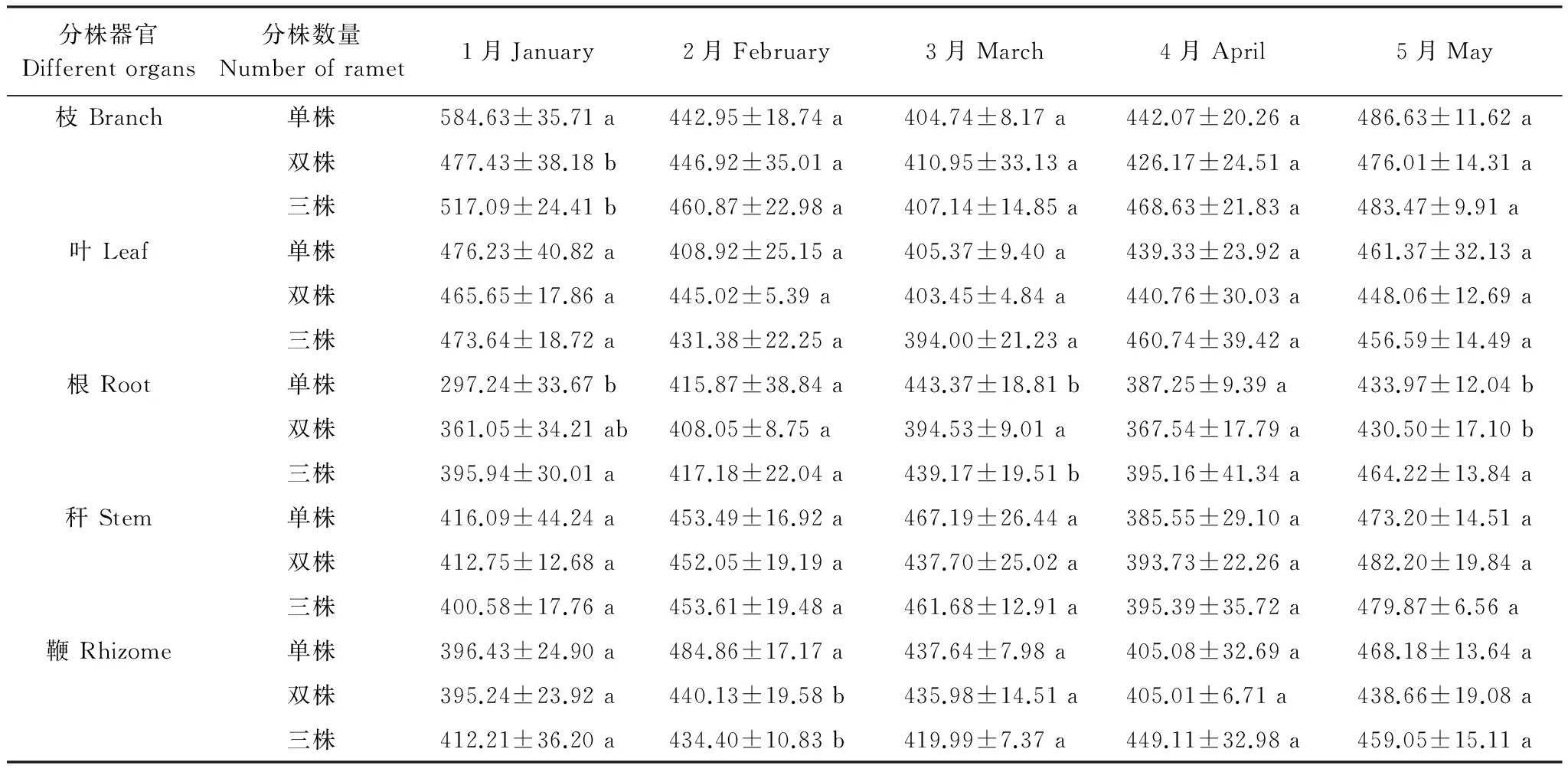
表4 不同分株數(shù)雷竹克隆系統(tǒng)各器官出筍期有機碳含量/(g/kg)
上述結果表明,筍期葉、稈有機碳含量不同分株數(shù)雷竹克隆系統(tǒng)間無顯著差異,枝、根、鞭有機碳含量不同分株數(shù)雷竹克隆系統(tǒng)在出筍不同時期存在顯著差異。單株雷竹克隆系統(tǒng)和多株雷竹克隆系統(tǒng)間各器官有機碳含量降幅與出筍量成反比,即隨克隆分株數(shù)量增加,出筍量增加,而系統(tǒng)內分株各器官有機碳含量降幅減小,不同雷竹克隆系統(tǒng)分株數(shù)量影響雷竹各器官有機碳含量變化。
2.3 多株雷竹克隆系統(tǒng)不同年齡分株有機碳分布變化規(guī)律
采用單因素重復測量法分析雙株、三株雷竹克隆系統(tǒng)中不同年齡分株出筍期各器官有機碳含量,結果如表5所示,雙株、三株系統(tǒng)各器官出筍不同階段有機碳含量差異均顯著。雙株系統(tǒng)中不同年齡雷竹鞭出筍期有機碳含量差異顯著,其它器官則差異不顯著;監(jiān)測時間與分株年齡對根、稈有機碳含量有交互作用,即根、稈的有機碳含量隨時間變化而變化,且時間因素隨雷竹克隆分株系統(tǒng)的不同而不同,對其他器官有機碳含量則無交互作用。三株系統(tǒng)中不同年齡雷竹枝、稈、鞭出筍期有機碳含量差異顯著,葉、根出筍期有機碳含量差異則不顯著;監(jiān)測時間與分株年齡對葉、根、稈出筍期含碳量變化有交互作用,對其他器官則無交互作用。
表5 不同年齡雷竹克隆分株各器官有機碳含量重復測量方差分析結果
Table 5 ANOVA for repeated measurements in organic carbon content of the various organs in different rametsPhyllostachysviolascenssystem

不同器官Differentorgans不同系統(tǒng)Differentsystem變異來源SourceofvariationSSMSdfFP枝Branch雙株處理時間(A)208995224.75547.477<0.001分株年齡(B)3.1153.11510.0210.892AB交互作用808.046202.01240.2890.881三株處理時間(A)61731.03915432.76422.485<0.001分株年齡(B)3161.1571580.57825.2530.048AB交互作用6162.281770.28581.1220.384葉Leaf雙株處理時間(A)10303.72575.91542.6820.069分株年齡(B)131.243131.24310.1460.722AB交互作用3798.81949.70340.9890.442三株處理時間(A)40407.3910101.85415.571<0.001分株年齡(B)3654.9631827.48122.7380.143AB交互作用30676.6793834.58585.911<0.001根Root雙株處理時間(A)20286.15071.515414.568<0.001分株年齡(B)2067.022067.02111.2740.322AB交互作用6353.361588.33944.5630.012三株處理時間(A)41860.94310465.24426.961<0.001分株年齡(B)5837.3372918.66822.710.145AB交互作用20520.4912565.06186.608<0.001稈Stem雙株處理時間(A)35126.38781.569410.283<0.001分株年齡(B)1634.571634.57211.2650.324AB交互作用12183.73045.92443.5670.029三株處理時間(A)73679.50318419.88433.788<0.001分株年齡(B)7935.1593967.579210.5680.011AB交互作用29700.2913712.53686.81<0.001鞭Rhizome雙株處理時間(A)27432.22420263.121.35411.7130.014分株年齡(B)4569.194569.19116.1540.016AB交互作用674.517498.2391.3540.2880.68三株處理時間(A)49727.14223699.462.09825.157<0.001分株年齡(B)18691.5439345.771212.9020.007AB交互作用1260.965300.4824.1960.3190.867
如表6所示,分析雙株、三株雷竹克隆系統(tǒng)中不同年齡雷竹分株各器官有機碳含量可知,出筍前(1月),雙株系統(tǒng)中不同年齡分株僅鞭有機碳含量差異顯著,而三株系統(tǒng)中葉、根、鞭不同年齡分株差異顯著,不同器官差異性不同。隨筍期延長,雙株、三株系統(tǒng)中不同年齡分株各器官有機碳含量差異性發(fā)生變化。由表6可知,出筍期,各器官有機碳含量均有先降低再升高趨勢,雙株系統(tǒng)中1年生雷竹各器官含碳率降幅:枝(8.71%)>稈(7.48%)>根(4.75%)>鞭(4.38%)>葉(3.59%);2年生雷竹各器官含碳率降幅:枝(6.78%)>鞭(4.49%)>根(3.93%)>稈(3.55%)>葉(2.57%)。總體來講,1年生雷竹出筍期有機碳含量降幅大于2年生竹。三株系統(tǒng)中,1年生雷竹各器官含碳率降幅:稈(12.46%)>枝(9.26%)>根(7.07%)>葉(4.16%)>鞭(3.37%);2年生雷竹各器官含碳率降幅:枝(11.52%)>葉(9.23%)>根(4.59%)>稈(3.53%)>鞭(3.11%);3年生雷竹各器官含碳率降幅:葉(15.05%)>枝(12.2%)>稈(7.83%)>根(7.38%)>鞭(4.83%)。總體來講,同一器官不同年齡出筍時有機碳含量降幅為3年生雷竹降幅大于1年、2年生竹,表明同一系統(tǒng)中分株年齡影響出筍時各器官有機碳含量變化量。
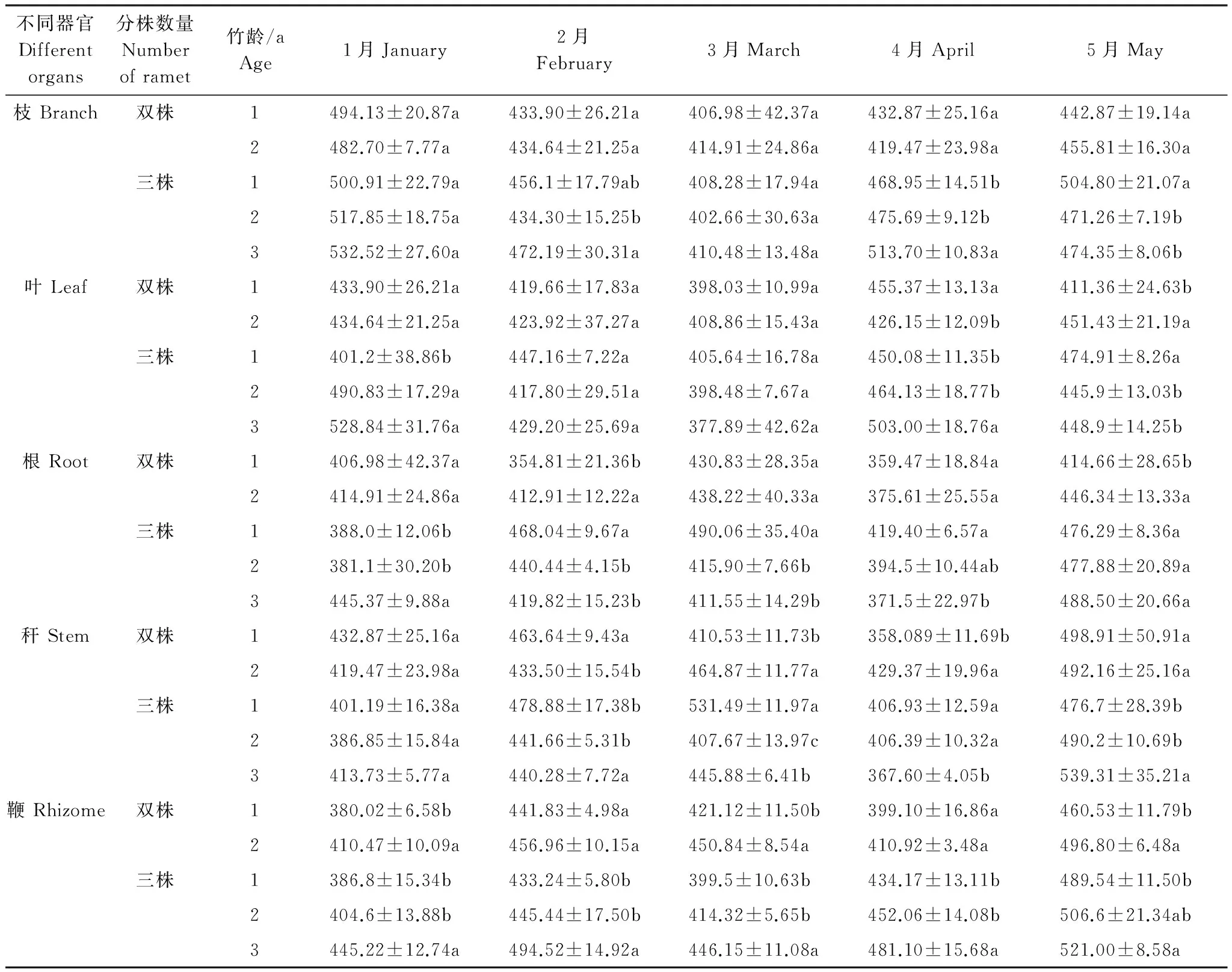
表6 不同分株數(shù)雷竹克隆系統(tǒng)不同年齡雷竹各器官有機碳含量/(g/kg)
上述結果表明,多株雷竹克隆系統(tǒng)中不同年齡分株有機碳含量變化量不同,雙株系統(tǒng)中1年生雷竹各器官有機碳含量降幅大于2年生竹,三株系統(tǒng)中3年生雷竹各器官有機碳含量降幅大于1年、2年生竹,說明分株年齡是影響雷竹不同器官對出筍響應有機碳含量變化的因素之一。
3 結論與討論
克隆植物具有生理整合、克隆生長等獨特生態(tài)適應特性[34- 36],在異質環(huán)境條件下,這種特性會體現(xiàn)得更加明顯[37]。碳素或光合產(chǎn)物在克隆植物分株間的分配是其生理整合過程中非常重要的一部分,由于特殊的生長、繁殖方式,使得克隆植物的碳素分配格局更加復雜[38]。克隆植物種類不同、所處的生境及發(fā)育階段不同,其碳素的分配格局也可能不同[1]。本研究發(fā)現(xiàn),雷竹不同器官有機碳含量范圍為:35.14%—52.64%,大小順序依次為:枝(52.64%)>葉(47.18%)>稈(40.98%)>鞭(40.13%)>根(35.14%),這與王兵等[39]研究的毛竹各器官含碳率介于42.22%—47.53%,各器官大小順序依次為:稈(47.53%)>枝(46.49%)>鞭(46.10%)>根(45.30%)>葉(42.22%)不同,由此可以看出,各器官碳含量的分配格局因竹種的不同而存在差異。
同種克隆植物的碳素整合格局會因環(huán)境資源條件及植株生長發(fā)育階段等因素的不同發(fā)生適應性調整或改變[40],研究表明隨光照強度增加,華西箭竹(Fargesianitida)的生物量積累逐漸增大[41],本研究表明出筍生長影響雷竹克隆系統(tǒng)中分株各器官碳素分配格局,各器官間有機質資源發(fā)生轉移,出筍生長使雷竹無性系種群產(chǎn)生生理整合的響應。理論上,竹林出筍時會造成種群資源損耗,種群通過提高生產(chǎn)強度以應對這種損耗,3—4月為出筍期,由于此時環(huán)境溫度低,光照較弱,光合作用固定同化物能力較弱,而出筍生長十分迅速,僅依靠光合作用固定同化物不能滿足筍的快速生長,主要還是依靠竹林種群的營養(yǎng)儲存,這與潘雁紅等人的研究結果一致[42]。尚未發(fā)育完全的幼株通常作為接受同化產(chǎn)物的匯,而相連的母株則是制造和輸出同化產(chǎn)物的源[1],即在出筍過程中,母株作為同化物源利用自身貯存的有機同化物資源供應筍生長,這與對根莖克隆植物畢氏苔草(Carexbigelowii)和燈心草(Unciniameridensis)的研究結果相同[10]。在出筍生長的不同階段,雷竹分株枝、葉含碳率隨時間呈“∨”型變化趨勢,根、稈、鞭含碳率呈“N”型變化趨勢,各器官間的源-匯格局發(fā)生變化,推斷這可能是由于雷竹克隆系統(tǒng)內母株充當制造和輸出同化產(chǎn)物的源,將同化產(chǎn)物通過連接物(地下莖)源源不斷的供給幼筍,在同一分株內枝、葉、稈中貯存的同化物傳遞至根、鞭貯存,出筍生長開始時根、鞭由原來有機同化物的匯變?yōu)樵蠢米陨碣A存的有機同化物供應筍的快速生長。隨著幼株發(fā)育過程逐漸完成,它們可能最終變得完全獨立而不再需要鄰近相連母株的支持,于是便停止這種運輸[43],則母株各器官含碳率逐漸上升,這與Price[44]對克隆植物虎仗(Reynoutriajaponica)的研究結果基本一致,即在冬季根、稈中同化物含量增加,成為源-匯關系中的匯,次年春季分株進行生長時,根、稈成為同化物的源將貯存的有機物運輸至新生分株中。不同系統(tǒng)各器官有機碳含量下降或上升時期有所不同,這是由于在不同系統(tǒng)中,開始發(fā)筍及筍停止生長的時間有所差異。
本研究中,由于所選雷竹樣株生長狀況基本相同,各株徑階大小一致,可視為各株雷竹生物量大小基本相同。分析發(fā)現(xiàn)單株、多株系統(tǒng)各器官含碳率降幅與出筍量成反比,即單株系統(tǒng)出筍量小于多株系統(tǒng),但各器官出筍時含碳率降幅大于多株系統(tǒng),可能是由于多株系統(tǒng)水平方向分株間也存在源匯關系[44],相連分株有機碳資源均向相連基部運輸供應筍的生長,多個分株同時為出筍提供有機質資源,因此多株系統(tǒng)各器官含碳率降幅相對于單株低。由于分株比例越大,供體分株生物量越大,在碳素資源傳遞過程中,供體分株的耗損對供體分株造成的損失越小,同時由于分株比例越大,資源傳輸需要消耗的能量也越少,對供體分株的耗損越小,因而生理整合強度越大[45- 46]。生理整合研究表明:資源在克隆分株間的傳遞具有雙向性[47],王昱生等人利用14C標記的方法示蹤克隆羊草內部光合有機產(chǎn)物存在向頂、向基及水平方向的傳遞,但其傳遞方向及傳遞的量很難精確描述[2]。本研究對不同分株數(shù)的克隆系統(tǒng)研究也表明在面對一定的資源壓力時,多株系統(tǒng)分株間資源進行共享,其共享強度還需要在以后的研究中進行進一步探討。出筍期,多株雷竹克隆系統(tǒng)內分株年齡影響各器官含碳率的變化幅度,雙株系統(tǒng)1年生竹變化量大于2年生竹,3年生竹變化量大于1、2年生竹,可能是由于1年生竹光合能力較強,3年生竹貯存的同化物較多,通過相連的基莖為出筍提供碳素資源,這與石松屬植物(Lycopdiumantoninum)及沙丘苔草(Carexarenaria)植物的研究結果一致[1],即不同年齡分株整合強度及整合能力可能不同,這是因為不同年齡分株,可以具有不同的光合能力和光合生產(chǎn)量[2],各分株之間資源轉移速率與轉移強度很可能是不同的。
綜上所述,雷竹不同器官對出筍的響應為有機質資源發(fā)生轉移,分株內碳素分配格局發(fā)生變化,各器官間的源-匯關系也會發(fā)生改變以滿足筍的快速生長。供體分株數(shù)量與出筍量是影響碳素資源傳遞的重要因素,但決定相連分株間碳素資源轉移強度的并不完全是分株數(shù)目和出筍量,還包括克隆系統(tǒng)內雷竹分株的年齡。在經(jīng)營雷竹林時,提高雷竹林株間連接度,在除草、施肥、整地時避免斷鞭,以便提高碳素資源整合度,調整雷竹林的年齡結構,相對較多地保留高光合能力的1年生竹和同化物貯存能力較高的3年生竹,調整雷竹林密度,提高雷竹林光合生產(chǎn)力對提高雷竹林的出筍量及其經(jīng)濟效益有一定的現(xiàn)實意義。對于克隆植物內部資源的整合影響因素較多,內部生理機制的影響及同化物轉移強度、轉移方向在未來研究中可以通過同位素示蹤及多年限重復試驗進行進一步研究,對雷竹林的經(jīng)營具有十分重要的意義。
[1] 盛麗娟, 李德志, 朱志玲, 王緒平, 石強, 王超華, 范旭麗, 柯世朕. 克隆植物的碳素生理整合及其生態(tài)學效應. 應用與環(huán)境生物學報, 2007, 13(6): 888- 894.
[2] 王昱生, 洪銳民, 黃大明, 滕小華, 李月樹, 鹽見正衛(wèi), 中村未樹. 羊草種群克隆分株之間光合產(chǎn)物的轉移. 生態(tài)學報, 2004, 24(5): 900- 907.
[3] 于文龍, 許民. 區(qū)域尺度下不同樹種木材含碳率測定研究. 安徽農業(yè)科學, 2013, 41(4): 1571- 1572.
[4] 徐小靜, 朱向輝, 汪方德, 毛華英, 程清明, 汪良義, 汪傳佳, 朱湯軍. 毛竹等11種浙江省碳匯造林樹種含碳率分析. 竹子研究匯刊, 2009, 28(1): 21- 24.
[5] 周國模, 姜培坤. 毛竹林的碳密度和碳貯量及其空間分布. 林業(yè)科學, 2004, 40(6): 20- 24.
[6] Zhang Q Z, Wang C K, Wang X C, Quan X K. Carbon concentration variability of 10 Chinese temperate tree species. Forest Ecology and Management, 2009, 258(5): 722- 727.
[7] 李江, 翟明普, 朱宏濤, 孟夢, 邱瓊, 毛云玲, 陳宏偉, 郭永清. 思茅松人工中幼林的含碳率研究. 福建林業(yè)科技, 2009, 36(4): 12- 15.
[8] 付堯, 孫玉軍. 植物有機碳測定研究進展. 世界林業(yè)研究, 2013, 26(1): 24- 30.
[9] Mogie M, Hutchings M J. Phylogeny, ontogeny and clonal growth in vascular plant. In: van Groenendael J, de Kroon H, eds. Clonal Growth in Plants: Regulation and Function. The Hague: SPB Academic Publishing, 1990: 3- 22.
[10] 何瑩瑩, 李德志, 徐潔, 王浩, 程立麗, 劉微. 不同光照條件下結縷草全株碳素生理整合格局及其生態(tài)效應. 植物資源與環(huán)境學報, 2013, 22(4): 1- 10.
[11] Li D Z, Takahashi S. Particularities of clonal plant species induced by physiological integration. Journal of Japanese Society of Grassland Science, 2003, 49(4): 395- 402.
[12] Hartnett D C, Bazzaz F A. Physiological integration among intraclonal ramets inSolidagocanadensis. Ecology, 1983, 64(4): 779- 788.
[13] Hartnett D C, Bazzaz F A. The genet and ramet population dynamics ofSolidagocanadensisin an abandoned field. Journal of Ecology, 1985, 73(2): 407- 413.
[14] Hartnett D C, Bazzaz F A. The integration of neighbourhood effects by clonal genets inSolidagocanadensis. Journal of Ecology, 1985, 73(2): 415- 427.
[15] 褚延梅, 楊健, 李景吉, 彭培好. 三種增溫情景對入侵植物空心蓮子草形態(tài)可塑性的影響. 生態(tài)學報, 2014, 34(6): 1411- 1417.
[16] 朱志玲. 不同土壤水分條件下斷稈處理對克隆植物結縷草表型可塑性的影響. 安徽農業(yè)科學, 2010, 38(33): 18627- 18630.
[17] 湯俊兵, 肖燕, 安樹青. 根稈克隆植物生態(tài)學研究進展. 生態(tài)學報, 2010, 30(11): 3028- 3036.
[18] 付京晶, 周華坤, 趙新全, 陳哲, 溫軍, 姚步青, 楊元武, 金艷霞. 草原克隆植物對光、溫、水、肥和牧壓變化梯度的響應. 草業(yè)科學, 2013, 30(4): 616- 626.
[19] 劉駿, 楊清培, 楊光耀, 方楷, 章情, 戴艷艷, 樂冬英, 歐陽明. 厚壁毛竹非結構性碳水化合物分配格局. 江西農業(yè)大學學報, 2011, 33(5): 924- 928.
[20] 何亞平, 費世民, 蔣俊明, 陳秀明, 余英, 唐森強, 朱維雙. 長寧毛竹和苦竹有機碳空間分布格局. 四川林業(yè)科技, 2007, 28(5): 10- 14.
[21] 張稱意, 楊持, 董鳴. 根稈半灌木羊柴對光合同化物的克隆整合. 生態(tài)學報, 2001, 21(12): 1986- 1993.
[22] Li D Z, Takahashi S, Zhu T C. Effects of soil nitrogen availability and clonal integration on the branching behaviors ofZoysiajaponica. Acta Prataculturae Sinica, 2006, 15(5): 135- 146.
[23] Price E A C, Marshall C, Hutching M J. Studies of growth in the clonal herbGlechomahederacea. I. Patterns of physiological integration. Journal of Ecology, 1992, 80(1): 25- 38.
[24] 陳勁松, 董鳴, 于丹, 劉慶. 不同光照條件下聚花過路黃的克隆構型和分株種群特征. 應用生態(tài)學報, 2004, 15(8): 1383- 1388.
[25] Ren F, Zhou H K, Zhao X Q, Han F, Shi L N, Duan J C, Zhao J Z. Influence of simulated warming using OTC on physiological-biochemical characteristics ofElymusnutansin alpine meadow on Qinghai-Tibetan plateau. Acta Ecologica Sinica, 2010, 30(3): 166- 171.
[26] 徐德兵, 趙粉俠, 賀斌, 李甜江, 段宗亮, 李根前. 中國沙棘克隆生長格局對不同灌水強度的響應. 東北林業(yè)大學學報, 2008, 36(9): 31- 32, 41- 41.
[27] 馬欽彥, 陳遐林, 王娟, 藺琛, 康峰峰, 曹文強, 馬志波, 李文宇. 華北主要森林類型建群種的含碳率分析. 北京林業(yè)大學學報, 2002, 24(5/6): 96- 100.
[28] Yen T M, Ji Y J, Lee J S. Estimating biomass production and carbon storage for a fast-growing makino bamboo (Phyllostachysmakinoi) plant based on the diameter distribution model. Forest Ecology and Management, 2010, 260(3): 339- 344.
[29] Sohel S I, Alamgir M, Akhter S, Rahman M. Carbon storage in a bamboo (Bambusavulgaris) plantation in the degraded tropical forests: implications for policy development. Land Use Policy, 2015, 49: 142- 151.
[30] Yen T M, Lee J S. Comparing aboveground carbon sequestration between moso bamboo (Phyllostachysheterocycla) and China fir (Cunninghamialanceolata) forests based on the allometric model. Forest Ecology and Management, 2011, 261(6): 995- 1002.
[31] Nath A J, Das G, Das A K. Above ground standing biomass and carbon storage in village bamboos in North East India. Biomass and Bioenergy, 2009, 33(9): 1188- 1196.
[32] 方楷, 楊清培, 趙廣東, 楊光耀, 施建敏, 郭起榮. 江西主要竹類植物碳密度的時空差異分析. 江西科學, 2010, 28(4): 449- 452.
[33] Wang Z W, Li L H, Han X G, Dong M. Do rhizome severing and shoot defoliation affect clonal growth ofLeymuschinensisat ramet population level?. Acta Oecologica, 2004, 26(3): 255- 260.
[34] 周資行, 焦健, 李毅, 李永兵, 張世虎. 民勤沙拐棗克隆種群構件結構及與環(huán)境因子的灰色關聯(lián)分析. 林業(yè)科學, 2012, 48(5): 141- 149.
[35] Riis T, Lambertini C, Olesen B, Clayton J S, Brix H, Sorrell B K. Invasion strategies in clonal aquatic plants: Are phenotypic differences caused by phenotypic plasticity or local adaptation?. Annals of Botany, 2010, 106(5): 813- 822.
[36] Yu F H, Wang N, He W M, Chu Y, Dong M. Adaptation of rhizome connections in drylands: increasing tolerance of clones to wind erosion. Annals of Botany, 2008, 102(4): 571- 577.
[37] Liu H D, Yu F H, He W M, Chu Y, Dong M. Are clonal plants more tolerant to grazing than co-occurring non-clonal plants in inland dunes?. Ecological Research, 2007, 22(3): 502- 506.
[38] Marshall C, Sgar G R. The interdependence of tillers inLoliummultiflorumLam.—a quantitative assessment. Journal of Experimental Botany, 1968, 19(4): 785- 794.
[39] 王兵, 楊清培, 郭起榮, 趙廣東, 方楷. 大崗山毛竹林與常綠闊葉林碳儲量及分配格局. 廣西植物, 2011, 31(3): 342- 348.
[40] Xu C Y, Schooler S S, Van Klinken R D. Effects of clonal integration and light availability on the growth and physiology of two invasive herbs. Journal of Ecology, 2010, 98(4): 833- 844.
[41] 陶建平,宋利霞. 亞高山暗針葉林不同林冠環(huán)境下華西箭竹的克隆可塑性. 生態(tài)學報,2006,26(12):4019- 4026.
[42] 潘雁紅, 高貴賓, 吳良如, 鐘浩, 周鴻翔, 袁娜. 生理整合對覆蓋雷竹林克隆分株光合性狀的影響. 安徽農業(yè)科學, 2015, 43(17): 230- 231, 245- 245.
[43] Marshall C. Source-sink relations of interconnected ramets//van Groenendael J M, de Kroon H, eds. Clonal Growth in Plants: Regulation and Function. The Hague: SPB Academic Publishing, 1990: 23- 41.
[44] Price E A C, Gamble R, Williams G G, Marshall C. Seasonal patterns of partitioning and remobilization of14C in the invasive rhizomatous perennial Japanese knotweed (Fallopiajaponica(Houtt.) Ronse Decraene). Evolutionary Ecology, 2001, 15(4/6): 347- 362.
[45] Li Q, Liu X, Yue M, Tang W T, Meng Q C. Response of physiological integration inTrifoliumrepensto heterogeneity of UV-B radiation. Flora-Morphology, Distribution, Functional Ecology of Plants, 2011, 206(8): 712- 719.
[46] 李倩, 劉曉, 張曉飛, 張瑞昌, 柴永福, 岳明. UV-B輻射方向對白三葉克隆整合的影響. 生態(tài)學報, 2014, 34(13): 3568- 3575.
[47] 程立麗, 李德志, 朱志玲, 何瑩瑩, 王浩, 紀倩倩, 劉微, 陳慧娟, 賴蘇雯. 克隆植物結縷草的水分生理整合格局特征及其生態(tài)效應分析. 西北植物學報, 2013, 33(7): 1442- 1451.
Organic carbon allocation pattern and changes regulation in various organs ofPhyllostachysviolascensclone system in shooting period
JI Linke, XIE Jinzhong*, ZHANG Wei, LU Peng, ZHANG Lei
TheResearchInstituteofSubtropicalForestry,TheChineseAcademyofForestry,Hangzhou311400,China
Carbon assimilation from photosynthesis provides most substrates and energy for plant growth. To date, the distribution of carbon in various plant species has been intensively studied. However, information on carbon distribution during the shooting period in bamboo plants is limited. In this study, carbon distribution in different organs ofPhyllostachysviolascensplants with one, two, or three ramets was investigated during the shooting period, which enabled further study of the mechanisms that regulate bamboo species development. The results showed that the organic carbon content was significantly different in various organs ofP.violascensand that it changed significantly during the shooting period. Before the shooting period started, organic carbon content decreased in the order branch > leaf > stem > ramet > root; after shooting was completed, it decreased in the order branch > stem > leaf > ramet > root. Organic carbon initially decreased and then subsequently increased in branch and leaf and there was an “N”-like variation in organic carbon content in root, stem, and rhizome. The decrease in organic carbon content was negatively related to the shoot yield in both single and multi-ramet plants. The magnitude of the reduction in organic carbon content of various organs decreased with increasing ramet number and shoot output. Variation in organic carbon content was also found between plants with different ramet age. In the two-ramet plants, the reduction in organic carbon content was larger in 1-year-old plants than in 2-year-old plants, and in three-ramet plants, the reduction in organic carbon content was larger in 3-year-old plants than in 2-year-old and 1-year-old plants. These findings indicate that bamboo shooting affects organic carbon allocation, with a decrease in branch and leaf content and an increase in root, stem, and rhizome content. The potential source-sink relationship of different organs changes to provide organic carbon for bamboo shoots. Physiological integration of photosynthetic products occurs within the clonal segments ofP.violascens. With an increasing number of ramets, organic carbon in the shoots increased and loss of organic carbon in the ramets decreased. The organic carbon content also varied withP.violascensplant age during the shooting period. These findings have important practical implication in terms of shoot output and economic benefit, by adjusting the natural above-ground structure ofP.violascens.
Phyllostachysviolascens; clone system;shooting period;organic carbon allocation pattern
浙江省林業(yè)科技推廣資助項目(2015B05);中國林業(yè)科學研究院基本科研業(yè)務費專項資金項目(2016)
2015- 12- 14;
2016- 05- 30
10.5846/stxb201512142488
*通訊作者Corresponding author.E-mail: jzhxie@163.net
冀琳珂,謝錦忠,張瑋,盧鵬,張磊.雷竹克隆系統(tǒng)出筍期有機碳分布變化規(guī)律.生態(tài)學報,2016,36(23):7624- 7634.
Ji L K, Xie J Z, Zhang W, Lu P, Zhang L.Organic carbon allocation pattern and changes regulation in various organs ofPhyllostachysviolascensclone system in shooting period.Acta Ecologica Sinica,2016,36(23):7624- 7634.
猜你喜歡
工業(yè)設計(2022年8期)2022-09-09 07:43:20
音樂探索(2022年2期)2022-05-30 21:01:37
軍民兩用技術與產(chǎn)品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
家庭影院技術(2017年9期)2017-09-26 03:41:45
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
