毒死蜱對斑馬魚胚胎氧化應激效應研究
2016-04-20 01:30:10張家禹劉麗麗李國超余凱敏閆艷春中國農業科學院研究生院北京100081
中國環境科學 2016年3期
關鍵詞:氧化應激
張家禹,劉麗麗,李國超,余凱敏,呂 鵬,閆艷春(中國農業科學院研究生院,北京 100081)
?
毒死蜱對斑馬魚胚胎氧化應激效應研究
張家禹,劉麗麗,李國超,余凱敏,呂鵬,閆艷春*(中國農業科學院研究生院,北京 100081)
摘要:以斑馬魚為模型,研究了毒死蜱對斑馬魚胚胎形態學的影響,及其對胚胎的氧化應激和氧化損傷作用.將斑馬魚胚胎暴露在梯度濃度的毒死蜱溶液中96h后,發現毒死蜱會造成斑馬魚胚胎嚴重畸形甚至死亡,其96h半致死濃度為1.18mg/L.對氧化應激相關基因的表達及抗氧化酶活性和丙二醛(MDA)含量進行檢測,結果表明,在毒死蜱脅迫下抗氧化酶(SOD、CAT)活性降低,并且其編碼基因(Cu/Zn-sod、Mn-sod、cat)的表達受到抑制;但在低濃度毒死蜱脅迫下,抗氧化酶活性并沒有受到顯著影響,而抗氧化酶基因的表達對毒死蜱更加敏感.毒死蜱能引起gstp2的表達上調,但GST活性與gstp2的表達變化并不一致.處理組胚胎中nrf2表達上調,從而上調抗氧化蛋白和II相解毒酶基因的表達.毒死蜱脅迫下,基因ucp2、cox1表達下調,能夠減少呼吸鏈ROS的產生.同時基因bcl2表達下調,表明凋亡的平衡受到破壞.毒死蜱處理組中MDA含量顯著升高,說明毒死蜱能造成斑馬魚胚胎氧化損傷.
關鍵詞:毒死蜱;斑馬魚胚胎;氧化應激;氧化損傷
* 責任作者, 教授, yanyanchun@caas.cn
毒死蜱是一種高效、廣譜、中等毒性的有機磷類殺蟲殺螨劑,對害蟲具有觸殺、胃毒和熏蒸作用.自從國家對高毒農藥的禁限用政策出臺后,毒死蜱作為主要替代品種之一,廣泛應用于農業和城市害蟲的防治[1-2].毒死蜱的大量使用對生態環境已產生了負面影響,研究顯示可在多種水體中檢測到其殘留[3-4],在農作物種植區附近的水域中其殘留量往往更高,水稻田出水中毒死蜱殘留最高達26.07μg/L[5].毒死蜱對于多數水生生物屬于高毒性物質,并且可以通過生物富集和生物放大作用對水體食物鏈產生影響,最終威脅人類的健康[6-7].
斑馬魚(Danio rerio)具有體型小、易于管理、產卵量大、早期胚胎便于觀察等特點,已成為毒理學研究領域的模式生物[8-10].目前斑馬魚被廣泛應用于發育毒理學、內分泌干擾和生殖毒理學、神經毒理學、納米毒理學等毒理學研究中,成為揭示化學品對人類健康和環境的影響及其機制的重要模式生物,并促進了新技術和新分析方法的發展[11].
環境污染物能夠破壞機體氧化與抗氧化之間的平衡,從而導致機體內活性氧(ROS)等自由基產生過多,細胞自身無法及時清除,并產生大量氧化中間產物,進而導致機體氧化損傷[12-15].機體內抗氧化系統包括酶促系統(CAT、SOD、GSH 等) 和非酶促系統(維生素類、生物堿類等)[16].已有研究報道,毒死蜱會引起鯉魚肝臟損傷及抗氧化系統的異常[17].高劑量毒死蜱能誘導小鼠肺組織細胞的抗氧化能力下降,造成肺組織的病理損傷[18].毒死蜱對蚯蚓SOD和CAT活性具有顯著抑制作用,同時會造成脂質過氧化[19].本研究通過對氧化應激相關基因的表達量及酶活性的檢測,探究毒死蜱對斑馬魚胚胎的毒性作用機制.
1 材料與方法
1.1材料
AB品系斑馬魚購自國家斑馬魚資源中心(CZRC,China Zebrafish Resource Center).
毒死蜱(Chlorpyrifos,CPF)購自于成都化夏化學試劑有限公司,其純度≥97%;谷胱甘肽—S轉移酶(GST)測定試劑盒(比色法)、過氧化氫酶(CAT)測定試劑盒(可見光法)、丙二醛(MDA)測定試劑盒(TBA法)、總超氧化物歧化酶(T-SOD)測試盒(羥胺法)、總蛋白(TP)測定試劑盒(帶標準:考馬斯亮藍法)購自南京建成生物研究所.反轉錄試劑盒 PrimeScriptTM RT reagent Kit with gDNA Eraser (Perfect Real Time)(Takara);SYBR Premix Ex Taq II試劑盒(Takara).
1.2斑馬魚的養殖
AB品系野生型斑馬魚在獨立養殖系統中進行養殖,自來水經過活性炭過濾,紫外燈滅菌,在系統中自動循環,水溫控制在(28±0.5)℃,光周期14(光):10(暗).每天喂食2~3,食物為豐年蟲幼蟲.
胚胎收集方法.提前一個晚上將成年種魚按照雌雄2:1的比例裝入交配盒中,并用隔板將雌雄魚分開,第2d早上光照開始時(10h后)將隔板抽出,半個小時后收集胚胎.挑選發育正常的0.5~ 1.0hpf(孵化后小時)的胚胎,隨機分裝于6孔塑料板中(10個胚胎/孔),每孔培養液體積為10mL.
1.3暴露方法
選用甲醇作為助溶劑.在處理液中,甲醇的濃度為0.025%.前期的研究表明,毒死蜱處理液中低濃度(<0.1%)的甲醇不會影響實驗結果(數據未給出).設置6個毒死蜱暴露濃度,分別為0.25、0.50、0.75、1.00、1.25和1.50mg/L,每個濃度組中有20個胚胎,并設置3個生物學重復.6孔板的每個孔中加入10個發育正常的3hpf胚胎,加入10mL處理液.將胚胎置于恒溫培養箱中,溫度(28±0.5)℃,光周期14(光):10(暗).采用半靜態的試驗方法,暴露總時間為96h,每24h更換一次溶液,并及時挑出死卵.
1.4斑馬魚胚胎形態學觀察
進行CPF暴露處理后,每24h對斑馬魚胚胎進行一次鏡檢觀察.統計分析胚胎畸形率和死亡率,并計算96h半致死濃度(96h-LC50).
1.5氧化應激相關基因的表達分析
根據預試驗設立3個CPF濃度組,分別是0.24、0.47、0.94mg/L (1/5LC50、2/5LC50、4/5LC50).毒死蜱暴露96h后,每個濃度的胚胎分別取20枚保存,各3份.收集胚胎,液氮速凍,-80℃保存.
使用TRIzol試劑提取總RNA,260nm下估算總RNA濃度,質量則通過260/280比率評估.使用1.2%瓊脂糖凝膠電泳對RNA的質量進行進一步驗證.
樣品總RNA經反轉錄得到cDNA.各目標基因引物由上海生工公司合成(表1),經驗證所有引物擴增效率均良好(90%~110%).qRT-PCR程序為:95℃ 30s、95℃ 5s、60℃ 45s、55℃ 10s.基因表達的差異倍數通過2-ΔΔCt方法計算.每個濃度組3個生物學重復.

表1 氧化應激相關基因和β-actin的引物Table 1 Primer sequences of antioxidant response-related genes and β-actin
1.6氧化應激相關生化指標檢測
設立3個CPF濃度組,分別是0.24、0.47、0.94mg/L (1/5LC50、2/5LC50、4/5LC50),每個濃度組3個平行,每個平行60個胚胎.CPF暴露96h 后,收集胚胎,液氮速凍,-80℃保存.
5%組織勻漿液的制備.按照重量(g):體積(mL)=1:19的比例加入19倍生理鹽水,首先使用超聲破碎儀冰水浴破碎斑馬魚胚胎,離心取上清,得到5%組織勻漿液.
使用南京建成生物研究所酶活檢測試劑盒,檢測各項生化指標,包括總超氧化物歧化酶(T-SOD)活力、過氧化氫酶(CAT)活力、谷胱甘肽-S轉移酶(GST)活力、丙二醛(MDA)含量和總蛋白(TP)含量.
1.7統計分析
使用SPSS20.0通過Levene檢驗方法檢測數據的方差齊性,并進行one-way ANOVA數據分析.P<0.05作為統計顯著性的標準,所有值表示方法均為平均值±標準誤(SE).
2 結果與討論
2.1結果

圖1 毒死蜱暴露96h對胚胎的致畸作用Fig.1 Teratogenetic effect of CPF on embryos after a 96exposure

圖2 毒死蜱暴露96h胚胎死亡率Fig.2 Effects of CPF on embryonic mortality after a 96exposure
2.1.1毒死蜱對斑馬魚胚胎發育影響本研究中,采用梯度濃度的毒死蜱(0.25、0.50、0.75、1.00、1.25和1.50mg/L)對斑馬魚胚胎進行處理. 在96h觀察到的主要的幼魚畸形包括心包水腫、尾部彎曲和卵黃囊畸形(圖1b、c),并統計了這三種常見的畸形在毒死蜱0.25、0.50、0.75、1.00 和1.25mg/L處理組中的比例(圖1d),1.5mg/L處理組胚胎死亡率過高,沒有進行畸形率的統計.通過非線性曲線擬合得到毒死蜱96h半致死濃度為1.18mg/L (圖2).
2.1.2毒死蜱對斑馬魚胚胎氧化應激相關基因的影響農藥污染物可以改變機體抗氧化防御系統中相關基因的表達量,因此本研究選取斑馬魚的9個抗氧化相關基因作為參考,從轉錄水平揭示毒死蜱對斑馬魚胚胎氧化應激效應的發生機制.熒光定量PCR結果顯示(圖3),相對于空白對照組,0.24mg/L和0.47mg/L CPF處理組中基因Cu/Zn-sod表達顯著下調(0.54、0.22倍),并且呈現劑量-效應關系(圖3a).Mn-sod的表達量在高濃度CPF處理組中同樣顯著下調,并呈現劑量-效應關系(圖3b).cat的表達在0.47mg/L和0.94mg/L CPF處理組中顯著下調(0.51、0.12倍),并且呈現劑量-效應關系(圖3c).gstp2表達在0.24mg/L和0.47mg/L CPF處理組中顯著上調(1.58、1.88倍),但在最高濃度組與對照組相比并無顯著差異,呈現倒“U”形的劑量-效應關系(圖3d).在3個CPF處理組中,nrf2基因表達顯著上調(6.18、7.72、11.55倍),并且呈現劑量-效應關系(圖3e).keap1基因的表達在CPF處理組與對照組中無顯著差異(圖3f).ucp2的表達在0.47mg/L和0.94mg/L CPF處理組中顯著下調(0.68、0.34倍),并且呈現劑量-效應關系(圖3g).在0.47mg/L和0.94mg/L CPF處理組中,cox1顯著下調(0.65、0.38倍),并且呈現劑量-效應關系(圖3h).bcl2的表達在0.47mg/L和0.94mg/L CPF處理組中顯著下調(0.7、0.31倍),并且呈現劑量-效應關系(圖3i).
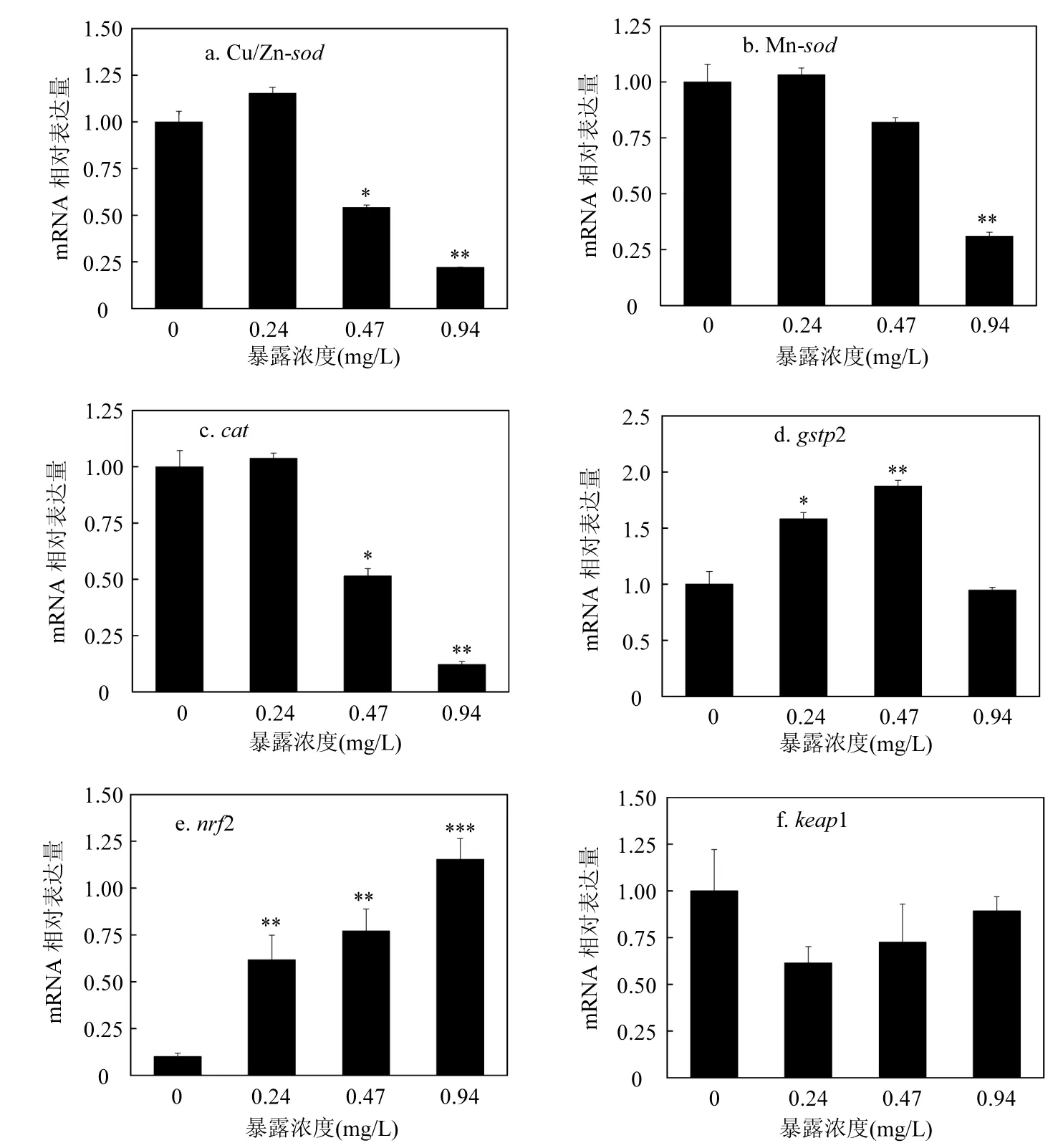

圖3 毒死蜱對氧化應激相關基因mRNA表達量的影響Fig.3 Effects of CPF on relative mRNA levels of antioxidant response-related genes與對照相比,*P<0.05,**P<0.01,***P<0.001
2.1.3毒死蜱對氧化應激酶活性的影響農藥污染物可以改變機體抗氧化防御系統中相關基因的表達量,進而改變機體內抗氧化酶的活性,同時也會造成脂質過氧化.由圖4可見,經0.24、0.47、0.94mg/L CPF處理后,胚胎總SOD活力分別是對照組的102%、81.3%、71.8%,0.94mg/L CPF處理組總SOD活力與對照組相比顯著降低,并呈現劑量-效應關系;處理組CAT活力分別是對照組的86.8%、76.7%、56.5%,CPF處理組CAT活力與對照組相比顯著降低,并呈現劑量-效應關系;處理組GST活力分別是對照組的136.7%、140.1%、119.5%;0.24、0.47、0.94mg/L CPF處理組MDA含量分別是對照組的305.4%、461.9%、558.1%,3個處理組與對照組相比顯著升高,并呈現劑量-效應關系.
2.2討論
本研究中,采用梯度濃度的毒死蜱對斑馬魚胚胎進行處理,毒死蜱暴露96h后可以觀察到,毒死蜱誘導胚胎產生了3種常見畸形心包水腫、尾部彎曲和卵黃囊畸形(圖1b、圖1c).經統計,3種畸形的畸形率的比例都隨著處理濃度的提高而顯著升高,呈現劑量-效應關系.由圖1d可看出,高濃度毒死蜱對斑馬魚胚胎有嚴重的致畸作用.同時毒死蜱也會對胚胎產生致死作用,致死率隨著處理濃度的提高而升高(圖2).
本研究通過檢測毒死蜱暴露96h斑馬魚胚胎抗氧化酶相關基因的表達及酶活力來研究毒死蜱對斑馬魚胚胎氧化應激的影響.SOD和CAT是抵御氧化損傷的第一道防線,其中Cu/Zn-sod 和Mn-sod是編碼超氧化物歧化酶(SOD)的基因,SOD的主要功能是清除氧自由基生成 H2O2和O2[20-22].過氧化氫酶(CAT)能夠清除體內產生的過多的H2O2,將其分解成O2和H2O,編碼基因是cat[20].在本研究中,高濃度毒死蜱暴露96h能夠下調Cu/Zn-sod、Mn-sod和cat的表達,并呈現劑量-效應關系(圖3a、圖3b、圖3c),這說明高濃度的毒死蜱會誘導胚胎產生氧化應激,導致胚胎體內產生過多的ROS無法及時清除,抑制了這3個基因的表達.酶活檢測的結果也顯示,高濃度毒死蜱能夠顯著抑制SOD和CAT活力,并呈現劑量-效應關系(圖4a、圖4b).
谷胱甘肽S-轉移酶(GST)具有Ⅱ相解毒酶和抗氧化酶的雙重功能,能夠保護機體免受環境污染物的損害,已被廣泛用作檢測環境污染物的生物指示物[23-26].在本研究中,毒死蜱能夠提高GST活力(圖4c),GST的編碼基因gstp2的表達檢測結果顯示,低濃度毒死蜱能夠顯著上調的表達,表明毒死蜱誘導胚胎產生解毒反應,但最高濃度毒死蜱并沒有顯著誘導gstp2的表達(圖3d),這可能是由于毒死蜱濃度過高,超過了胚胎的解毒能力,誘導機體產生過多的ROS,抑制了基因的表達.但在本研究中GST的活力在處理組與對照組中差異并不顯著,說明其對CPF脅迫并不敏感,但基因gstp2卻可作為CPF污染的潛在生物指示物.

圖4 毒死蜱對抗氧化酶活性和MDA含量的影響Fig.4 Effects of CPF on antioxidant enzyme activities and MDA content與對照相比,*P<0.05,**P<0.01,***P<0.001
轉錄因子NF-E2相關因子(nrf2),是細胞氧化應激反應中的關鍵因子,受胞質接頭蛋白(keap1)的調控,通過與抗氧化反應元件ARE相互作用,能夠上調抗氧化蛋白和II相解毒酶的表達,keap1-nrf2/ARE通路是細胞抗氧化反應的中樞調節者[27-28].在毒死蜱脅迫下,斑馬魚胚胎體內nrf2表達量顯著提高,同時keap1下調,能夠誘導nrf2的積聚及活性增強(圖3e、圖3f),從而上調抗氧化蛋白和II相解毒酶(GST等)的表達,誘導斑馬魚胚胎產生抗氧化反應.
過量ROS可侵害生物膜中的不飽和脂肪酸,引起脂質過氧化反應.丙二醛是膜脂過氧化最重要的產物之一,它的高低間接反映了機體細胞受自由基攻擊的程度[29].毒死蜱處理組胚胎MDA含量顯著高于對照組,并呈現劑量-效應關系(圖4d),說明毒死蜱誘導機體產生過多的ROS,引起脂質過氧化,侵害機體細胞,可能引起細胞凋亡.
cox1是細胞色素c氧化酶亞基I基因,細胞色素氧化酶是線粒體內呼吸鏈電子傳遞的終末復合物,是線粒體氧化能力的關鍵調節物質[30].毒死蜱能夠顯著下調其表達(圖3h),對細胞色素c 氧化酶產生抑制作用,說明毒死蜱可以抑制呼吸鏈,能夠減少ROS的產生和ATP的生成,同時電子傳遞和氧化磷酸化障礙能夠引起細胞凋亡.ucp2是解偶聯蛋白2的編碼基因,解偶聯蛋白2可以使氧化磷酸化過程解偶聯,從而使ATP合成減少,這可導致細胞功能下降,降低機體對損傷的耐受性;同時也能抑制呼吸鏈ROS的產生.而在機體正常生理條件下,90% ROS來自線粒體呼吸鏈,因此可保護組織和細胞,避免產生過多ROS對機體造成損傷.所以它對機體的作用既有利也有弊[31].毒死蜱能夠誘導ucp2顯著下調(圖3g),從而促進氧化磷酸化過程,有利于機體抵抗毒死蜱的氧化損傷作用.
bcl2編碼蛋白主要功能是抑制細胞凋亡[32],在毒死蜱脅迫下,其表達量顯著下調(圖3i),由此可見,毒死蜱破壞了機體內細胞凋亡的平衡.正常的細胞凋亡可以清除體內受損或者已經完成特定使命的細胞,是對機體的保護.而在毒死蜱脅迫下,斑馬魚胚胎體內MDA顯著上升,引起脂質過氧化,并且cox1下調,造成電子傳遞和氧化磷酸化障礙,在這些凋亡信號的刺激下,抑制凋亡基因bcl2的表達下調,凋亡平衡機制遭到破壞,可能會引起斑馬魚胚胎細胞凋亡過度,從而對胚胎造成損傷.
根據本研究結果,抗氧化酶SOD和CAT及抗氧化相關基因Cu/Zn-sod、Mn-sod、cat等都可成為CPF脅迫下斑馬魚胚胎氧化應激檢測的潛在生物指示物.但是在低濃度CPF脅迫下, SOD、CAT和GST的活性差異并不顯著.由此可見,在CPF脅迫下抗氧化酶活性及其相關基因的表達的改變并不完全一致,顯然抗氧化相關基因的表達對CPF脅迫更加敏感.MDA作為氧化損傷的指示物,在本研究中其在斑馬魚胚胎中的含量受CPF濃度的影響,并具有濃度-效應關系.
3 結論
通過對毒死蜱脅迫下斑馬魚胚胎形態學的觀察,以及氧化應激相關基因的表達和抗氧化酶含量的檢測,發現較高濃度的毒死蜱對斑馬魚胚胎有嚴重致畸和致死作用,并且毒死蜱能夠誘導斑馬魚胚胎產生氧化應激,誘導nrf2高表達,從而促進gstp2的表達.同時高濃度毒死蜱能夠抑制部分抗氧化酶(SOD、CAT)的活性及其相關基因(Cu/Zn-sod、Mn-sod、cat)的表達,引起MDA含量升高,造成脂質過氧化.CPF也能夠誘導呼吸鏈相關基因cox1、氧化磷酸化相關基因ucp2和抗凋亡基因bcl2表達顯著下調.結果表明毒死蜱會誘導斑馬魚胚胎產生氧化應激,造成脂質過氧化,導致斑馬魚胚胎氧化損傷.
參考文獻:
[1] 劉占山,黃安輝,肖明山.毒死蜱的研究應用現狀及產業發展前景 [J]. 世界農藥, 2009,31(S1):59-61.
[2] 汪家銘.毒死蜱市場競爭優勢及發展建議 [J]. 化工科技市場, 2010,33(6):11-15.
[3] 徐迎春,王松松,徐建軍,等.煙臺市農村飲用水水源地農藥殘留監測分析 [J]. 現代預防醫學, 2015,42(9):1704-1707.
[4] 吳長興,趙學平,吳聲敢,等.丘陵地區水稻田使用毒死蜱對水體的污染及其生態風險 [J]. 生態與農村環境學報, 2011,27(3): 108-112.
[5] 郭強,田慧,毛瀟萱,等.珠江河口水域有機磷農藥水生生態系統風險評價 [J]. 環境科學, 2014,35(3):1029-1034.
[6] Varó I, Serrano R, Pitarch E, et al. Navarro. Bioaccumulation of chlorpyrifos through an experimental food chain: study of protein HSP70as biomarker of sublethal stress in fish [J]. Archives of Environmental Contamination and Toxicology, 2002,42(2):229-235.
[7] Michiel A, Paul J Van den Brink, António J A. Impact of single and repeated applications of the insecticide chlorpyrifos on tropical freshwater plankton communities. Ecotoxicology, 2008, 17(8):756-771.
[8] 王佳佳,徐超,屠云杰,等.斑馬魚及其胚胎在毒理學中的實驗研究與應用進展 [J]. 生態毒理學報, 2007,2(2):123-135.
[9] 端正花,陳曉歐,劉靈麗,等.苯并三唑和鎘對斑馬魚肝臟的聯合毒性效應 [J]. 中國環境科學, 2015,35(1):1872-1876.
[10] 林琎,王紅艷,王開運,等.氟吡菌胺對斑馬魚的毒性效應 [J].中國環境科學, 2014,34(12):3230-3236.
[11] Bugel S M, Tanguay R L, Planchart A. Zebrafish: A marvel of high-throughput biology for 21st century toxicology [J]. Current Environmental Health Reports, 2014,1(4):341-352.
[12] Dong M, Zhu L S, Zhu S Y, et al. Toxic effects of 1-decyl-3-methylimidazolium bromide ionic liquid on the antioxidant enzyme system and DNA in zebrafish (Danio rerio) livers [J]. Chemosphere, 2013,91(8):1107-1112.
[13] 田文靜,白偉,趙春祿,等.納米ZnO對斑馬魚胚胎抗氧化酶系統的影響 [J]. 中國環境科學, 2010,30(5):705-709.
[14] Valavanidis A, Vlahogianni T, Dassenakis M, et al. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants [J]. Ecotoxicology andEnvironmental Safety, 2006,64(2):178-189.
[15] 黃權,蘇琳.動物體內氧化應激與抗氧化劑應用研究進展[J]. 中國獸藥雜志, 2013,47(5):66-69.
[16] 胡文琴,王恬,孟慶利.動物中活性氧的產生及清除機制 [J].家畜生態, 2004,25(3):64-67.
[17] 韓英,郝其睿,魏菁等.毒死蜱和阿特拉津對鯉肝臟的影響[J]. 東北農業大學學報, 2015,40(3):67-73.
[18] 焦銘,廖文莉,孫雪等.毒死蜱致小鼠肺細胞氧化損傷及維生素E的拮抗 [J]. 環境與健康雜志, 2014,31(2):134-136+189.
[19] 周世萍,段昌群,劉守慶,等.土壤中亞致死劑量毒死蜱對蚯蚓抗氧化防御系統酶活性的影響 [J]. 土壤學報, 2014,51(2):389-393.
[20] 張寶,周玫,陳瑗,等.細胞外超氧化物歧化酶(EcSOD) [J].醫學綜述, 2000,6(8):340-341.
[21] Jin Y X, Zheng S S, Pu Y, et al. Cypermethrin has the potential to induce hepatic oxidative stress, DNA damage and apoptosis in adult zebrafish (Danio rerio) [J]. Chemosphere, 2011,82(3):398-404.
[22] 趙飛,王擺,張曉娜,等.久效磷農藥對金魚肝細胞DNA的損傷及其機制研究 [J]. 中國環境科學, 2015,35(5):1563-1569.
[23] Matos P, Fonta?′nhas-Fernandesa A, Peixotoa F, et al. Biochemical and histological hepatic changes of Nile tilapia Oreochromis niloticus exposed to carbaryl [J]. Pesticide Biochemistry and Physiology, 2007,89(1):73-80.
[24] Glisic B, Mihaljevic I, Popovic M, et al. Characterization of glutathione-S-transferases in zebrafish (Danio rerio) [J]. Aquatic Toxicology, 2015,158:50-62.
[25] 雷安平,陳歡,黎雙飛,等.谷胱甘肽S-轉移酶的功能、應用及克隆表達 [J]. 環境科學與技術, 2009,32(12):85-91.
[26] Garner L V, Di Giulio R T. Glutathione transferase pi class 2 (GSTp2) protects against the cardiac deformities caused by exposure to PAHs but not PCB-126in zebrafish embryos [J]. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2012,155(4):573-579.
[27] Wang L, Gallagher E P. Role of Nrf2 antioxidant defense in mitigating cadmium-induced oxidative stress in the olfactory system of zebrafish [J]. Toxicology and Applied Pharmacology, 2013,266(2):177-186.
[28] 李煌元.Keap1-Nrf2/ARE通路在分子毒理學中的研究進展 [J].國外醫學(衛生學分冊), 2006,33(3):129-135.
[29] 李紅,王克躍,石修權.鋅對鎘所致大鼠肝臟脂質過氧化損傷的保護作用研究 [J]. 遵義醫學院學報, 2004,27(4):322-323.
[30] 史華,張洪波,陳波.丹參對腎缺血再灌注損傷細胞色素C氧化酶活性的影響 [J]. 黑龍江醫藥科學, 2003,26(2):11-12.
[31] 賈曉麗.UCP2的研究進展 [J]. 赤峰學院學報(自然科學版), 2011,27(7):39-41.
[32] Yu K M, Li G C, Feng W M, et al. Chlorpyrifos is estrogenic and alters embryonic hatching, cell proliferation and apoptosis in zebrafish [J]. Chemico-Biological Interactions, 2015,239:26-33.
Oxidative stress effects of chlorpyrifos on zebrafish embryos.
ZHANG Jia-yu, LIU Li-li, LI Guo-chao, YU Kai-min, LV Peng, YAN Yan-chun*(Graduate School, Chinese Academy of Agricultural Sciences, Beijing 100081, China). China Environmental Science, 2016,36(3):927~934
Abstract:We elucidated the effects of chlorpyrifos exposure on the morphology, oxidative stress and oxidative damage in zebrafish (Danio rerio) embryos. After exposed to concentration gradient of chlorpyrifos solution for 96hours, the rates of deformities and death were dramatically higher than those in the control groups, and the median lethal concentration of CPF was calculated to be 1.18mg/L after a 96h exposure. The expression levels of oxidative stress-related genes, antioxidant enzyme activities and malondialdehyde (MDA) content were investigated. In CPF treatment groups, the antioxidant enzyme (SOD, CAT) activities and the expression levels of their encoding genes (Cu/Zn-sod, Mn-sod, cat) significantly decreased, compared with the control groups. At low concentrations of chlorpyrifos, antioxidant enzyme activities were not affected significantly, while the expression levels of their encoding genes were more sensitive to chlorpyrifos. Chlorpyrifos could induce the expression of nrf2, which played an important role in increased expression of a group of genes encoding antioxidant and phaseⅡdetoxification enzymes, but the kinetics of GST and gstp2 were not consistent with each other. The down-regulation of expression levels of ucp2 and cox1 made contributions to reducing ROS generated by the respiratory chain in CPF treatment groups. Meanwhile, in CPF treatment groups, the expression level of bcl2 was significantly down-regulated, suggesting that the balance of apoptosis was destroyed. MDA content was significantly higher in CPF treatment groups than that in control, indicating that chlorpyrifos could cause oxidative damage in zebrafish embryos.
Key words:chlorpyrifos;zebrafish embryo;oxidative stress;oxidative damage
作者簡介:張家禹(1990-),男,遼寧大連人,碩士研究生,主要從事微生物分子生物學與基因工程研究.
基金項目:國家自然科學基金項目(31170119),中國農科科學院基礎研究基金項目(0042014006,0042012003,0042011006)
收稿日期:2015-06-25
中圖分類號:X171.5
文獻標識碼:A
文章編號:1000-6923(2016)03-0927-08
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
